
Multi-Environment Testing Based G × E Interactions Reveal Stable Host-Plant Resistance against Sterility Mosaic Disease in Pigeonpea

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preliminary SMD Field Screening
2.2. Genotype Assessment at Multi-Environment Locations
2.3. Data Observation and Statistical Analysis
3. Results
3.1. Disease Symptoms and Impact
3.2. Preliminary SMD Field Screening
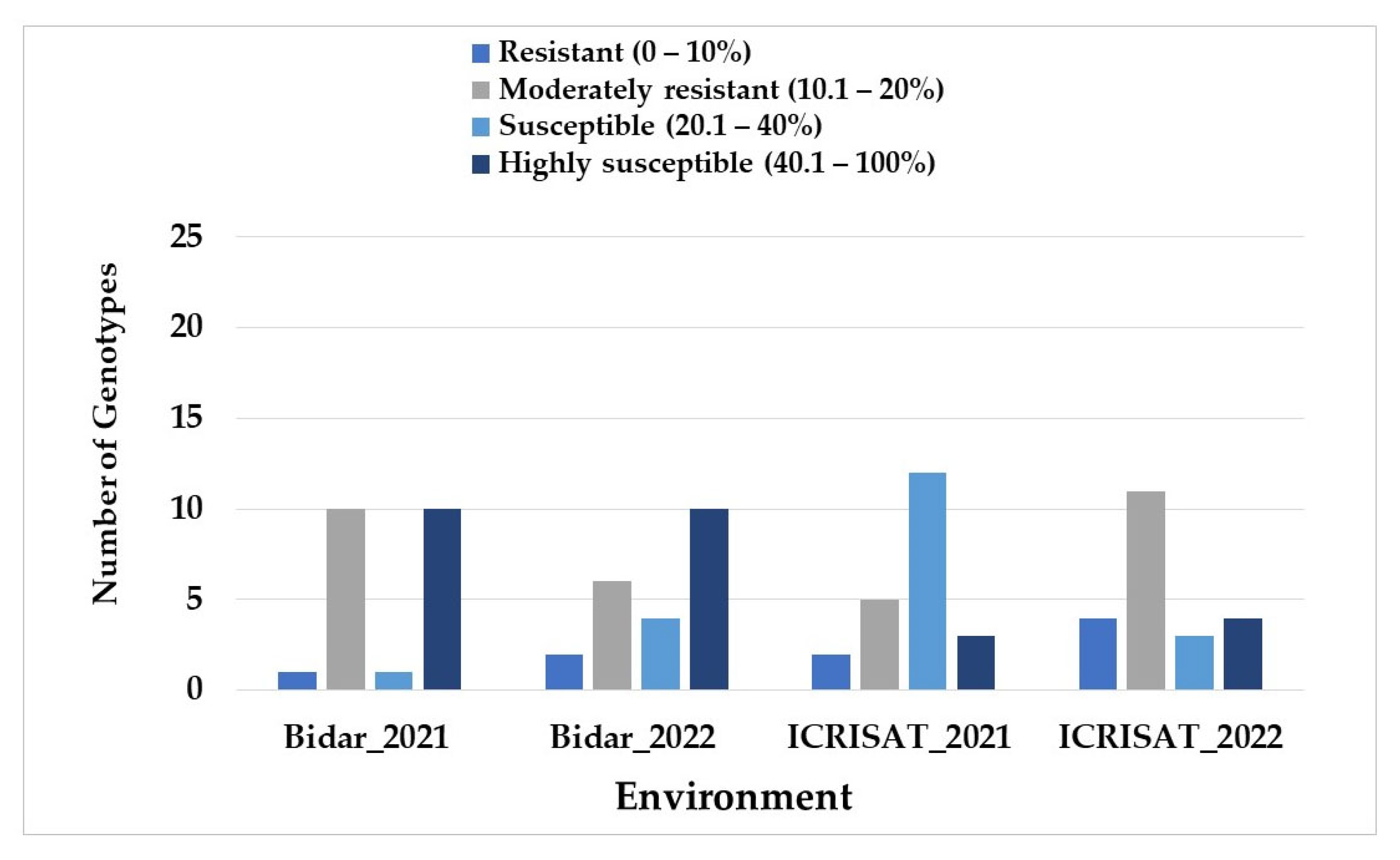
3.3. Multi-Environmental Testing of SMD
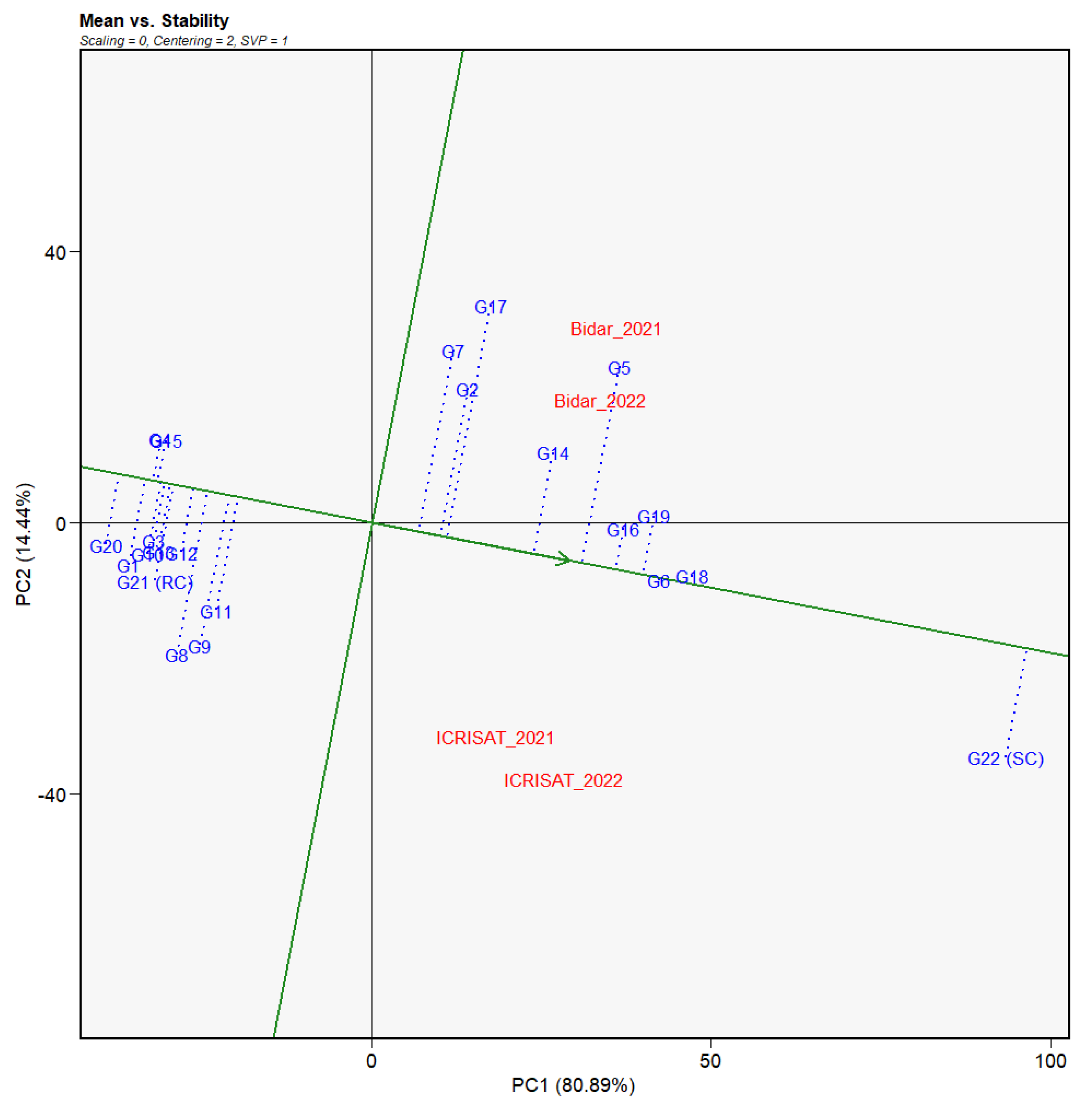
3.4. Stability Analysis of Genotypes and Environments
3.4.1. Evaluation of Factor G
3.4.2. Evaluation of Factor E
3.5. Identification of Mega-Environments
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van der Maesen, L.J.G. Pigeonpea: Origin, history, evolution and taxonomy. In The Pigeonpea; Nene, Y.L., Hall, S.D., Sheila, V.K., Eds.; CAB International: Wallingford, UK, 1990; pp. 15–46. [Google Scholar]
- FAOSTAT. Food and Agriculture Organization of the United Nations, Rome. Available online: http://faostat.fao.org.2021 (accessed on 30 March 2022).
