
Composition of Strawberry Flower Volatiles and Their Effects on Behavior of Strawberry Pollinators, Bombus terrestris and Apis mellifera

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strawberry Variety and Bee Species
2.2. Flower Volatiles Collection and Detection
2.3. EAG Responses to Volatile Compounds
2.4. Y-Tube Olfactometer Behavior Tests to Volatile Compounds
2.5. Statistical Analyses
3. Results
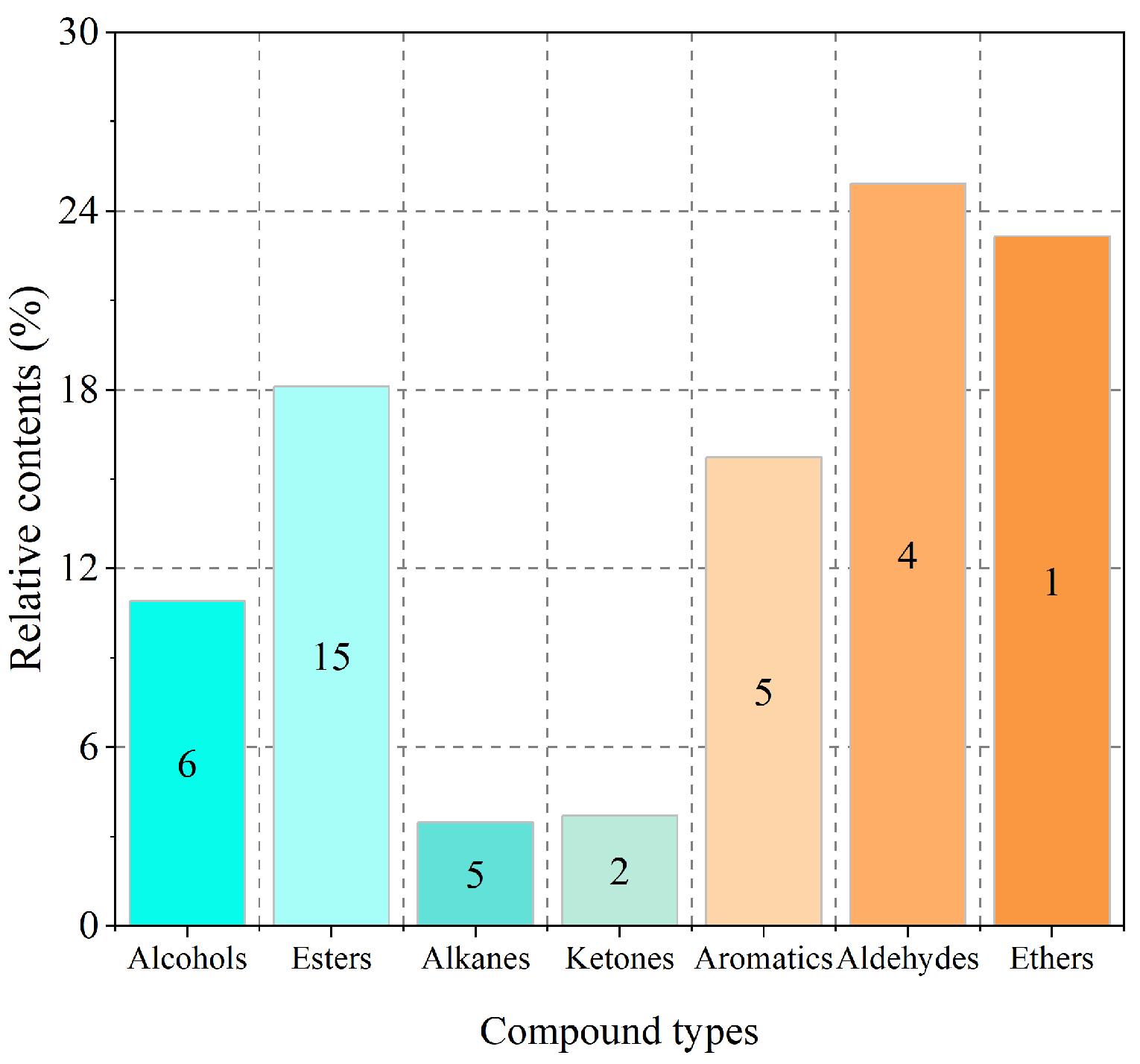
3.1. Types and Contents of Volatile Compounds of Strawberry Flowers
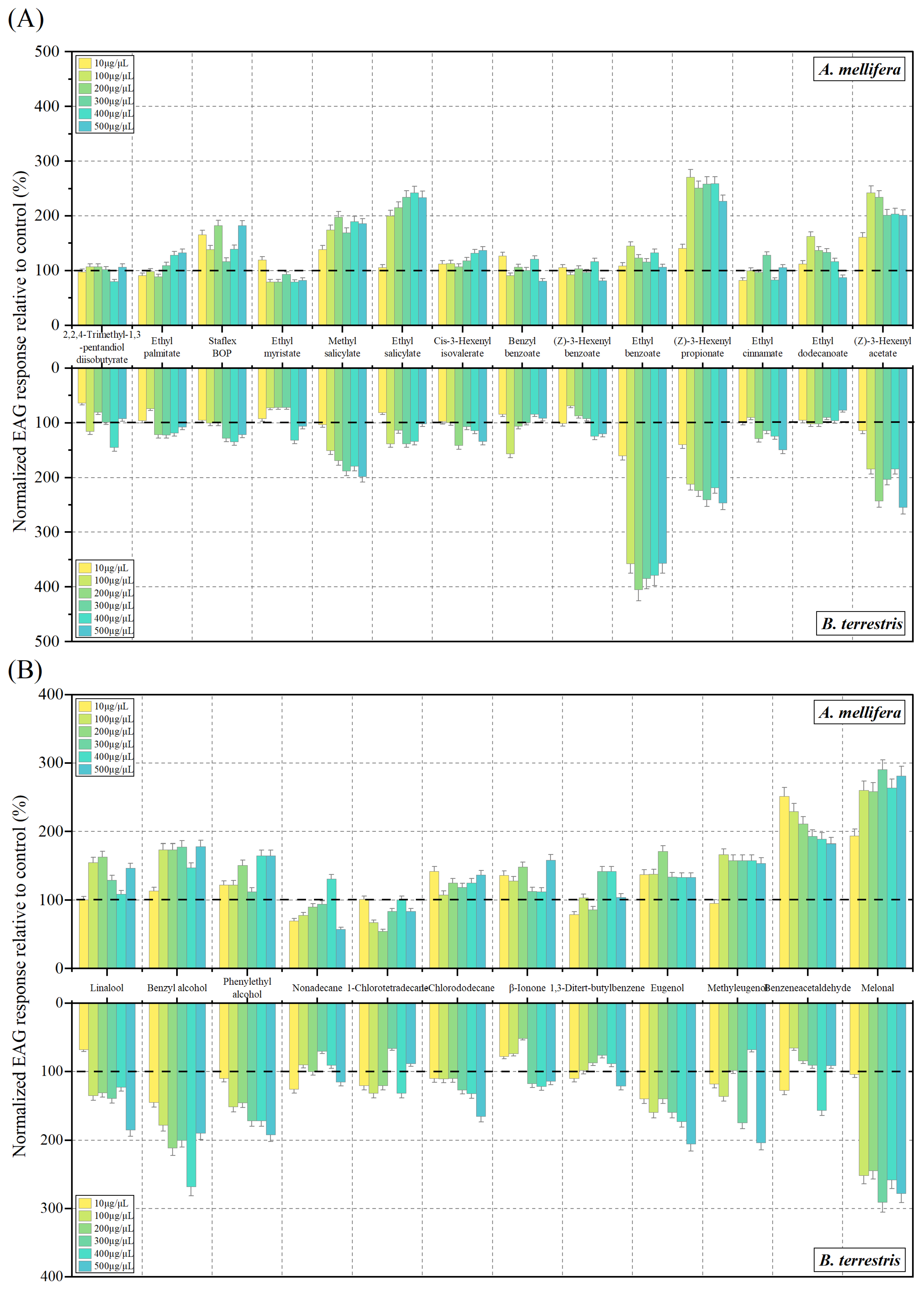
3.2. EAG Response to Volatile Compounds
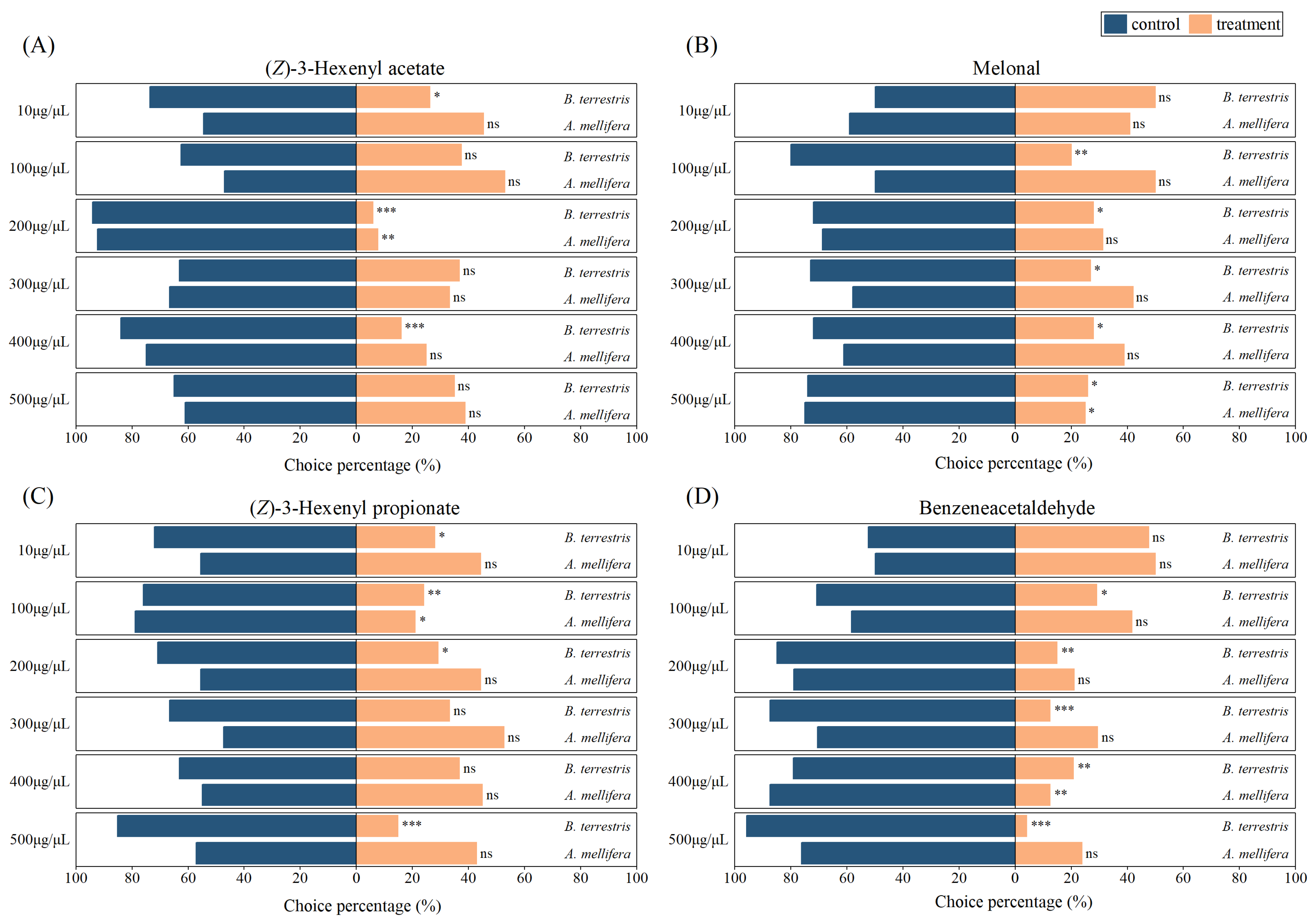
3.3. Y-Tube Behavior Tests of Bees
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahrenfeldt, E.J.; Klatt, B.K.; Arildsen, J.; Trandem, N.; Andersson, G.K.S.; Tscharntke, T.; Smith, H.G.; Sigsgaard, L. Pollinator communities in strawberry crops–variation at multiple spatial scales. Bull. Entomol. Res. 2015, 105, 497–506. [Google Scholar] [CrossRef]
- Fu, B.; Yang, J. Application of common pollination bees in facility agriculture. J. Shanxi Agric. Sci. 2014, 42, 925–928. [Google Scholar]
- Klein, A.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, C.D.; Fountain, M.T.; Brown, M.J.F. Varietal and seasonal differences in the effects of commercial bumblebees on fruit quality in strawberry crops. Agric. Ecosyst. Environ. 2019, 281, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Han, C.; Breeze, T.D.; Li, M.; Mashilingi, S.K.; Hua, J.; Zhang, W.; Zhang, X.; Zhang, S.; An, J. Bumblebee pollination enhances yield and flavor of tomato in gobi desert greenhouses. Agriculture 2022, 12, 795. [Google Scholar] [CrossRef]
- Adala, A.G. Pollination of Strawberry in Kenya, by Stingless Bees (Hymenoptera: Meliponini) and Honey Bee (Hymenoptera: Apini) for Improved Fruit Quality. Ph.D. Thesis, University of Nairobi, Nairobi, Kenya, 2012. [Google Scholar]
- Chen, W.; An, J.; Dong, J.; Ding, K.; Gao, S. Flower-visiting behavior and pollination ecology of different bee species on greenhouse strawberry. Chin. J. Ecol. 2011, 30, 290–296. [Google Scholar]
- Luo, W.; Cao, L.; Gao, L.; Cheng, S.; Yin, S.; Wang, R.; Dai, R.; Liang, M.; Xie, H.; Liu, J. Yield, quality and efficiency of different bee pollination on greenhouse strawberry in Chongqing. J. Yunnan Agric. Univ. Nat. Sci. 2017, 32, 498–503. [Google Scholar]
- Li, J.; Peng, W.; Wu, J.; An, J.; Guo, Z.; Tong, Y.; Huang, J. Strawberry pollination by Bombus lucorum and Apis mellifera in greenhouses. Acta Entomol. Sin. 2006, 49, 342–348. [Google Scholar]
