

Agroclimatic Requirements of Traditional European Pear (Pyrus communis L.) Cultivars from Australia, Europe, and North America

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Monitoring of Phenology
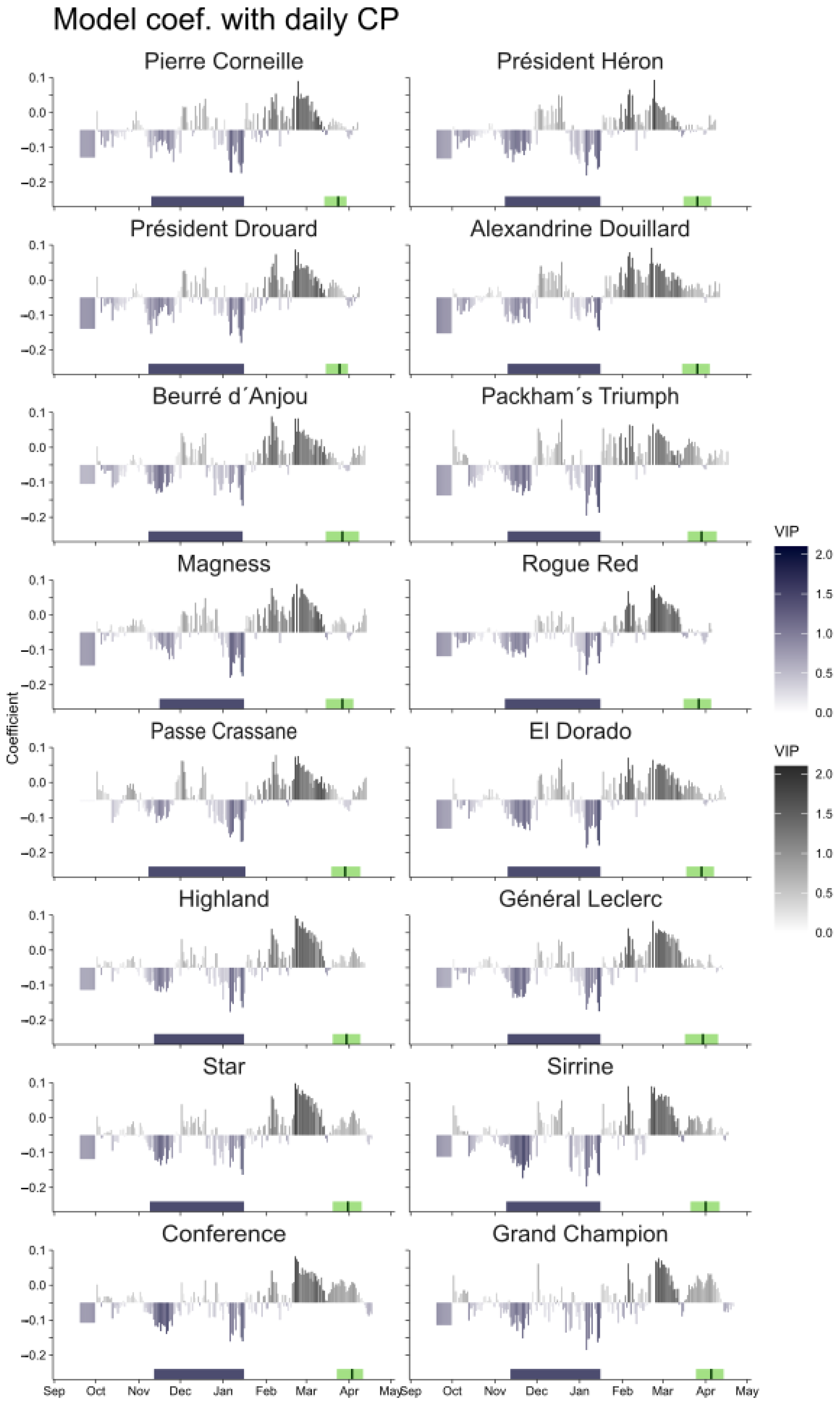
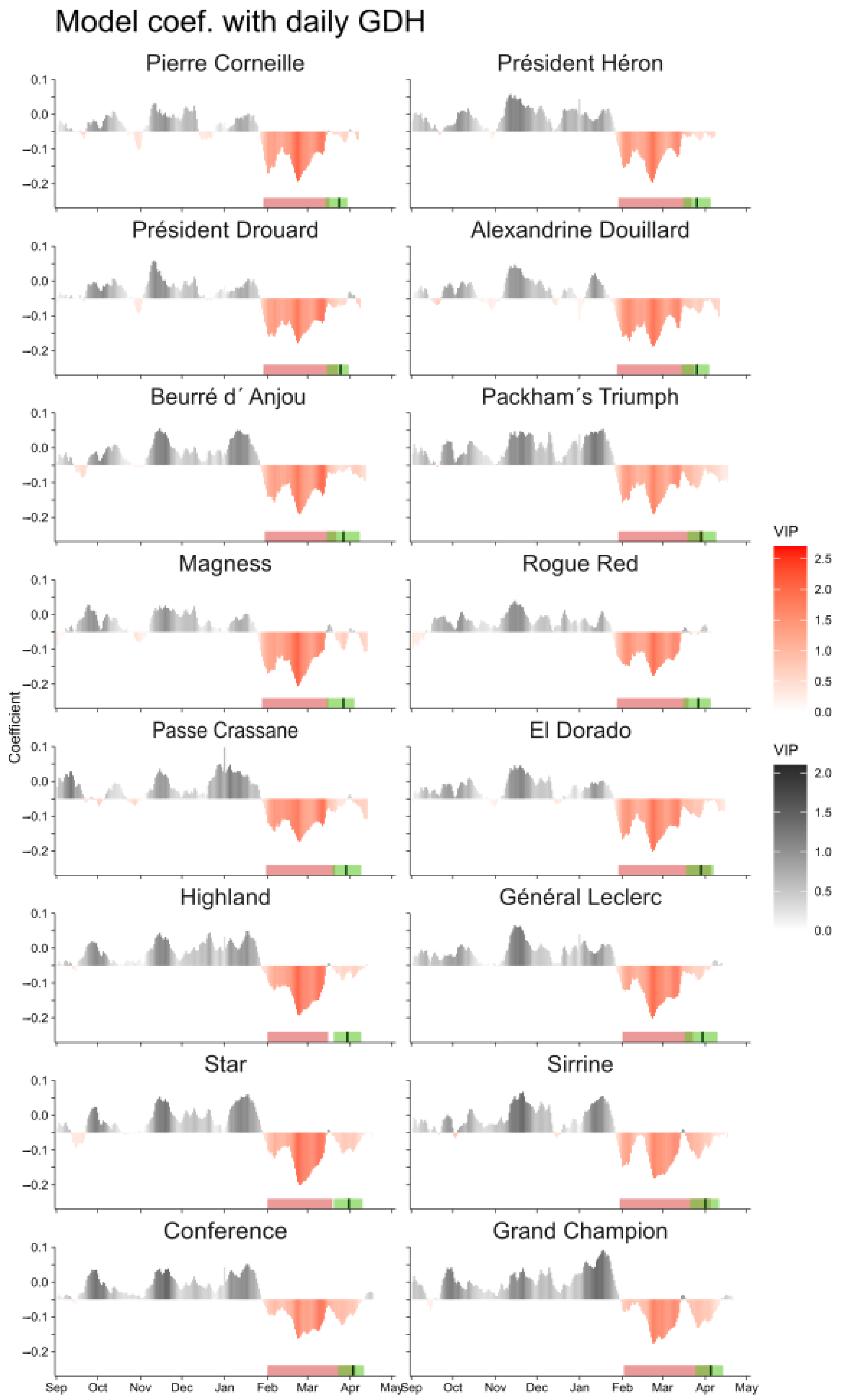
2.2. Delimitation of Chilling and Forcing Periods
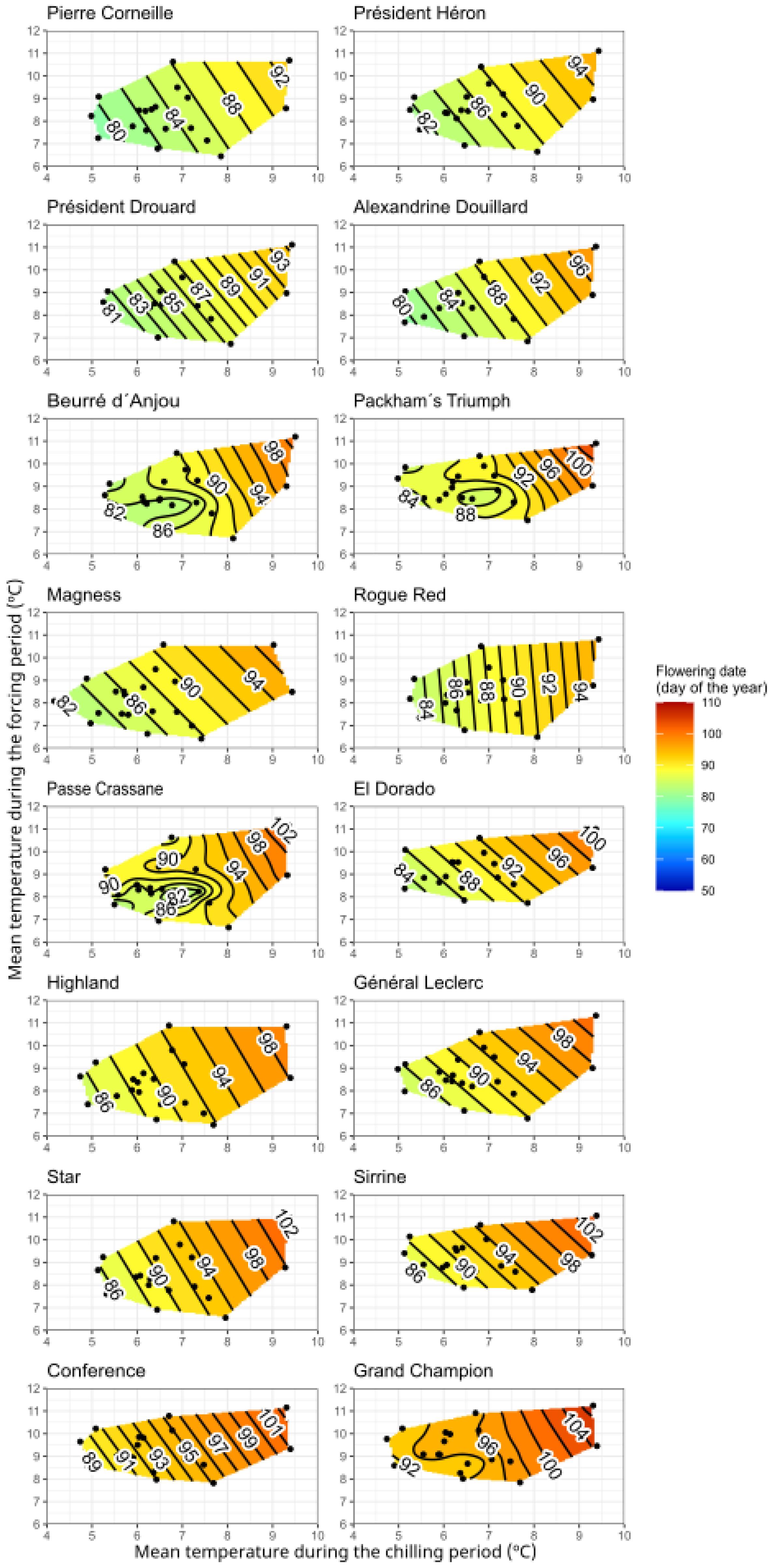
2.3. Determination of Chill and Heat Requirements
2.4. Chill and Heat Accumulation and Flowering Response
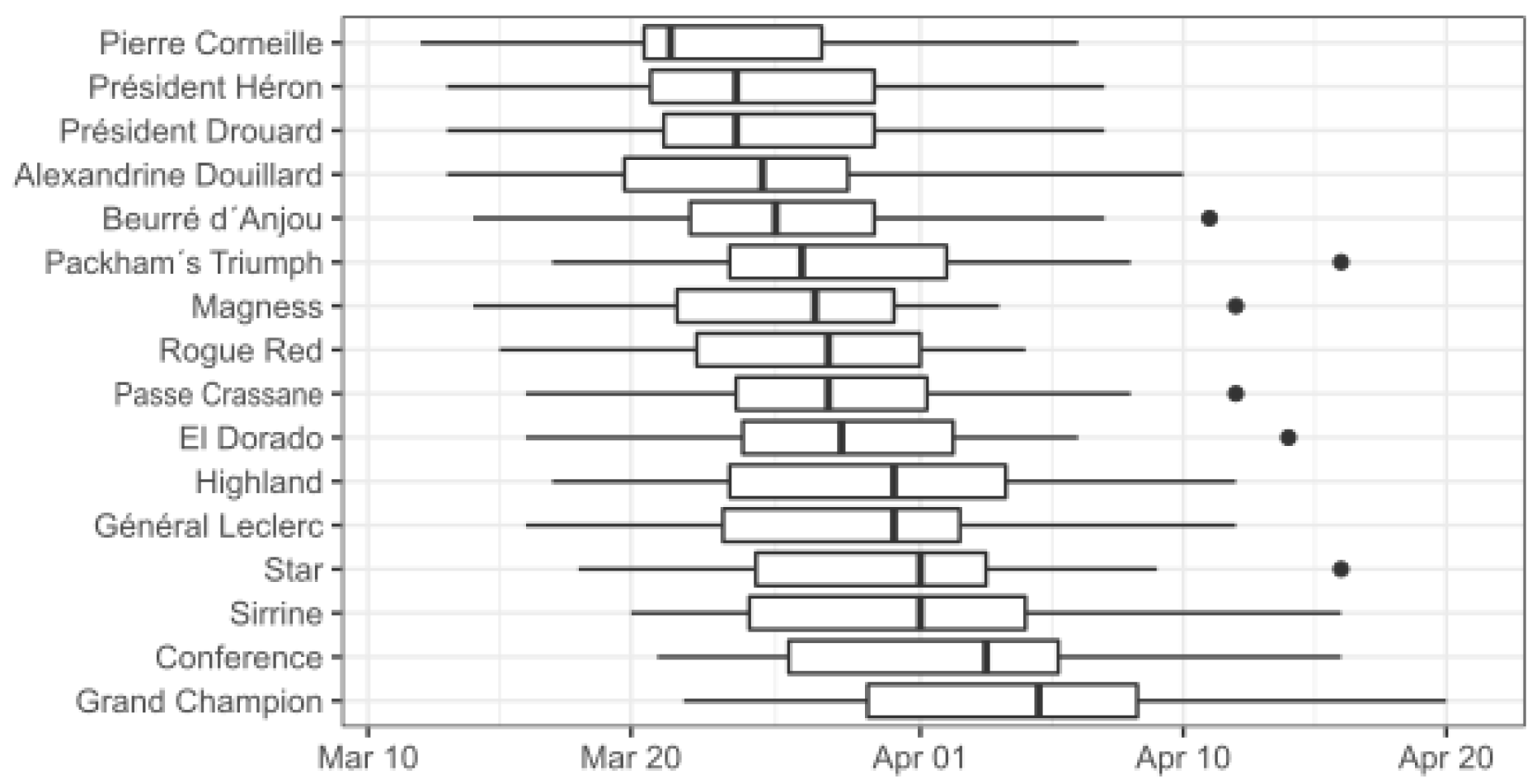
3. Results
3.1. Chilling Period Delineation
3.2. Forcing Period Delineation
3.3. Chill and Heat Requirements for Flowering
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Rohde, A.; Bhalerao, R.P. Plant dormancy in the perennial context. Trends Plant Sci. 2007, 12, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Campoy, J.A.; Ruiz, D.; Egea, J. Dormancy in temperate fruit trees in a global warming context: A review. Sci. Hortic. 2011, 130, 357–372. [Google Scholar] [CrossRef]
- Lang, G.A.; Early, J.D.; Martin, G.C.; Darnell, R.L. Endodormancy, paradormancy, and ecodormancy—Physiological terminology and classification for dormancy research. HortScience 1987, 22, 371–377. [Google Scholar] [CrossRef]
- Fadón, E.; Herrera, S.; Guerrero, B.I.; Guerra, M.E.; Rodrigo, J. Chilling and heat requirements of temperate stone fruit trees (Prunus sp.). Agronomy 2020, 10, 409. [Google Scholar] [CrossRef]
