


Physio-Chemical and Agronomic-Based Characterization of Synthetic Hexaploid Wheat Germplasm under Field Imposed Conditions of Drought and Heat Stress


Abstract
:1. Introduction
2. Material and Methods
2.1. Crop Husbandry, Treatments, and Data Collection
2.2. Estimation of Physiological Traits
2.3. Estimation of Biochemical Traits
2.4. Estimation of Agronomic Traits
2.5. Statistical Analysis
3. Results
3.1. Physiological Traits
3.2. Biochemical Traits
3.3. Agronomic Traits
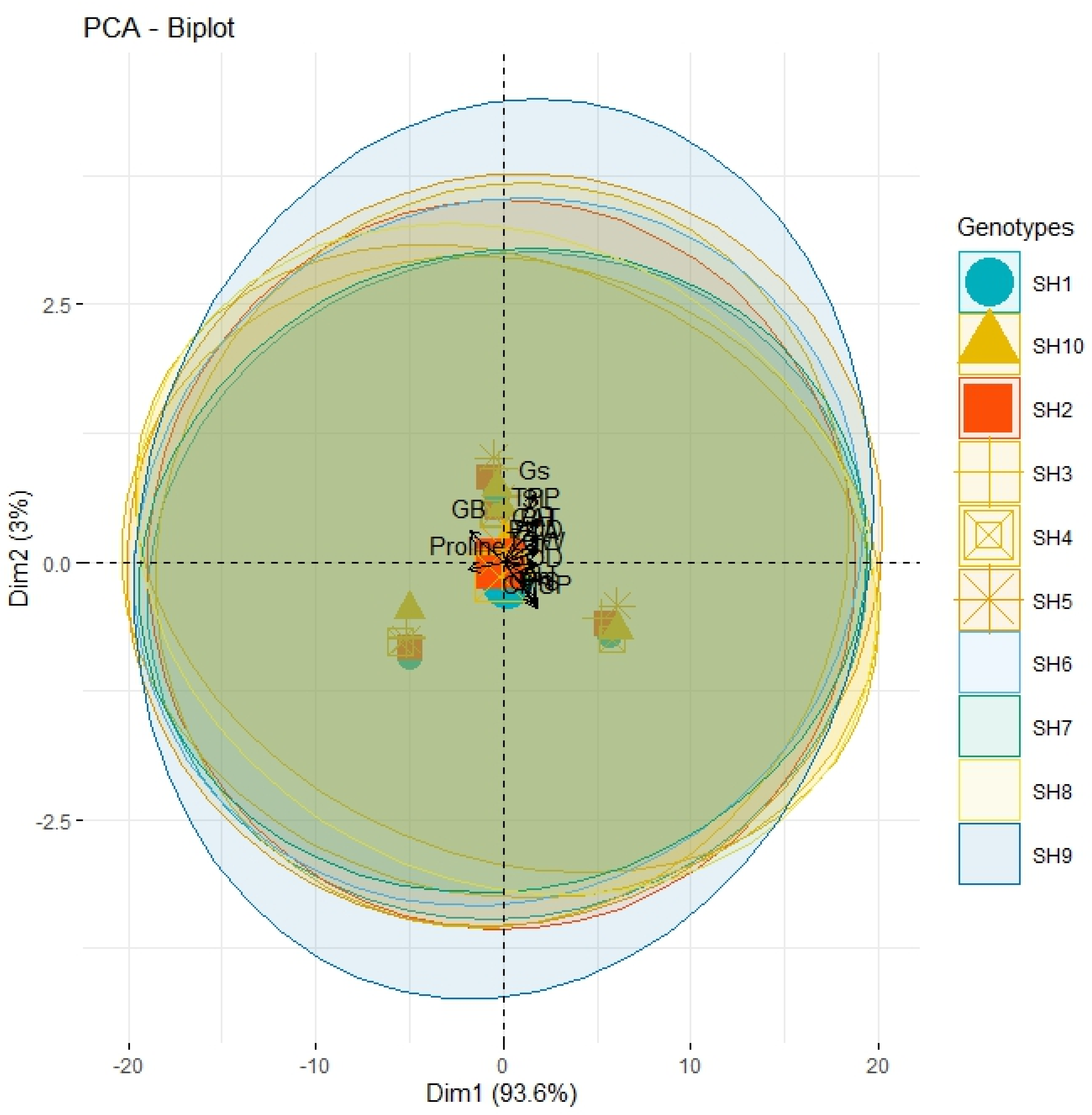
3.4. Correlation, PCA and Heatmap Analysis
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Arora, N.K. Impact of climate change on agriculture production and its sustainable solutions. Environ. Sustain. 2019, 2, 95–96. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop Production under Drought and Heat Stress: Plant Responses and Management Options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought Stress Impacts on Plants and Different Approaches to Alleviate Its Adverse Effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef]
- Chaudhary, S.; Devi, P.; Bhardwaj, A.; Jha, U.C.; Sharma, K.D.; Prasad, P.V.V.; Siddique, K.H.M.; Bindumadhava, H.; Kumar, S.; Nayyar, H. Identification and Characterization of Contrasting Geno-types/Cultivars for Developing Heat Tolerance in Agricultural Crops: Current Status and Prospects. Front. Plant Sci. 2020, 11, 587264. [Google Scholar] [CrossRef] [PubMed]
- Sattar, A.; Sher, A.; Ijaz, M.; Ul-Allah, S.; Rizwan, M.S.; Hussain, M.; Jabran, K.; Cheema, M. Terminal drought and heat stress alter physiological and biochemical attributes in flag leaf of bread wheat. PLoS ONE 2020, 15, e0232974. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Narayanan, S.; Erdayani, E.; Prasad, P.V.V. Effects of high temperature stress during anthesis and grain filling periods on photosynthesis, lipids and grain yield in wheat. BMC Plant Biol. 2020, 20, 268. [Google Scholar] [CrossRef]
- Dwivedi, S.K.; Basu, S.; Kumar, S.; Kumar, G.; Prakash, V.; Kumar, S.; Mishra, J.S.; Bhatt, B.; Malviya, N.; Singh, G.; et al. Heat stress induced impairment of starch mobilisation regulates pollen viability and grain yield in wheat: Study in Eastern Indo-Gangetic Plains. Field Crops Res. 2017, 206, 106–114. [Google Scholar] [CrossRef]