- Ae, N.; Arihara, J.; Okada, K.; Yoshihara, T.; Johansen, C. Phosphorus uptake by pigeonpea and its role in cropping system of the Indian subcontinent. Curr. Sci. 1990, 248, 477–480. [Google Scholar]
- Saxena, K.B.; Kumar, R.V.; Rao, P.V. Pigeonpea nutrition and its improvement. J. Crop Prod. 2002, 5, 227–260. [Google Scholar] [CrossRef]
- Singh, N.; Tyagi, R.K.; Pandey, C. Genetic Resources of Pigeonpea (Cajanus cajan): Conservation for Use; National Bureau of Plant Genetic Resources: New Delhi, India, 2013. [Google Scholar]
- Sharma, M.; Telangre, R.; Ghosh, R.; Pande, S. Multi-environment field testing to identify broad, stable resistance to sterility mosaic disease of pigeonpea. J. Gen. Plant Pathol. 2015, 81, 249–259. [Google Scholar] [CrossRef]
- Ghosh, R.; Tarafdar, A.; Mediga, K.R.; Srinivas, K.; Sharma, M. Pigeonpea crop improvement: Genomics and post genomics. In Smart Plant Breeding for Field Crops in Post-Genomics Era; Springer Nature: Singapore, 2023; pp. 271–292. [Google Scholar]
- Reddy, M.V.; Sharma, S.B.; Nene, Y.L. Pigeonpea disease management. In The Pigeonpea; Nene, Y.L., Hall, S.D., Sheila, V.K., Eds.; CAB International: Wallingford, UK, 1990; pp. 303–347. [Google Scholar]
- Mitra, M. Report of the Imperial Mycologist; Scientific Reports of the Indian Agriculture Research Institute: Calcutta, India, 1931; pp. 58–71. [Google Scholar]
- Kumar, P.L.; Jones, A.T.; Sreenivasulu, P.; Reddy, D.V.R. Break-through in the identification of the causal virus of pigeonpea sterility mosaic disease. J. Mycol. Plant Pathol. 2000, 30, 249. [Google Scholar]
- Elbeaino, T.; Digiaro, M.; Uppala, M.; Sudini, H.K. Deep sequencing of dsRNAs recovered from mosaic-diseased pigeonpea reveals the presence of a novel emaravirus, pigeonpea sterility mosaic virus. Arch. Virol. 2015, 160, 2019–2029. [Google Scholar] [CrossRef] [PubMed]
- Seth, M.L. Transmission of pigeonpea sterility by an eriophyid mite. Indian Phytopathol. 1962, 15, 225–227. [Google Scholar]
- Kulkarni, N.K.; Kumar, P.L.; Muniyappa, V.; Jones, A.T.; Reddy, D.V.R. Transmission of Pigeonpea sterility mosaic virus by the eriophyid mite, Aceria cajani (Acari: Arthropoda). Plant Dis. 2002, 86, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Patil, B.L.; Meenakshi, D.; Ritesh, M. Variability of Emaravirus species associated with sterility mosaic disease of pigeonpea in India provides evidence of segment reassortment. Viruses 2017, 9, 183. [Google Scholar] [CrossRef]
- Pande, S.; Sharma, M.; Gopika, G.; Rameshwar, T. High Throughput Phenotyping of Pigeonpea Diseases: Stepwise Identification of Host Plant Resistance; Information Bulletin No. 93; International Crops Research Institute for the Semi-Arid Tropics: Patancheru, India, 2012. [Google Scholar]