- Fadón, E.; Fernandez, E.; Behn, H.; Luedeling, E. A conceptual framework for winter dormancy in deciduous trees. Agronomy 2020, 10, 241. [Google Scholar] [CrossRef]
- Luedeling, E.; Zhang, M.; Girvetz, E.H. Climatic changes lead to declining winter chill for fruit and nut trees in California during 1950–2099. PLoS ONE 2009, 4, e6166. [Google Scholar] [CrossRef] [PubMed]
- Luedeling, E.; Girvetz, E.H.; Semenov, M.A.; Brown, P.H. Climate change affects winter chill for temperate fruit and nut trees. PLoS ONE 2011, 6, e20155. [Google Scholar] [CrossRef]
- Maulión, E.; Valentini, G.H.; Kovalevski, L.; Prunello, M.; Monti, L.L.; Daorden, M.E.; Quaglino, M.; Cervigni, G.D.L. Comparison of methods for estimation of chilling and heat requirements of nectarine and peach genotypes for flowering. Sci. Hortic. 2014, 177, 112–117. [Google Scholar] [CrossRef]
- Fernandez, E.; Mojahid, H.; Fadón, E.; Rodrigo, J.; Ruiz, D.; Egea, J.A.; Ben Mimoun, M.; Kodad, O.; El Yaacoubi, A.; Ghrab, M.; et al. Climate change impacts on winter chill in Mediterranean temperate fruit orchards. Reg. Environ. Chang. 2023, 23, 7. [Google Scholar] [CrossRef]
- Fernandez, E.; Whitney, C.; Luedeling, E. The importance of chill model selection—A multi-site analysis. Eur. J. Agron. 2020, 119, 126103. [Google Scholar] [CrossRef]
- Luedeling, E.; Brown, P.H. A global analysis of the comparability of winter chill models for fruit and nut trees. Int. J. Biometeorol. 2011, 55, 411–421. [Google Scholar] [CrossRef]