- Alghabari, F.; Shah, Z.H.; Elfeel, A.A.; Alyami, J.H. Biochemical and Physiological Responses of Thermostable Wheat Genotypes for Agronomic Yield under Heat Stress during Reproductive Stages. Agronomy 2021, 20, 80. [Google Scholar] [CrossRef]
- dos Santos, T.B.; Ribas, A.F.; de Souza, S.G.H.; Budzinski, I.G.F.; Domingues, D.S. Physiological Responses to Drought, Salinity, and Heat Stress in Plants: A Review. Stresses 2022, 2, 113–135. [Google Scholar] [CrossRef]
- Shah, Z.H.; Rehman, H.M.; Akhtar, T.; Daur, I.; Nawaz, M.A.; Ahmad, M.Q.; Rana, I.A.; Atif, R.M.; Yang, S.H.; Chung, G. Redox and Ionic Homeostasis Regulations against Oxidative, Salinity and Drought Stress in Wheat (A Systems Biology Approach). Front Genet. 2017, 8, 141. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Lu, M.; Wang, Y.; Wang, Y.; Liu, Z.; Chen, S. Response Mechanism of Plants to Drought Stress. Horticulturae 2021, 7, 50. [Google Scholar] [CrossRef]
- Qayyum, A.; Al Ayoubi, S.; Sher, A.; Bibi, Y.; Ahmad, S.; Shen, Z.; Jenks, M.A. Improvement in drought tolerance in bread wheat is related to an improvement in osmolyte production, antioxidant enzyme activities, and gaseous exchange. Saudi J. Biol. Sci. 2021, 28, 5238–5249. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.P.; Zhang, X.Y.; Li, F.; Luo, Y.; Wang, W. Overaccumulation of glycine betaine enhances tolerance to drought and heat stress in wheat leaves in the protection of photosynthesis. Photosynthetica 2010, 48, 117–126. [Google Scholar] [CrossRef]
- Pour-Aboughadareh, A.; Omidi, M.; Naghavi, M.R.; Etminan, A.; Mehrabi, A.A.; Poczai, P.; Bayat, H. Effect of Water Deficit Stress on Seedling Biomass and Physio-Chemical Characteristics in Different Species of Wheat Possessing the D Genome. Agronomy 2019, 9, 522. [Google Scholar] [CrossRef]
- Annunziata, M.G.; Ciarmiello, L.F.; Woodrow, P.; Dell’Aversana, E.; Carillo, P. Spatial and temporal profile of glycine betaine accumulation in plants under abiotic stresses. Front. Plant Sci. 2019, 10, 230. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Aslam, Z.; Javed, T.; Hussain, S.; Raza, A.; Shabbir, R.; Mora-Poblete, F.; Saeed, T.; Zulfiqar, F.; Ali, M.M.; et al. Screening of Wheat (Triticum aestivum L.) Genotypes for Drought Tolerance through Agronomic and Physiological Response. Agronomy 2022, 12, 287. [Google Scholar] [CrossRef]