- Li, S.; Tan, G.; Yang, J.; Li, X.; Zhang, Y.; Jiang, Y. Effects on yield and quality of strawberry with bumblebee pollination. J. Changjiang Vege. 2016, 2, 56–58. [Google Scholar]
- Sun, M.; Chen, R.; Chen, J.; Chai, W.; Wang, Y.; Yao, H.; Zhou, J. Effects of the bumblebee and honeybee pollination on yield and fruit quality of strawberry. J. Changjiang Vege. 2008, 11b, 42–44. [Google Scholar]
- Zaitoun, S.T.; Al-Ghzawi, A.A.; Shannag, H.K.; Al-Tawaha, A.R.M. Comparative study on the pollination of strawberry by bumble bees and honeybees under plastic house conditions in Jordan valley. J. Food Agric. Environ. 2006, 4, 237–240. [Google Scholar]
- Parachnowitsch, A.L.; Kessler, A. Pollinators exert natural selection on flower size and floral display in Penstemon digitalis. New Phytol. 2010, 188, 393–402. [Google Scholar] [CrossRef]
- Dötterl, S.; Vereecken, N.J. The chemical ecology and evolution of bee-flower interactions: A review and perspectives. Can. J. Zool. 2010, 88, 668–697. [Google Scholar] [CrossRef]
- Schiestl, F.P. Ecology and evolution of floral volatile-mediated information transfer in plants. New Phytol. 2015, 206, 571–577. [Google Scholar] [CrossRef]
- Wright, G.A.; Schiestl, F.P. The evolution of floral scent: The influence of olfactory learning by insect pollinators on the honest signalling of floral rewards. Funct. Ecol. 2009, 23, 841–851. [Google Scholar] [CrossRef] [Green Version]
- Mozūraitis, R.; Hall, D.; Trandem, N.; Ralle, B.; Tunström, K.; Sigsgaard, L.; Baroffio, C.; Fountain, M.; Cross, J.; Wibe, A.; et al. Composition of strawberry floral volatiles and their effects on behavior of strawberry blossom weevil, Anthonomus rubi. J. Chem. Ecol. 2020, 46, 1069–1081. [Google Scholar] [CrossRef]
- Ceuppens, B.; Ameye, M.; Van Langenhove, H.; Roldan-Ruiz, I.; Smagghe, G. Characterization of volatiles in strawberry varieties ‘Elsanta’ and ‘Sonata’ and their effect on bumblebee flower visiting. Arthropod-Plant Interact. 2015, 9, 281–287. [Google Scholar] [CrossRef]
- Henning, J.A.; Teuber, L.R. Combined gas chromatography-electroantennogram characterization of alfalfa floral volatiles recognized by honey bees (Hymenoptera: Apidae). J. Econ. Entomol. 1992, 85, 226–232. [Google Scholar] [CrossRef]
- Morse, A.; Kevan, P.; Shipp, L.; Khosla, S.; McGarvey, B. The impact of greenhouse tomato (Solanales: Solanaceae) floral volatiles on bumble bee (Hymenoptera: Apidae) pollination. Environ. Entomol. 2012, 41, 855–864. [Google Scholar] [CrossRef]
- Chess, S.K.R.; Raguso, R.A.; LeBuhn, G. Geographic divergence in floral morphology and scent in Linanthus dichotomus (Polemoniaceae). Am. J. Bot. 2008, 95, 1652–1659. [Google Scholar] [CrossRef]