- Fadón, E.; Rodrigo, J. Unveiling winter dormancy through empirical experiments. Environ. Exp. Bot. 2018, 152, 28–36. [Google Scholar] [CrossRef]
- Brown, D.S.; Kotob, F.A. Growth of flower buds of apricot, peach, and pear during the rest period. Proc. Am. Soc. Hortic. Sci. 1957, 69, 158–164. [Google Scholar]
- El Yaacoubi, A.; Malagi, G.; Oukabli, A.; Citadin, I.; Hafidi, M.; Bonhomme, M.; Legave, J.M. Differentiated dynamics of bud dormancy and growth in temperate fruit trees relating to bud phenology adaptation, the case of apple and almond trees. Int. J. Biometeorol. 2016, 60, 1695–1710. [Google Scholar] [CrossRef]
- Malagi, G.; Sachet, M.R.; Citadin, I.; Herter, F.G.; Bonhomme, M.; Regnard, J.L.; Legave, J.M. The comparison of dormancy dynamics in apple trees grown under temperate and mild winter climates imposes a renewal of classical approaches. Trees Struct. Funct. 2015, 29, 1365–1380. [Google Scholar] [CrossRef]
- Luedeling, E.; Gassner, A. Partial Least Squares Regression for analyzing walnut phenology in California. Agric. For. Meteorol. 2012, 158–159, 43–52. [Google Scholar] [CrossRef]
- Luedeling, E.; Kunz, A.; Blanke, M.M. Identification of chilling and heat requirements of cherry trees—A statistical approach. Int. J. Biometeorol. 2013, 57, 679–689. [Google Scholar] [CrossRef]
- Fadón, E.; Rodrigo, J.; Luedeling, E. Cultivar-specific responses of sweet cherry flowering to rising temperatures during dormancy. Agric. For. Meteorol. 2021, 307, 108486. [Google Scholar] [CrossRef]