- Shah, S.M.D.M.; Shabbir, G.; Malik, S.I.; Raja, N.I.; Shah, Z.H.; Rauf, M.; Zahrani, Y.A.; Alghabari, F.; Alsamadany, H.; Shahzad, K.; et al. Delineation of Physiological, Agronomic and Genetic Responses of Different Wheat Genotypes under Drought Condition. Agronomy 2022, 12, 1056. [Google Scholar] [CrossRef]
- Abhinandan, K.; Skori, L.; Stanic, M.; Hickerson, N.M.; Jamshed, M.; Samuel, M.A. Abiotic stress signaling in wheat–an inclusive overview of hormonal interactions during abiotic stress responses in wheat. Front. Plant Sci. 2018, 9, 734. [Google Scholar] [CrossRef]
- Li, A.; Liu, D.; Yang, W.; Kishii, M.; Mao, L. Synthetic hexaploid wheat: Yesterday, today, and tomorrow. Engineering 2018, 4, 552–558. [Google Scholar] [CrossRef]
- Pradhan, G.P.; Prasad, P.V.; Fritz, A.K.; Kirkham, M.B.; Gill, B.S. Effects of drought and high temperature stress on synthetic hexaploid wheat. Funct. Plant Biol. 2012, 39, 190–198. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Boyle, D.L.; Welti, R.; Jagadish, S.V.K.; Prasad, P.V.V. Decreased photosynthetic rate under high temperature in wheat is due to lipid desaturation, oxidation, acylation, and damage of organelles. BMC Plant Biol. 2018, 8, 55. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Shayeganpour, A.; Brocks, D.R.; Lavasanifar, A.; Samuel, J. High-performance liquid chromatography analysis of curcumin in rat plasma: Application to pharmacokinetics of polymeric micellar formulation of curcumin. Biomed. Chrom. 2007, 21, 546–552. [Google Scholar] [CrossRef]
- Yadav, M.R.; Choudhary, M.; Singh, J.; Lal, M.K.; Jha, P.K.; Udawat, P.; Gupta, N.K.; Rajput, V.D.; Garg, N.K.; Maheshwari, C.; et al. Impacts, Tolerance, Adaptation, and Mitigation of Heat Stress on Wheat under Changing Climates. Int. J. Mol. Sci. 2022, 23, 2838. [Google Scholar] [CrossRef] [PubMed]
- Qaseem, M.F.; Qureshi, R.; Shaheen, H. Effects of Pre-Anthesis Drought, Heat and Their Combination on the Growth, Yield and Physiology of diverse Wheat (Triticum aestivum L.) Genotypes Varying in Sensitivity to Heat and drought stress. Sci. Rep. 2019, 9, 6955. [Google Scholar] [CrossRef]
- Caverzan, A.; Casassola, A.; Brammer, S.P. Antioxidant responses of wheat plants under stress. Genet. Mol. Biol. 2016, 39, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Qamer, Z.; Chaudhary, M.T.; Du, X.; Hinze, L.; Azhar, M.T. Review of oxidative stress and antioxidative defense mechanisms in Gossypium hirsutum L. in response to extreme abiotic conditions. J. Cotton Res. 2021, 4, 1–9. [Google Scholar] [CrossRef]