- Hamilton-Kemp, T.R.; Loughrin, J.H.; Andersen, R.A. Identification of some volatile compounds from strawberry flowers. Phytochemistry 1990, 29, 2847–2848. [Google Scholar] [CrossRef]
- Klatt, B.K.; Burmeister, C.; Westphal, C.; Tscharntke, T.; von Fragstein, M. Flower volatiles, crop varieties and bee responses. PLoS ONE 2013, 8, e72724. [Google Scholar] [CrossRef]
- Soler, C.; Hossaert-McKey, M.; Buatois, B.; Bessière, J.; Schatz, B.; Proffit, M. Geographic variation of floral scent in a highly specialized pollination mutualism. Phytochemistry 2011, 72, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Adler, L.S.; Irwin, R.E. What you smell is more important than what you see? Natural selection on floral scent. New Phytol. 2012, 195, 510–511. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Saona, C.; Parra, L.; Quiroz, A.; Isaacs, R. Variation in highbush blueberry floral volatile profiles as a function of pollination status, cultivar, time of day and flower part: Implications for flower visitation by bees. Ann. Bot. 2011, 107, 1377–1390. [Google Scholar] [CrossRef]
- Balakrishnan, K.; Holighaus, G.; Weißbecker, B.; Schütz, S. Electroantennographic responses of red flour beetle Tribolium castaneum Herbst (Coleoptera: Tenebrionidae) to volatile organic compounds. J. Appl. Entomol. 2016, 141, 477–486. [Google Scholar] [CrossRef]
- Bichão, H.; Borg-Karlson, A.; Araújo, J.; Mustaparta, H. Five types of olfactory receptor neurons in the strawberry blossom weevil Anthonomus rubi: Selective responses to inducible host-plant volatiles. Chem. Senses 2005, 30, 153–170. [Google Scholar] [CrossRef] [Green Version]
- Ashman, T.L.; Bradburn, M.; Cole, D.H.; Blaney, B.H.; Raguso, R.A. The Scent of a male: The role of floral volatiles in pollination of a gender dimorphic plant. Ecology 2005, 86, 2099–2105. [Google Scholar] [CrossRef] [Green Version]
- Wibe, A.; Borg-Karlson, A.; Cross, J.; Bichão, H.; Fountain, M.; Liblikas, I.; Sigsgaard, L. Combining 1,4-dimethoxybenzene, the major flower volatile of wild strawberry Fragaria vesca, with the aggregation pheromone of the strawberry blossom weevil Anthonomus rubi improves attraction. Crop Prot. 2014, 64, 122–128. [Google Scholar] [CrossRef]
- Blažytė-Čereškienė, L.; Apšegaitė, V.; Būda, V. Flower VOCs’ perception: Specialization sign in generalist pollinator. In Proceedings of the Joint Meeting of the 33rd Annual Meeting of the ISCE and The 9th Meeting of APACE, Kyoto, Japan, 23–27 August 2017; p. 194. [Google Scholar]
- Carlsson, M.A.; Hansson, B.S. Detection and coding of flower volatiles in nectar-foraging insects. In Biology of Floral Scent; Dudareva, N., Pichersky, E., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 243–261. [Google Scholar]