- Sharma, M.; Rathore, A.; Mangala, U.N.; Ghosh, R.; Sharma, S.; Upadhyay, H.D.; Pande, S. New sources of resistance to Fusarium wilt and sterility mosaic disease in a mini-core collection of pigeonpea germplasm. Eur. J. Plant Pathol. 2012, 133, 707–714. [Google Scholar] [CrossRef]
- Kumar, P.L.; Jones, A.T.; Waliyar, F. Virus diseases of pigeonpea. In Characterization, Diagnosis and Management of Plant Viruses; Vol. 3: Vegetable and Pulse, Crops; Rao, G.P., Kumar, P.L., Holguin-Pena, R.J., Eds.; Studium Press: Houston, TX, USA, 2008; pp. 235–258. [Google Scholar]
- Sayiprathap, B.R.; Patibanda, A.K.; Prasanna Kumari, V.; Jayalalitha, K.; Ramappa, H.K.; Rajeswari, E.; Karthiba, L.; Saratbabu, K.; Sharma, M.; Sudini, H.K. Salient Findings on Host Range, Resistance Screening, and Molecular Studies on Sterility Mosaic Disease of Pigeonpea Induced by Pigeonpea sterility mosaic viruses (PPSMV-I and PPSMV-II). Front. Microbiol. 2022, 13, 838047. [Google Scholar] [CrossRef]
- Amin, K.S.; Reddy, M.V.; Nene, Y.L.; Raju, T.N.; Shukla, P.; Zote, K.K.; Arjunan, G.; Bendre, N.J.; Rathi, Y.P.; Sinha, B.K.; et al. Multi location evaluation of pigeonpea (Cajanus cajan) for broad-based resistance to sterility-mosaic disease in India. Indian J. Agric. Sci. 1993, 63, 542–546. [Google Scholar]
- Nene, Y.L.; Reddy, M.V.; Beniwal, S.P.S.; Mahmood, M.; Zote, K.K.; Singh, R.N.; Sivaprakasam, K. Multilocational testing of pigeonpea for broad-based resistance sterility mosaic in India. Indian Phytopathol. 1989, 42, 444–448. [Google Scholar]
- Sharma, M.; Pande, S. New sources of resistance to Fusarium wilt, sterility mosaic disease and Phytophthora blight in vegetable pigeonpea germplasm. Indian J. Plant Prot. 2011, 39, 288–293. [Google Scholar]
- Reddy, M.V.; Raju, T.N.; Ghanekar, A.M.; Amin, K.S.; Arjunan, G.; Nene, Y.L.; Ghanekar, A. Variability in sterility mosaic pathogen of pigeonpea in India. Indian Phytopathol. 1993, 46, 206–212. [Google Scholar]
- Smiley, R.W.; Yan, H. Variability of fusarium crown rot tolerances among cultivars of spring and winter wheat. Plant Dis. 2009, 93, 954–961. [Google Scholar] [CrossRef] [PubMed]
- Parihar, A.K.; Basandrai, A.K.; Sirari, A.; Dinakaran, D.; Singh, D.; Kannan, K.; Kushawaha, K.P.; Adinarayan, M.; Akram, M.; Latha, T.K.; et al. Assessment of mungbean genotypes for durable resistance to yellow mosaic disease: Genotype × environment interactions. Plant Breed. 2017, 136, 94–100. [Google Scholar] [CrossRef]
- Nene, Y.L.; Reddy, M.V. A new technique to screen pigeonpea for resistance to sterility mosaic. Trop. Grain Legume Bull. 1976, 5, 23–24. [Google Scholar]
- Yan, W.; Cornelius, P.L.; Crossa, J.; Hunt, L.A. Two types of GGE biplots for analyzing multi-environment trial data. Crop Sci. 2001, 41, 656–663. [Google Scholar] [CrossRef]