- Nasirzadeh, L.; Sorkhilaleloo, B.; Majidi Hervan, E.; Fatehi, F. Changes in antioxidant enzyme activities and gene expression profiles under drought stress in tolerant, intermediate, and susceptible wheat genotypes. Cereal Res. Commun. 2021, 49, 83–89. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Pedigree |
|---|---|
| SH1 | 68.111/RGB-U//WARD/3/FGO/4/RABI/5/AE.SQUARROSA (882)/6/ATTILA/10 |
| SH2 | RABE/2*MO88/3/CAZO/KAUZ//KAUZ |
| SH3 | URES/JUN//KAUZ/3/ALTAR 84/AE. SQ//2*OPATA |
| SH4 | CROC_1/AE.SQUARROSA (205)//KAUZ/3/ENEIDA/4/FINSI |
| SH5 | FILIN/IRENA/5/CNDO/R143//ENTE/MEXI-2/3/AEGILOPS SQUAROSA (TAUS)/4/WEAVER/6/ |
| SH6 | FILI/IRENA/5/CNDO/R143//ENTE/MEXI_2/3/AEGILOPS SQUAROSA (TAUS)/4/WEAVER/6/BERKUT |
| SH7 | FILI/IRENA/5/CNDO/R143//ENTE/MEXI_2/3/AEGILOPS SQUAROSA (TAUS)/4/WEAVER/6/BERKUT |
| SH8 | FILIN/3/CROC_1/AE. SQUAROSA (205)//KAUZ/4/FILIN/5/VEE/MJI//2* TUI/3/PASTOR |
| SH9 | CROC_1/AE. SQUAROSA (205)//KAUZ/3/PRL/SARA//TSI/VEE#5 |
| SH10 | QT8343//PASTOR*2/OPATA |
| Months | Temperature (°C) | Precipitation (mm) | Humidity (%) | Rainy Days | Average Sun Hours | ||
|---|---|---|---|---|---|---|---|
| Min | Max | Avg | |||||
| October | 25.5 | 34 | 29.6 | 3 | 65 | 0 | 10.4 |
| November | 23.4 | 31 | 27.1 | 26 | 63 | 2 | 9.7 |
| December | 21.2 | 28.5 | 24.7 | 15 | 61 | 2 | 9.1 |
| January | 19.4 | 26.8 | 23 | 25 | 59 | 2 | 8.7 |
| February | 19.8 | 28.2 | 23.9 | 2 | 60 | 1 | 9.8 |
| March | 20.7 | 29.8 | 25.2 | 4 | 59 | 1 | 10.6 |
| April | 22.9 | 32.2 | 27.6 | 4 | 57 | 0 | 11.3 |
| May | 25 | 34.8 | 30 | 0 | 55 | 0 | 11.8 |
| Treatments | Photosynthesis Rate (µmm−2S−1) | Stomatal Conductance (mmm−2S−1) | Transpiration Rate (mmm−2S−1) | Total Chlorophyll (g kg−1) | CMSP | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2020–21 | 2021–22 | 2020–21 | 2021–22 | 2020–21 | 2021–22 | 2020–21 | 2021–22 | 2020–21 | 2021–22 | |
| Stress Types | ||||||||||
| Control | 33.54 a | 35.82 a | 890.60 a | 920.00 a | 14.00 a | 16.50 a | 1.60 a | 1.68 a | 77 a | 80 a |
| Drought Heat Drought + heat | 23.63 b 24.55 c 18.00 d | 26.24 b 27.50 c 18.50 d | 797.25 b 790.50 c 605.00 d | 829.50 b 835.00 c 650.00 d | 11.25 b 10.00 c 7.50 d | 12.34 b 11.25 c 8.15 d | 0.87 b 0.82 c 0.50 d | 0.95 b 0.83 c 0.55 d | 50 b 52 c 46 d | 53 b 57 c 47 d |