- Dobson, H.E.M. Relationship between floral fragrance composition and type of pollinator. In Biology of Floral Scent; Dudareva, N., Pichersky, E., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 147–198. [Google Scholar]
- Jürgens, A.; Witt, T.; Gottsberger, G. Flower scent composition in Dianthus and Saponaria species (Caryophyllaceae) and its relevance for pollination biology and taxonomy. Biochem. Syst. Ecol. 2003, 31, 345–357. [Google Scholar] [CrossRef]
- Knudsen, J.T.; Tollsten, L.; Bergström, L.G. Floral scents-A checklist of volatile compounds isolated by head-space techniques. Phytochemistry 1993, 33, 253–280. [Google Scholar] [CrossRef]
- Blight, M.M.; Le Métayer, M.; Delègue, M.P.; Pickett, J.A.; Marion-Poll, F.; Wadhams, L.J. Identification of floral volatiles involved in recognition of oilseed rape flowers, Brassica napus by honeybees, Apis mellifera. J. Chem. Ecol. 1997, 23, 1715–1727. [Google Scholar] [CrossRef]
- Theis, N. Fragrance of Canada thistle (Cirsium arvense) attracts both floral herbivores and pollinators. J. Chem. Ecol. 2006, 32, 917–927. [Google Scholar] [CrossRef] [PubMed]
- Wadhams, L.J.; Blight, M.M.; Kerguelen, V.; Le Métayer, M.; Marion-Poll, F.; Masson, C.; Pham-Delègue, M.H.; Woodcock, C.M. Discrimination of oilseed rape volatiles by honey bee: Novel combined gas chromatographic-electrophysiological behavioral assay. J. Chem. Ecol. 1994, 20, 3221–3231. [Google Scholar] [CrossRef]
- Williams, N.H.; Whitten, W.M. Orchid floral fragrances and male euglossine bees: Methods and advances in the last sesquidecade. Biol. Bull. 1983, 164, 355–395. [Google Scholar] [CrossRef]
- Dobson, H.E.M.; Danielson, E.M.; Wesep, I.D.V. Pollen odor chemicals as modulators of bumble bee foraging on Rosa rugosa Thunb. (Rosaceae). Plant Species Biol. 1999, 14, 153–166. [Google Scholar] [CrossRef]
- Ladd, T.L.; McGovern, T.P.; Beroza, M. Attraction of bumble bees and honey bees to traps baited with lures for the Japanese beetle. J. Econ. Entomol. 1974, 67, 307–308. [Google Scholar] [CrossRef]
- Huang, M.; Sanchez Moreiras, A.M.; Abel, C.; Sohrabi, R.; Lee, S.; Gershenzon, J.; Tholl, D. The major volatile organic compound emitted from Arabidopsis thaliana flowers, the sesquiterpene (E)-β-caryophyllene, is a defense against a bacterial pathogen. New Phytol. 2012, 193, 997–1008. [Google Scholar] [CrossRef]
- Junker, R.R.; Blüthgen, N. Floral scents repel facultative flower visitors, but attract obligate ones. Ann. Bot. 2010, 105, 777–782. [Google Scholar] [CrossRef] [Green Version]