- Tabuenca, M.C. Necesidades de frio invernal de variedades de albaricoquero, melocotonero y peral. Ann. Aula Dei 1964, 7, 113–132. [Google Scholar]
- Tabuenca, M.C.; Herrero, J. Influencia de la temperatura en la época de floración de frutales. An. La Estac. Exp. Aula Dei 1966, 8, 115–153. [Google Scholar]
- Tabuenca, M.C. Winter chilling requirements of plum varieties. An. La Estac. Exp. Aula Dei 1967, 8, 383–391. [Google Scholar]
- Delgado, A.; Dapena, E.; Fernandez, E.; Luedeling, E. Climatic requirements during dormancy in apple trees from northwestern Spain—Global warming may threaten the cultivation of high-chill cultivars. Eur. J. Agron. 2021, 130, 126374. [Google Scholar] [CrossRef]
- Fernandez, E.; Luedeling, E.; Behrend, D.; Van de Vliet, S.; Fadón, E. Mild Water Stress Makes Apple Buds More Likely to Flower and More Responsive to Artificial Forcing—Impacts of an Unusually Warm and Dry Summer in Germany. Agronomy 2020, 10, 274. [Google Scholar] [CrossRef]
- Ferlito, F.; Di Guardo, M.; Allegra, M.; Nicolosi, E.; Continella, A.; La Malfa, S.; Gentile, A.; Distefano, G. Assessment of chilling requirement and threshold temperature of a low chill pear (Pyrus communis L.) germplasm in the mediterranean area. Horticulturae 2021, 7, 45. [Google Scholar] [CrossRef]