| LSD value at 0.05 | 0.32 | 0.40 | 7.88 | 8.25 | 0.15 | 0.17 | 0.06 | 0.056 | 1.00 | 0.95 |
| Genotypes | ||||||||||
| SH1 | 27.25 | 28.61 | 825.55 | 835.60 | 13.15 | 13.78 | 1.45 | 1.54 | 24.25 | 25.00 |
| SH2 | 28.39 | 28.90 | 830.17 | 838.25 | 14.25 | 15.20 | 1.50 | 1.52 | 25.15 | 26.15 |
| SH3 | 30.50 | 30.60 | 866.48 | 856.40 | 14.75 | 15.23 | 1.56 | 1.58 | 27.25 | 25.69 |
| SH4 SH5 SH6 SH7 SH8 SH9 SH10 | 32.45 31.25 28.50 30.50 29.50 33.50 34.00 | 29.78 32.00 29.50 31.63 31.75 34.00 33.75 | 850.55 840.55 855.40 860.76 858.59 845.50 848.58 | 855.50 848.45 873.42 866.56 862.49 855.40 868.49 | 13.65 14.00 14.25 13.78 14.65 15.00 14.75 | 14.13 14.65 15.00 14.25 14.78 15.38 15.37 | 1.66 1.45 1.53 1.59 1.63 1.54 1.60 | 1.69 1.54 1.57 1.65 1.69 1.60 1.65 | 26.15 23.18 24.45 23.25 24.45 24.50 25.55 | 27.23 24.16 25.76 24.15 25.25 26.00 26.76 |
| LSD value at 0.05 | 2.30 | 1.85 | 7.46 | 3.70 | 0.95 | 0.93 | 0.05 | 0.06 | 1.60 | 1.72 |
| Significance | ||||||||||
| Stress types | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** |
| Genotype | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| Stress × genotype | * | * | * | ** | ** | ** | ** | ** | ** | ** |
| Treatments | Superoxide Dismutase (Enzyme Unit) | Peroxidase (Enzyme Unit) | Catalase (Enzyme Unit) | Glycine Betaine (μmol g−1 DW) | Proline (μg g−1 FW) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2020–21 | 2021–22 | 2020–21 | 2021–22 | 2020–21 | 2021–22 | 2020–21 | 2021–22 | 2020–21 | 2021–22 | |
| Stress Types | ||||||||||
| Control | 35.54 a | 37.82 a | 0.70 a | 0.75 a | 16.00 a | 16.50 a | 60 a | 63 a | 20.00 a | 21.16 a |
| Drought Heat Drought + heat | 25.63 b 24.20 c 18.15 d | 26.62 b 25.30 c 19.30 d | 0.50 b 0.45 c 0.21 d | 0..53 b 0.48 c 0.23 d | 12.00 b 11.50 c 7.00 d | 13.34 b 12.25 c 7.35 d | 140 b 130 c 170 d | 143 b 133 c 177 d | 32.15 b 30.25 c 40.15 d | 31.25 b 32.30 c 43.15 d |
| LSD value at 0.05 | 0.32 | 0.40 | 0.01 | 0.02 | 0.05 | 0.07 | 2.15 | 2.00 | 1.00 | 0.95 |
| Genotypes | ||||||||||
| SH1 | 33.23 | 35.15 | 0.73 | 0.75 | 15.25 | 15.30 | 147 | 150 | 24.25 | 25.00 |
| SH2 | 32.16 | 33.18 | 0.69 | 0.70 | 15.00 | 15.25 | 150 | 153 | 25.15 | 26.15 |
| SH3 | 30.71 | 31.10 | 0.65 | 0.68 | 15.17 | 15.23 | 153 | 158 | 27.25 | 25.69 |