- Kessler, D.; Diezel, C.; Clark, D.G.; Colquhoun, T.A.; Baldwin, I.T. Petunia flowers solve the defence/apparency dilemma of pollinator attraction by deploying complex floral blends. Ecol. Lett. 2013, 16, 299–306. [Google Scholar] [CrossRef]
- Chandra, S.; Smith, B.H. An analysis of synthetic processing of odor mixtures in the honeybee (Apis mellifera). J. Exp. Biol. 1998, 201, 3113–3121. [Google Scholar] [CrossRef] [PubMed]
- Laloi, D.; Sandoz, J.C.; Picard-Nizou, A.L.; Marchesi, A.; Pouvreau, A.; Tasei, J.N.; Poppy, G.; Pham-delègue, M.H. Olfactory conditioning of the proboscis extension in bumble bees. Entomol. Exp. Appl. 1999, 90, 123–129. [Google Scholar] [CrossRef]
- Kost, C.; Heil, M. Herbivore-induced plant volatiles induce an indirect defence in neighbouring plants. J. Ecol. 2006, 94, 619–628. [Google Scholar] [CrossRef]
- Knudsen, J.T.; Eriksson, R.; Gershenzon, J.; Ståhl, B. Diversity and distribution of floral scent. Bot. Rev. 2006, 72, 1. [Google Scholar] [CrossRef]
- Zhang, H.; Shan, S.; Gu, S.; Huang, X.; Li, Z.; Khashaveh, A.; Zhang, Y. Prior experience with food reward influences the behavioral responses of the honeybee Apis mellifera and the bumblebee Bombus lantschouensis to tomato floral scent. Insects 2020, 11, 884. [Google Scholar] [CrossRef]
- Pham-Delegue, M.H.; Etievant, P.; Guichard, E.; Masson, C. Sunflower volatiles involved in honeybee discrimination among genotypes and flowering stages. J. Chem. Ecol. 1989, 15, 329–343. [Google Scholar] [CrossRef]
- Wright, G.A.; Skinner, B.D.; Smith, B.H. Ability of honeybee, Apis mellifera, to detect and discriminate odors of varieties of canola (Brassica rapa and Brassica napus) and snapdragon flowers (Antirrhinum majus). J. Chem. Ecol. 2002, 28, 721–740. [Google Scholar] [CrossRef]
- Sachse, S.; Galizia, C.G. The coding of odour-intensity in the honeybee antennal lobe: Local computation optimizes odour representation. Eur. J. Neurosci. 2003, 18, 2119–2132. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Purity (%) | Origin |
|---|---|---|
| Ethyl benzoate | >99.5 | Aladdin 1 |
| Methyl salicylate | ≥99.5 | Aladdin |
| Ethyl salicylate | 99.0 | Aladdin |
| Ethyl palmitate | ≥99.0 | Aladdin |
| (Z)-3-Hexenyl acetate | 98.0 | Aladdin |
| (Z)-Hex-3-enyl benzoate | 97.0 | Aladdin |
| 2,2,4-Trimethyl-1,3-pentadienol diisobutyrate | 98.5 | Aladdin |
| Ethyl cinnamate | 99.0 | Macklin 2 |
| Cis-3-Hexenyl isovalerate | 97.0 | Aladdin |
| Ethyl myristate | ≥98.0 | Aladdin |
| (Z)-3-Hexenyl propionate | 97.0 | Macklin |
| Ethyl dodecanoate | 99.0 | Aladdin |
| Staflex BOP | ≥95.0 | Macklin |
| Benzyl benzoate | >99.0 | Aladdin |