- Yan, W.; Kang, M.S. GGE Biplot Analysis: A Graphical Tool for Breeders, Geneticists, and Agronomists; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Yan, W.; Falk, D.E. Biplot analysis of host-by-pathogen data. Plant Dis. 2002, 86, 1396–1401. [Google Scholar] [CrossRef]
- Sharma, M.; Sharath Chandran, U.S.; Rani, U.; Singh, S.K.; Basandrai, A.K.; Basandrai, D. Stability and suitability of genotypes and environment to Ascochyta blight of chickpea. Front. Plant Sci. 2023, 14, 1006099. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Nene, Y.L. Sterility mosaic of pigeonpea: The challenge continues. Indian J. Mycol. Plant Pathol. 1995, 25, 1–11. [Google Scholar]
- Jones, A.T.; Kumar, P.L.; Saxena, K.B.; Kulkarni, N.K.; Muniyappa, V.; Waliyar, F. Sterility mosaic disease-the Green Plague of pigeonpea, advances in understanding the etiology, transmission and control of a major virus disease. Plant Dis. 2004, 88, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.L.; Jones, A.T.; Reddy, D.V.R. A novel mite-transmitted virus with a divided RNA genome closely associated with pigeonpea sterility mosaic disease. Phytopathology 2003, 93, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, N.K.; Reddy, A.S.; Kumar, P.L.; Vijaynarasimha, J.; Rangaswamy, K.T.; Muniyappa, V.; Reddy, L.J.; Saxena, K.B.; Jones, A.T.; Reddy, D.V. Broad-based resistance to pigeonpea sterility mosaic disease in the accessions of Cajanus scarabaeoides. Indian J. Plant Prot. 2003, 31, 6–11. [Google Scholar]
- Nagaraj, K.M.; Chikkadevaiah Muniyappa, V.; Rangaswamy, K.T.; Kumar, P.L. Evaluation of pigeonpea genotypes for resistance to Pigeonpea sterility mosaic virus-B isolate. Indian J. Plant Prot. 2006, 34, 216–220. [Google Scholar]
- Alam, A.K.M.M.; Somta, P.; Jompuk, C.; Chatwachirawong, P.; Srinives, P. Evaluation of mungbean genotypes based on yield stability and reaction to mungbean yellow mosaic virus disease. Plant Pathol. J. 2014, 30, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Dipshikha, K.; Seweta, S.; Chandra, N.B.; Chauhan, V.B.; Singh, R.N. Correlation between mite population (Aceria cajani) and environmental factors causing sterility mosaic disease of pigeonpea. Int. J. Life Sci. 2013, 1, 228–232. [Google Scholar]
- Pallavi, M.S.; Ramappa, H.K.; Singh, P. Pigeonpea sterility mosaic virus disease in Karnataka: Epidemiological aspects. J. Pharmacogn. Phytochem. 2019, 8, 893–899. [Google Scholar]
- Pour-Aboughadareh, A.; Khalili, M.; Poczai, P.; Olivoto, T. Stability indices to deciphering the genotype-by-environment interaction (GEI) effect: An applicable review for use in plant breeding programs. Plants 2022, 11, 414. [Google Scholar] [CrossRef]
- Yan, W.; Kang, M.S.; Ma, B.; Woods, S.; Cornelius, P.L. GGE biplot vs. AMMI analysis of genotype-by-environment data. Crop Sci. 2007, 47, 643–653. [Google Scholar] [CrossRef]