| SH4 SH5 SH6 SH7 SH8 SH9 SH10 | 32.45 28.75 29.36 28.51 30.15 29.16 34.50 | 34.15 30.14 31.15 29.18 32.23 31.46 36.56 | 0.72 0.68 0.69 0.73 0.65 0.68 0.72 | 0.75 0.71 0.73 0.70 0.68 0.70 0.73 | 14.75 15.50 14.69 15.35 14.75 15.65 15.35 | 15.00 16.15 15.25 15.69 15.25 16.25 15.75 | 149 153 161 155 149 158 162 | 153 155 159 157 153 159 160 | 26.15 23.18 24.45 23.25 24.45 24.50 25.55 | 27.23 24.16 25.76 24.15 25.25 26.00 26.76 |
| LSD value at 0.05 | 1.30 | 0.32 | 0.05 | 0.06 | 1.13 | 0.13 | 0.05 | 0.06 | 1.60 | 1.72 |
| Significance | ||||||||||
| Stress types | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** |
| Genotype | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| Stress × genotype | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** |
| Treatments | Flag Leaf Area (cm2) | Plant Height (cm) | Tillers Plant−1 | Grains Spike−1 | Spike Length (cm) | Thousand Grain Weight (g) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2020–21 | 2021–22 | 2020–21 | 2021–22 | 2020–21 | 2021–22 | 2020–21 | 2021–22 | 2020–21 | 2021–22 | 2020–21 | 2021–22 | |
| Stress Types | ||||||||||||
| Control | 32 a | 35 a | 80 a | 81 a | 7.15 a | 8.15 a | 53 a | 54 a | 10.15 a | 10.75 a | 37 a | 38 a |
| Drought Heat Drought + heat | 24 b 25 c 14 d | 25 b 26 c 15 d | 75 b 73 c 63 d | 74 b 71 c 64 d | 5.25 b 6.10 c 3.00 d | 5.35 b 5.75 c 3.15 d | 44 b 42 c 38 d | 45 b 41 c 39 d | 9.15 b 8.50 c 5.50 d | 9.00 b 8.25 c 5.35 d | 33 b 31 c 27 d | 35 b 33 c 28 d |
| LSD value at 0.05 | 1.00 | 0.95 | 1.80 | 8.25 | 0.5 | 0.60 | 0.61 | 0.56 | 0.05 | 0.05 | 0.95 | 0.97 |
| Genotypes | ||||||||||||
| SH1 | 28 | 29 | 76 | 75 | 7.15 | 7.35 | 49 | 51 | 10.00 | 9.95 | 35 | 36 |
| SH2 | 29 | 31 | 77 | 76 | 7.35 | 7.45 | 50 | 49 | 9.75 | 10.00 | 34 | 37 |
| SH3 | 30 | 32 | 75 | 74 | 6.90 | 7.50 | 51 | 50 | 10.15 | 9.85 | 36 | 35 |
| SH4 SH5 SH6 SH7 SH8 SH9 SH10 | 29 32 31 33 29 31 33 | 30 29 32 31 30 32 31 | 74 73 76 75 77 78 75 | 77 74 75 77 74 77 75 | 6.85 7.00 7.15 6.90 6.85 7.10 8.00 | 7.00 7.15 7.50 7.00 7.15 7.50 7.85 | 53 52 50 49 50 51 48 | 52 53 52 51 49 52 51 | 9.85 10.25 9.85 10.15 10.35 10.30 10.40 | 10.15 10.40 10.35 9.85 10.15 9.95 10.25 | 37 35 38 37 34 36 37 | 36 38 35 36 37 35 36 |
| LSD value at 0.05 | 2.30 | 0.32 | 3.4 | 3.7 | 0.13 | 0.13 | 2.00 | 2.15 | 0.045 | 0.05 | 1.15 | 1.23 |
| Significance | ||||||||||||
| Stress types | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** |