- Tamura, F.; Tanabe, K.; Itai, A.; Morimoto, M. Variations in the chilling requirements for breaking leaf bud endodormancy in wild pear species and pear cultivars. J. Jpn. Soc. Hortic. Sci. 2001, 70, 596–598. [Google Scholar] [CrossRef]
- Martínez-Lüscher, J.; Hadley, P.; Ordidge, M.; Xu, X.; Luedeling, E. Delayed chilling appears to counteract flowering advances of apricot in southern UK. Agric. For. Meteorol. 2017, 237–238, 209–218. [Google Scholar] [CrossRef]
- Benmoussa, H.; Ben Mimoun, M.; Ghrab, M.; Luedeling, E. Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location over 30 years of observation. Agric. For. Meteorol. 2017, 239, 34–46. [Google Scholar] [CrossRef]
- Fadón, E.; Rodrigo, J.; Luedeling, E. Temperature requirements for blooming in 12 sweet cherry cultivars. Acta Hortic. 2022, 1342, 103–110. [Google Scholar] [CrossRef]
- Delgado, A.; Egea, J.A.; Luedeling, E.; Dapena, E. Agroclimatic requirements and phenological responses to climate change of local apple cultivars in northwestern Spain. Sci. Hortic. 2021, 283, 110093. [Google Scholar] [CrossRef]
- Díez-Palet, I.; Funes, I.; Savé, R.; Biel, C.; Herralde, F.d.; Miarnau, X.; Vargas, F.; Àvila, G.; Carbó, J.; Aranda, X. Blooming under Mediterranean climate: Estimating cultivar-specific chill and heat requirements of almond and apple trees using a statistical approach. Agronomy 2019, 9, 760. [Google Scholar] [CrossRef]
- Watkins, R. Apple and Pear. In Evolution of Crop Plants; Simmonds, N.W., Ed.; Longman: London, UK; New York, NY, USA, 1976. [Google Scholar]