| Linalool | 98.0 | Aladdin |
| Benzyl alcohol | ≥99.5 | Aladdin |
| Phenylethyl alcohol | ≥99.5 | Aladdin |
| Nonadecane | 98.0 | Macklin |
| 1-Chlorododecane | 98.0 | Aladdin |
| 1-Chlorotetradecane | 98.0 | Aladdin |
| β-Ionone | 97.0 | Aladdin |
| Methyleugenol | 98.0 | Aladdin |
| 1,3-Ditert-butylbenzene | >98.0 | Aladdin |
| Eugenol | 99.0 | Aladdin |
| Melonal | 80.0 | Aladdin |
| Benzeneacetaldehyde | 95.0 | Aladdin |
| Types/No. | Compounds | CAS Number | RT | Relative Content (%) |
|---|---|---|---|---|
| Alcohols | ||||
| 1 | Linalool | 78-70-6 | 13.601 | 9.12 |
| 2 | Benzyl alcohol | 110-51-6 | 12.246 | 1.23 |
| 3 | Phenylethyl alcohol | 60-12-8 | 17.134 | 0.05 |
| 4 | Erythro-1-phenyl-1,2-propanediol | 1075-04-3 | 13.01 | 0.02 |
| 5 | Bicyclo[3.3.1]non-2-en-9-ol, syn- | 19877-78-2 | 17.967 | 0.34 |
| 6 | Ledol | 577-27-5 | 31.641 | 0.16 |
| Esters | ||||
| 7 | Ethyl benzoate | 93-89-0 | 16.412 | 6.27 |
| 8 | Methyl salicylate | 119-36-8 | 17.429 | 3.08 |
| 9 | Ethyl salicylate | 118-61-6 | 21.599 | 2.62 |
| 10 | Ethyl palmitate | 628-97-7 | 36.995 | 1.31 |
| 11 | (Z)-3-Hexenyl acetate | 3681-71-8 | 9.71 | 1.23 |
| 12 | (Z)-3-Hexenyl benzoate | 25152-85-6 | 30.292 | 0.77 |
| 13 | 2,2,4-Trimethyl-1,3-pentadienol diisobutyrate | 6846-50-0 | 30.722 | 0.67 |
| 14 | Tris ethyl cinnamate | 4192-77-2 | 28.229 | 0.43 |
| 15 | Cis-3-Hexenyl isovalerate | 35154-45-1 | 19.565 | 0.36 |
| 16 | Ethyl myristate | 124-06-1 | 33.694 | 0.36 |
| 17 | (Z)-3-Hexenyl propionate | 33467-74-2 | 19.368 | 0.31 |
| 18 | Ethyl dodecanoate | 106-33-2 | 30.653 | 0.25 |
| 19 | Staflex BOP | 84-78-6 | 34.859 | 0.13 |
| 20 | Benzyl benzoate | 120-51-4 | 33.452 | 0.09 |
| 21 | Benzyl valerate | 10361-39-4 | 26.434 | 0.23 |
| Alkanes | ||||
| 22 | Nonadecane | 629-92-5 | 39.455 | 1.72 |
| 23 | 1-Chlorododecane | 112-52-7 | 21.133 | 0.55 |
| 24 | 1-Chlorotetradecane | 2425-54-9 | 28.77 | 0.35 |
| 25 | Heptacosane | 593-49-7 | 42.332 | 0.84 |
| 26 | 1-Chloro-5-methyl hexane | 33240-56-1 | 24.997 | 0.02 |
| Ketones | ||||
| 27 | N-Succinimidyl benzoate | 23405-15-4 | 8.191 | 2.79 |
| 28 | β-Ionone | 79-77-6 | 28.632 | 0.91 |
| Aromatics | ||||
| 29 | Methyleugenol | 93-15-2 | 26.818 | 0.33 |
| 30 | 1,3-Ditert-butylbenzene | 1014-60-4 | 20.464 | 4.11 |
| 31 | Eugenol | 97-53-0 | 25.42 | 9.65 |
| 32 | Butylated hydroxytoluene (BHT) | 128-37-0 | 29.183 | 0.23 |
| 33 | 2,4-Ditert-butylphenol | 96-76-4 | 29.338 | 1.42 |
| Aldehydes | ||||
| 34 | Melonal | 106-72-9 | 4.98 | 21.25 |
| 35 | Benzeneacetaldehyde | 122-78-1 | 11.199 | 2.96 |
| 36 | 1,3,4-Trimethyl-3-cyclohexene-1-carboxaldehyde | 40702-26-9 | 18.64 | 0.57 |