| Genotype | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| Stress × genotype | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** |
| Stress Type | Geno. | Pn (µmm−2S−1) | Gs (mmm−2S−1) | Tr (mmm−2S−1) | Chl (gKg−1) | CMSP | Proline (μgg−1FW) | SOD (EU) | POD (EU) | CAT (EU) | GB (μmol g−1 DW) | FLA (cm2) | PH (cm) | TPP | GPS | SL (cm) | TGW (g) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | SH1 | 33 | 900 | 14 | 1.63 | 76 | 20 | 35 | 0.75 | 16 | 63 | 33 | 80 | 7.15 | 54 | 10.65 | 38 |
| SH2 | 32 | 910 | 15 | 1.65 | 77 | 21 | 33 | 0.73 | 15 | 61 | 34 | 81 | 7.35 | 53 | 10.35 | 37 | |
| SH3 | 31 | 890 | 14 | 1.67 | 78 | 22 | 32 | 0.71 | 16 | 64 | 31 | 79 | 7.60 | 52 | 10.45 | 37 | |
| SH4 SH5 SH6 SH7 SH8 SH9 SH10 | 34 32 33 34 32 31 34 | 895 920 917 905 888 879 886 | 15 16 13 14 15 16 15 | 1.60 1.67 1.59 1.63 1.64 1.68 1.69 | 80 79 77 78 79 76 79 | 20 21 19 23 21 21 22 | 31 34 32 33 36 35 37 | 0.75 0.76 0.72 0.75 0.71 0.73 0.75 | 17 18 16 16 15 17 17 | 65 62 61 62 60 63 61 | 35 32 33 35 33 33 34 | 80 81 80 82 81 80 79 | 7.00 7.45 7.25 7.65 7.55 7.70 7.80 | 54 52 51 53 53 52 51 | 10.55 10.35 10.25 10.58 11.00 10.35 10.85 | 38 39 38 37 39 38 39 | |
| Drought | SH1 | 23 | 800 | 11 | 0.95 | 53 | 32 | 25 | 0.53 | 13 | 145 | 24 | 73 | 5.50 | 53 | 9.15 | 33 |
| SH2 | 22 | 810 | 10 | 0.90 | 52 | 31 | 24 | 0.51 | 13 | 143 | 23 | 71 | 5.10 | 44 | 8.75 | 31 | |
| SH3 | 21 | 815 | 11 | 0.87 | 50 | 33 | 26 | 0.52 | 12 | 141 | 25 | 72 | 5.65 | 45 | 8.80 | 32 | |
| SH4 SH5 SH6 SH7 SH8 SH9 SH10 | 24 22 23 24 22 21 24 | 821 808 817 805 825 821 813 | 9 10 11 10 11 10 11 | 0.83 0.91 0.88 0.93 0.91 0.90 0.95 | 51 49 50 52 51 53 51 | 35 32 31 34 31 33 32 | 27 24 23 25 26 23 25 | 0.50 0.53 0.51 0.52 0.51 0.54 0.51 | 11 12 12 13 11 12 12 | 145 141 140 139 142 144 141 | 22 25 22 21 24 25 23 | 70 72 73 71 72 73 72 | 5.35 5.45 5.25 5.60 5.75 5.70 5.55 | 43 45 43 44 45 43 42 | 9.10 9.05 8.75 8.90 8.85 8.95 8.75 | 33 32 31 33 34 33 32 | |
| Heat | SH1 | 22 | 806 | 10 | 0.83 | 52 | 31 | 24 | 0.45 | 11 | 133 | 23 | 73 | 6.10 | 41 | 8.25 | 32 |
| SH2 | 21 | 817 | 11 | 0.80 | 53 | 32 | 23 | 0.48 | 12 | 136 | 22 | 70 | 5.95 | 40 | 8.10 | 30 | |
| SH3 | 20 | 823 | 12 | 0.81 | 50 | 30 | 25 | 0.43 | 13 | 135 | 24 | 71 | 5.85 | 42 | 8.35 | 31 | |