- Layne, R.E.C.; Qamme, H.A. Pears. In Advances in Fruit Breeding; Janick, J., Moore, J.N., Eds.; Purdue University Press: West Lafayette, IN, USA, 1975. [Google Scholar]
- Bell, R.L.; Quamme, H.A.; Layne, R.E.C.; Skirvin, R.M. Pears. In Fruit Breeding Vol. I. Tree and Tropical Fruits; Janik, J., More, J.N., Eds.; John Wiley & Sons, Inc.: New York, NY, USA, 1996. [Google Scholar]
- Fleckinger, J. Les stades végétatifs des arbres fruitiers, en rapport avec les traitements. Pomol. Fr. 1948, 1, 81–93. [Google Scholar]
- Meier, U. Growth Stages of Mono- and Dicotyledonous Plants: BBCH Monograph; Meier, U., Ed.; Federal Biological Research Centre for Agriculture and Forestry: Berlin/Braunschweig, Germany, 2001. [Google Scholar]
- Martínez-Nicolás, J.J.; Legua, P.; Melgarejo, P.; Martínez, R.; Hernández, F. Phenological growth stages of nashi tree (Pyrus pyrifolia): Codification and description according to the BBCH scale. Ann. Appl. Biol. 2015, 166, 136–142. [Google Scholar] [CrossRef]
- Brooks, R.M.; Olmo, H.P. The Brooks and Olmo Register of Fruit and Nut Varieties; American Society for Horticultural: Alexandria, VA, USA, 1997. [Google Scholar]
- Pyrus Genetic Resources of the USDA-ARS National Clonal Germplasm Repository (Corvallis, Oregon). Available online: https://www.ars.usda.gov/pacific-west-area/corvallis-or/national-clonal-germplasm-repository/docs/ncgr-corvallis-pyrus-germplasm/ (accessed on 5 December 2022).