| 35 | (Z)-7-Hexadecenal | 56797-40-1 | 32.3 | 0.13 |
| Ethers | ||||
| 38 | Dimethyl ether | 115-10-6 | 1.798 | 23.14 |
| Mean EAG Response to Different Compounds (%, Diluted in Liquid Paraffin) | |||||||
|---|---|---|---|---|---|---|---|
| Bee Species/Compounds | 10 μg/μL | 100 μg/μL | 200 μg/μL | 300 μg/μL | 400 μg/μL | 500 μg/μL | |
| A. mellifera | Ethyl benzoate | 8.94 ± 0.19 b | 45.41 ± 0.27 a | 23.05 ± 0.18 b | 15.92 ± 0.16 b | 33.04 ± 0.15 ab | 6.24 ± 0.11 b |
| Ethyl salicylate | 25.89 ± 0.81 b | 38.66 ± 0.27 b | 57.39 ± 0.17 ab | 53.13 ± 0.10 ab | 179.38 ± 0.75 a | 218.25 ± 0.81 a | |
| Ethyl palmitate | 8.12 ± 0.01 c | 6.29 ± 0.02 c | 14.64 ± 0.05 bc | 14.04 ± 0.03 bc | 30.54 ± 0.10 ab | 37.02 ± 0.03 a | |
| (Z)-3-Hexenyl acetate | 41.84 ± 0.12 b | 213.12 ± 0.10 a | 219.15 ± 0.27 a | 142.29 ± 0.48 ab | 145.00 ± 0.50 ab | 70.86 ± 0.17 ab | |
| Tris ethyl cinnamate | 15.24 ± 0.04 b | 25.38 ± 0.08 b | 23.86 ± 0.02 b | 9.08 ± 0.17 b | 47.16 ± 0.05 a | 13.89 ± 0.04 b | |
| Cis-3-Hexenyl isovalerate | 4.32 ± 0.01 b | 37.30 ± 0.09 a | 36.25 ± 0.09 a | 17.18 ± 0.08 ab | 19.86 ± 0.05 ab | 19.19 ± 0.13 ab | |
| Ethyl myristate | 12.49 ± 0.03 c | 12.67 ± 0.01 c | 20.03 ± 0.08 bc | 28.08 ± 0.11 bc | 63.34 ± 0.03 a | 43.56 ± 0.07 ab | |
| (Z)-3-Hexenyl propionate | 27.10 ± 0.06 b | 75.84 ± 0.29 ab | 59.35 ± 0.16 ab | 116.10 ± 0.25 a | 103.27 ± 0.31 ab | 96.34 ± 0.23 ab | |
| Ethyl dodecanoate | 51.72 ± 0.10 a | 24.43 ± 0.07 b | 21.53 ± 0.06 b | 26.20 ± 0.08 b | 14.12 ± 0.02 b | 15.25 ± 0.01 b | |
| Butyl octyl phthalate | 17.23 ± 0.08 bc | 14.73 ± 0.04 c | 24.27 ± 0.04 abc | 41.65 ± 0.07 ab | 45.44 ± 0.10 a | 32.89 ± 0.15 abc | |
| Linalool | 37.06 ± 0.05 ab | 30.06 ± 0.07 b | 70.78 ± 0.17 a | 56.78 ± 0.13 ab | 41.52 ± 0.08 ab | 42.98 ± 0.03 ab | |
| Benzyl alcohol | 34.64 ± 0.10 b | 34.10 ± 0. 04 b | 23.96 ± 0.12 b | 142.58 ± 0.22 a | 25.54 ± 0.03 b | 30.36 ± 0.14 b | |
| Phenylethyl alcohol | 48.28 ± 0.08 ab | 30.00 ± 0.03 b | 64.85 ± 0.16 ab | 37.66 ± 0.07 ab | 73.29 ± 0.08 a | 59.06 ± 0.16 ab | |
| 1-Chlorododecane | 41.46 ± 0.02 c | 17.42 ± 0.04 d | 24.51 ± 0.05 cd | 45.37 ± 0.02 bc | 62.96 ± 0.10 b | 118.77 ± 0.11 a | |
| 1-Chlorotetradecane | 26.08 ± 0.07 c | 62.02 ± 0.05 a | 39.81 ± 0.01 bc | 27.42 ± 0.05 c | 54.90 ± 0.05 ab | 32.22 ± 0.06 c | |
| β-Ionone | 30.20 ± 0.03 b | 10.51 ± 0.06 b | 51.23 ± 0.01 b | 127.68 ± 0.04 a | 56.07 ± 0.27 b | 42.24 ± 0.18 b | |
| Methyleugenol | 3.35 ± 0.01 b | 45.42 ± 0.04 a | 31.76 ± 0.09 ab | 35.05 ± 0.17 ab | 28.48 ± 0.10 ab | 27.12 ± 0.09 ab | |
| 1,3-Di-tert-butylbenzene | 8.12 ± 0.02 b | 25.09 ± 0.02 ab | 22.26 ± 0.03 ab | 35.41 ± 0.06 a | 24.99 ± 0.05 ab | 31.24 ± 0.12 a | |