- Das, A.; Parihar, A.K.; Saxena, D.; Singh, D.; Singha, K.D.; Kushwaha, K.P.S.; Chand, R.; Bal, R.S.; Chandra, S.; Gupta, S. Deciphering genotype-by-environment interaction for targeting test environments and rust resistant genotypes in field pea (Pisum sativum L.). Front. Plant Sci. 2019, 10, 825. [Google Scholar] [CrossRef] [PubMed]
- Ganapathy, K.N.; Byre Gowda, M.; Ajay, B.C.; Venkatesha, S.C.; Gnanesh, B.N.; Gomashe, S.S.; Prasanth, B.; Girish, G.; Prasad, P.S.; Veerakumar, G.N.; et al. Inheritance studies of sterility mosaic disease (SMD) resistance in vegetable type pigeonpea (Cajanus cajan (L.) Millsp.). Aust. J. Crop Sci. 2012, 6, 1154–1158. [Google Scholar]
- Yan, W.; Pageau, D.; Frégeau-Reid, J.; Durand, J. Assessing the representativeness and repeatability of test locations for genotype evaluation. Crop Sci. 2011, 51, 1603–1610. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Genotype | Code | Duration | Environment | Mean (Genotype) | Reaction Type | |||
|---|---|---|---|---|---|---|---|---|---|
| Bidar_2021 | Bidar_2022 | ICRISAT_2021 | ICRISAT_2022 | ||||||
| 1 | NAM 2082 | G1 | Mid Early | 13.00 | 15.52 | 19.06 | 10.92 | 14.63 | MR |
| 2 | NAM 2085 | G2 | Mid Early | 59.09 | 47.74 | 23.21 | 16.90 | 36.74 | S |
| 3 | NAM 2088 | G3 | Mid Early | 17.39 | 17.59 | 12.18 | 15.38 | 15.64 | MR |
| 4 | NAM 2089 | G4 | Mid Early | 19.23 | 30.07 | 6.76 | 3.67 | 14.93 | MR |
| 5 | NAM 2092 | G5 | Mid Early | 67.86 | 68.87 | 28.82 | 24.00 | 47.39 | HS |
| 6 | NAM 2150 | G6 | Mid Early | 62.50 | 53.57 | 35.08 | 57.14 | 52.07 | HS |
| 7 | NAM 2151 | G7 | Mid Early | 57.14 | 52.38 | 20.11 | 11.26 | 35.22 | MS |
| 8 | GRG 811 | G8 | Mid Early | 16.67 | 9.17 | 34.06 | 19.05 | 19.73 | MR |
| 9 | TS-3R-58-53-1 | G9 | Mid Early | 7.14 | 27.08 | 39.89 | 13.42 | 21.88 | S |
| 10 | TS-3R-58-53-2 | G10 | Mid Early | 15.00 | 17.71 | 15.89 | 13.55 | 15.54 | MR |
| 11 | GRG 622 | G11 | Medium | 22.22 | 15.88 | 33.30 | 16.82 | 22.05 | S |
| 12 | GRG 152 | G12 | Medium | 17.86 | 21.34 | 20.97 | 12.96 | 18.28 | MR |
| 13 | KRG 33 | G13 | Medium | 18.52 | 16.67 | 23.21 | 9.02 | 16.85 | MR |
| 14 | GL 1 | G14 | Medium | 60.00 | 54.58 | 32.46 | 28.08 | 43.78 | HS |
| 15 | NAM 2162 | G15 | Medium | 20.00 | 29.76 | 5.03 | 5.71 | 15.13 | MR |
| 16 | NAM 2545 | G16 | Medium | 66.67 | 49.72 | 39.88 | 42.31 | 49.64 | HS |
| 17 | NAM 2435 | G17 | Medium | 64.71 | 56.35 | 19.66 | 8.89 | 37.40 | S |
| 18 | GRG-617-2 | G18 | Medium | 65.00 | 58.60 | 46.78 | 50.62 | 55.25 | HS |
| 19 | GRG-617-3 | G19 | Medium | 67.86 | 56.42 | 46.85 | 37.78 | 52.23 | HS |
| 20 | BSMR 736 | G20 | Medium | 17.24 | 8.13 | 10.97 | 12.60 | 12.23 | MR |
| 21 | ICPL 87119 (RC | G21 | Medium | 17.86 | 13.24 | 26.76 | 10.82 | 17.17 | MR |