- Oficina Del Regante. Available online: http://www.oficinaregante.aragon.es/ (accessed on 15 June 2022).
- Almorox, J.; Hontoria, C.; Benito, M. Statistical validation of daylength definitions for estimation of global solar radiation in Toledo, Spain. Energy Convers. Manag. 2005, 46, 1465–1471. [Google Scholar] [CrossRef]
- Linvill, D.E. Calculating Chilling Hours and Chill Units from Daily Maximum and Minimum Temperature Observations. HortScience 2019, 25, 14–16. [Google Scholar] [CrossRef]
- Luedeling, E.; Fernanez, E. chillR: Statistical Methods for Phenology Analysis in Temperate Fruit Trees, R Package Version 0.72.8; R Foundation for Statistical Computing: Vienna, Austria, 2022; URL http://www.R-project.org/.
- Weinberger, J.H. Chilling requirements of peach varieties. Proc. Am. Soc. Hortic. Sci. 1950, 56, 122–128. [Google Scholar]
- Richardson, E.A.; Seeley, S.D.; Walker, D.R. A model for estimating the completion of rest for “Redhaven” and “Elberta” peach trees. HortScience 1974, 9, 331–332. [Google Scholar] [CrossRef]
- Fishman, S.; Erez, A.; Couvillon, G.A. The temperature dependence of dormancy breaking in plants: Mathematical analysis of at two-step model involving a cooperative transition. J. Theor. Biol. 1987, 124, 473–483. [Google Scholar] [CrossRef]
- Anderson, J.L.; Richardson, E.A.; Kesner, C.D. Validation of Chill Unit and flower bud phenology models for “Montmorency”sour cherry. Acta Hortic. 1986, 184, 71–78. [Google Scholar] [CrossRef]
- Guo, L.; Dai, J.; Wang, M.; Xu, J.; Luedeling, E. Responses of spring phenology in temperate zone trees to climate warming: A case study of apricot flowering in China. Agric. For. Meteorol. 2015, 201, 1–7. [Google Scholar] [CrossRef]
- Wünsch, A.; Hormaza, J.I. Characterization of variability and genetic similarity of European pear using microsatellite loci developed in apple. Sci. Hortic. 2007, 113, 37–43. [Google Scholar] [CrossRef]
- Brini, W.; Mars, M.; Hormaza, J.I. Genetic diversity in local Tunisian pears (Pyrus communis L.) studied with SSR markers. Sci. Hortic. 2008, 115, 337–341. [Google Scholar] [CrossRef]
- Tabuenca, M.C. Winter chilling requirements of pear varieties. An. La Estac. Exp. Aula Deiles 1968, 9, 1–9. [Google Scholar]
- Spiegel-Roy, P.; Alston, F.H. Chilling and Post-Dormant Heat Requirement as Selection Criteria for Late-Flowering Pears. J. Hortic. Sci. 1979, 54, 115–120. [Google Scholar] [CrossRef]
- Fadón, E.; Fernandez, E.; Do, H.T.; Kunz, A.; Krefting, P.; Luedeling, E. Chill and heat accumulation modulates phenology in temperate fruit trees. Acta Hortic. 2021, 1327, 413–420. [Google Scholar] [CrossRef]
- Fernandez, E.; Krefting, P.; Kunz, A.; Do, H.; Fadón, E.; Luedeling, E. Boosting statistical delineation of chill and heat periods in temperate fruit trees through multi-environment observations. Agric. For. Meteorol. 2021, 310, 108652. [Google Scholar] [CrossRef]
- Fadón, E.; Do, H.; Blanke, M.; Rodrigo, J.; Luedeling, E. Apparent Differences in Agroclimatic Requirements for Sweet Cherry Across Climatic Settings Reveal Shortcomings in Common Phenology Models. SSRN Electron. J. 2022. [Google Scholar] [CrossRef]