| B. terrestris | Ethyl palmitate | 50.03 ± 0.04 ab | 71.82 ± 0.09 a | 34.50 ± 0.22 ab | 29.06 ± 0.11 ab | 27.79 ± 0.10 ab | 15.07 ± 0.07 b |
| (Z)-3-Hexenyl acetate | 27.42 ± 0.13 b | 84.36 ± 0.37 ab | 142.34 ± 0.62 ab | 108.13 ± 0.45 ab | 124.85 ± 0.75 ab | 208.97 ± 0.22 a | |
| Cis-3-Hexenyl isovalerate | 5.51 ± 0.05 c | 20.76 ± 0.11 c | 42.26 ± 0.10 b | 31.28 ± 0.08 bc | 31.28 ± 0.06 bc | 79.79 ± 0.13 a | |
| Ethyl myristate | 5.88 ± 0.02 b | 7.96 ± 0.03 b | 24.03 ± 0.06 ab | 23.45 ± 0.10 ab | 13.48 ± 0.03 ab | 69.02 ± 0.30 a | |
| Benzyl benzoate | 11.66 ± 0.03 b | 63.38 ± 0.27 a | 35.92 ± 0.12 ab | 7.53 ± 0.03 b | 22.36 ± 0.13 ab | 36.93 ± 0.05 ab | |
| Linalool | 42.10 ± 0.08 ab | 21.80 ± 0.11 ab | 26.96 ± 0.21 ab | 1.56 ± 0.01 b | 43.61 ± 0.07 ab | 84.96 ± 0.40 a | |
| Benzyl alcohol | 19.71 ± 0.05 b | 28.22 ± 0.03 b | 79.08 ± 0.42 ab | 71.68 ± 0.56 ab | 227.13 ± 0.91 a | 61.31 ± 0.27 b | |
| Phenylethyl alcohol | 3.75 ± 0. 01 b | 69.68 ± 0.24 ab | 74.68 ± 0.28 ab | 41.26 ± 0.15 ab | 71.53 ± 0.31 ab | 92.33 ± 0.25 a | |
| 1-Chlorotetradecane | 44.44 ± 0.05 a | 32.53 ± 0.13 ab | 49.11 ± 0.10 a | 23.66 ± 0.05 ab | 43.85 ± 0.14 a | 6.95 ± 0.01 b | |
| 1,3-Di-tert-butylbenzene | 45.44 ± 0.05 ab | 28.22 ± 0.08 b | 27.26 ± 0.11 b | 62.51 ± 0.08 a | 50.49 ± 0.09 ab | 59.10 ± 0.10 a | |
| Eugenol | 33.08 ± 0.07 b | 47.28 ± 0.07 b | 47.28 ± 0.07 b | 81.37 ± 0.21 ab | 57.88 ± 0.07 ab | 105.66 ± 0.24 a | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Chen, M.; Ma, W.; Zheng, L.; Zhang, B.; Zhao, H.; Jiang, Y. Composition of Strawberry Flower Volatiles and Their Effects on Behavior of Strawberry Pollinators, Bombus terrestris and Apis mellifera. Agronomy 2023, 13, 339. https://doi.org/10.3390/agronomy13020339
Liu J, Chen M, Ma W, Zheng L, Zhang B, Zhao H, Jiang Y. Composition of Strawberry Flower Volatiles and Their Effects on Behavior of Strawberry Pollinators, Bombus terrestris and Apis mellifera. Agronomy. 2023; 13(2):339. https://doi.org/10.3390/agronomy13020339
Chicago/Turabian StyleLiu, Jinjia, Min Chen, Weihua Ma, Lifang Zheng, Bing Zhang, Huiting Zhao, and Yusuo Jiang. 2023. "Composition of Strawberry Flower Volatiles and Their Effects on Behavior of Strawberry Pollinators, Bombus terrestris and Apis mellifera" Agronomy 13, no. 2: 339. https://doi.org/10.3390/agronomy13020339
APA StyleLiu, J., Chen, M., Ma, W., Zheng, L., Zhang, B., Zhao, H., & Jiang, Y. (2023). Composition of Strawberry Flower Volatiles and Their Effects on Behavior of Strawberry Pollinators, Bombus terrestris and Apis mellifera. Agronomy, 13(2), 339. https://doi.org/10.3390/agronomy13020339