| SH4 SH5 SH6 SH7 SH8 SH9 SH10 | 23 21 22 23 21 20 23 | 826 817 826 825 829 827 821 | 9.0 9.0 10 11 12 11 10 | 0.84 0.81 0.83 0.80 0.85 0.82 0.84 | 56 53 51 53 55 52 51 | 33 30 31 32 33 30 32 | 26 23 22 25 25 22 24 | 0.45 0.47 0.42 0.44 0.46 0.45 0.47 | 11 13 12 11 12 13 12 | 134 137 136 133 135 131 133 | 21 24 23 22 23 24 25 | 70 71 72 70 71 72 71 | 6.15 6.25 6.10 5.85 5.95 6.10 6.15 | 41 40 41 39 40 40 42 | 8.00 8.15 8.27 8.20 8.34 8.31 8.26 | 32 31 30 32 33 32 31 | |
| Drought + Heat | SH1 | 18 | 606 | 7.3 | 0.50 | 45 | 40 | 19 | 0.25 | 7.5 | 173 | 14 | 62 | 3.25 | 39 | 5.25 | 27 |
| SH2 | 17 | 615 | 8.4 | 0.52 | 46 | 41 | 18 | 0.23 | 6.3. | 171 | 15 | 63 | 3.35 | 38 | 5.35 | 28 | |
| SH3 | 16 | 611 | 7.5 | 0.55 | 44 | 42 | 17 | 0.21 | 8.0 | 169 | 14 | 61 | 3.45 | 39 | 5.50 | 27 | |
| SH4 SH5 SH6 SH7 SH8 SH9 SH10 | 18 19 17 18 17 19 17 | 609 623 607 616 627 617 619 | 5.7 7.3 6.6 8.3 7.4 6.8 8.0 | 0.51 0.54 0.49 0.53 0.48 0.55 0.52 | 41 43 40 42 43 41 42 | 41 43 40 41 43 41 42 | 18 19 17 19 17 18 19 | 0.24 0.22 0.25 0.22 0.26 0.23 0.24 | 7.4 7.0 6.7 7.3 7.6 7.4 8.0 | 174 176 170 172 175 172 177 | 13 15 14 13 14 14 15 | 62 63 60 61 62 60 63 | 3.15 3.34 3.16 3.23 3.40 3.20 3.55 | 38 37 37 38 37 39 37 | 5.15 5.30 5.25 5.45 5.35 5.28 5.55 | 26 26 27 27 28 26 27 | |
| LSD | 2.00 | 6.00 | 0.75 | 0.03 | 2.00 | 1.85 | 1.03 | 0.005 | 0.3 | 5.00 | 1.2 | 2.0 | 0.06 | 1.00 | 0.30 | 1.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alghabari, F.; Shah, Z.H.; Seo, H. Physio-Chemical and Agronomic-Based Characterization of Synthetic Hexaploid Wheat Germplasm under Field Imposed Conditions of Drought and Heat Stress. Agronomy 2023, 13, 458. https://doi.org/10.3390/agronomy13020458
Alghabari F, Shah ZH, Seo H. Physio-Chemical and Agronomic-Based Characterization of Synthetic Hexaploid Wheat Germplasm under Field Imposed Conditions of Drought and Heat Stress. Agronomy. 2023; 13(2):458. https://doi.org/10.3390/agronomy13020458
Chicago/Turabian StyleAlghabari, Fahad, Zahid Hussain Shah, and Hyojin Seo. 2023. "Physio-Chemical and Agronomic-Based Characterization of Synthetic Hexaploid Wheat Germplasm under Field Imposed Conditions of Drought and Heat Stress" Agronomy 13, no. 2: 458. https://doi.org/10.3390/agronomy13020458
APA StyleAlghabari, F., Shah, Z. H., & Seo, H. (2023). Physio-Chemical and Agronomic-Based Characterization of Synthetic Hexaploid Wheat Germplasm under Field Imposed Conditions of Drought and Heat Stress. Agronomy, 13(2), 458. https://doi.org/10.3390/agronomy13020458