| 22 | ICP 8863 (SC) | G22 | Medium | 70.59 | 87.54 | 70.46 | 91.38 | 79.99 | HS |
| Mean (Environment) | 38.34 | 36.72 | 27.79 | 23.29 | - | - | |||
| Source of Variation | Degree of Freedom | Sum of Squares | Mean Sum of Squares | F Value | Pr (> F) | Variation (%) a |
|---|---|---|---|---|---|---|
| Environment | 3 | 2866.70 | 955.57 | 21.30 | 2.37E-10 | 7.32 |
| Genotype | 21 | 24,871.24 | 1184.34 | 26.40 | 1.93E-28 | 63.56 |
| G × E | 63 | 7624.28 | 121.02 | 2.70 | 1.22E-05 | 19.48 |
| Error | 84 | 3768.87 | 44.87 | - | - | - |
| Total | 171 | 39,131.09 | - | - | - | |
| CV (%) | 20.27937 | |||||
| Environments | Bidar_2021 | Bidar_2022 | ICRISAT_2021 | ICRISAT_2022 |
|---|---|---|---|---|
| Bidar_2021 | 1.0000000 | |||
| Bidar_2022 | 0.8548024 | 1.0000000 | ||
| ICRISAT_2021 | 0.4915255 | 0.4455110 | 1.0000000 | |
| ICRISAT_2022 | 0.5395481 | 0.4940711 | 0.7809147 | 1.0000000 |
| DI | Max. Tm | Min. Tm | RH (%) | Rainfall | Wind Velocity | |
|---|---|---|---|---|---|---|
| DI | 1 | |||||
| Max. Tm | −0.93305 | 1 | ||||
| Min. Tm | −0.52454 | 0.35285 | 1 | |||
| RH (%) | 0.03401 | −0.39119 | 0.34456 | 1 | ||
| Rainfall | 0.50298 | −0.21619 | −0.14835 | −0.69701 | 1 | |
| Wind Velocity | −0.22946 | 0.38731 | −0.70704 | −0.47014 | −0.20129 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mediga, K.R.; Sunkad, G.; Kulkarni, S.; Chandran, U.S.S.; Ghosh, R.; Kshirsagar, D.; Sonnappa, M.; Katravath, S.; Parthasarathy, A.; Sharma, M. Multi-Environment Testing Based G × E Interactions Reveal Stable Host-Plant Resistance against Sterility Mosaic Disease in Pigeonpea. Agronomy 2023, 13, 2859. https://doi.org/10.3390/agronomy13122859
Mediga KR, Sunkad G, Kulkarni S, Chandran USS, Ghosh R, Kshirsagar D, Sonnappa M, Katravath S, Parthasarathy A, Sharma M. Multi-Environment Testing Based G × E Interactions Reveal Stable Host-Plant Resistance against Sterility Mosaic Disease in Pigeonpea. Agronomy. 2023; 13(12):2859. https://doi.org/10.3390/agronomy13122859
Chicago/Turabian StyleMediga, Kasi Rao, Gururaj Sunkad, Sunil Kulkarni, U. S. Sharath Chandran, Raju Ghosh, Dipak Kshirsagar, Muniswamy Sonnappa, Srinivas Katravath, Ashwini Parthasarathy, and Mamta Sharma. 2023. "Multi-Environment Testing Based G × E Interactions Reveal Stable Host-Plant Resistance against Sterility Mosaic Disease in Pigeonpea" Agronomy 13, no. 12: 2859. https://doi.org/10.3390/agronomy13122859
APA StyleMediga, K. R., Sunkad, G., Kulkarni, S., Chandran, U. S. S., Ghosh, R., Kshirsagar, D., Sonnappa, M., Katravath, S., Parthasarathy, A., & Sharma, M. (2023). Multi-Environment Testing Based G × E Interactions Reveal Stable Host-Plant Resistance against Sterility Mosaic Disease in Pigeonpea. Agronomy, 13(12), 2859. https://doi.org/10.3390/agronomy13122859