- Jiménez, S.; Reighard, G.L.; Bielenberg, D.G. Gene expression of DAM5 and DAM6 is suppressed by chilling temperatures and inversely correlated with bud break rate. Plant Mol. Biol. 2010, 73, 157–167. [Google Scholar] [CrossRef]
- Quesada-Traver, C.; Guerrero, B.I.; Badenes, M.L.; Rodrigo, J.; Ríos, G.; Lloret, A. Structure and Expression of Bud Dormancy-Associated MADS-Box Genes (DAM) in European Plum. Front. Plant Sci. 2020, 11, 1288. [Google Scholar] [CrossRef]
- Leida, C.; Romeu, J.F.; García-Brunton, J.; Ríos, G.; Badenes, M.L. Gene expression analysis of chilling requirements for flower bud break in peach. Plant Breed. 2012, 131, 329–334. [Google Scholar] [CrossRef]
- Fadón, E.; Herrero, M.; Rodrigo, J. Dormant flower buds actively accumulate starch over winter in sweet cherry. Front. Plant Sci. 2018, 9, 171. [Google Scholar] [CrossRef]
- Herrera, S.; Lora, J.; Fadón, E.; Alonso, J.M.; Hormaza, J.I.; Rodrigo, J. Male meiosis as a biomarker of the transition from endodormancy to ecodormancy in apricot. Front. Plant Sci. 2022, 13, 842333. [Google Scholar] [CrossRef] [PubMed]
- Julian, C.; Herrero, M.; Rodrigo, J. Anther meiosis time is related to winter cold temperatures in apricot (Prunus armeniaca L.). Environ. Exp. Bot. 2014, 100, 20–25. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | Cultivar | Release Year | Breeder/Breeding Program |
|---|---|---|---|
| Australia | Packham’s Triumph | 1897 | Sam Packham (New South Wales) |
| France | Alexandrine Douillard | 1849 | Constant Douillard (Nantes) |
| Beurré d´Anjou | 1819 | ||
| Général Leclerc | <1950 | Fruit Research Station of Angers | |
| Passe Crassane | 1845 | Louis Boisbunel (Rouen, Normardie) | |
| Président Drouard | 1876 | ||
| Président Héron | 1894 | M. Arsene Sannier (Rouen, Normardie) | |
| Pierre Corneille | 1894 | ||
| United Kingdom | Conference | 1885 | Thomas River (Rivers Nursery, Sawbridgeworth) |
| USA | Grand Champion | 1936 | |
| El Dorado | 1925 | Seedling (Placerville, California) | |
| Highland | 1974 | Robert C. Lamb (New York State Agriculture Experiment Station, Geneva) | |
| Magness | 1960 | Howard J. Brooks (United States Department of Agriculture, Beltsville, Maryland) | |
| Rogue Red | 1969 | F.C. Reimer, E. Degman, and V. Quackenbush (Oregon State University) | |
| Sirrine | 1954 | F. Atwood Sirrine | |
| Star | 1968 |
| Cultivar | Chilling | Forcing | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Period | Accumulation (Mean ± sd) | Period | Accumulation (Mean ± sd) | |||||||
| Start | End | Duration (Days) | CP | CU | CH | Start | End (Flowering Date, Average) | Duration (Days) | GDH | |
| Alexandrine Douillard | 10 Nov | 16 Jan | 67 | 47.5 ± 2.5 | 1027 ± 141 | 753 ± 117 | 28 Jan | 26 Mar | 58 | 7287 ± 945 |
| Beurré d´Anjou | 08 Nov | 15 Jan | 65 | 47.7 ± 2.8 | 1100 ± 100 | 745 ± 117 | 30 Jan | 03 Apr | 62 | 6619 ± 750 |
| Conference | 12 Nov | 16 Jan | 67 | 46.5 ± 2.3 | 1113 ± 186 | 745 ± 114 | 1 Feb | 29 Mar | 60 | 7347 ± 905 |
| El Dorado | 10 Nov | 16 Jan | 67 | 47.5 ± 2.5 | 1163 ± 30 | 753 ± 117 | 29 Jan | 30 Mar | 58 | 7094 ± 1082 |
| Général Leclerc | 10 Nov | 16 Jan | 65 | 47.5 ± 2.5 | 1028 ± 140 | 753 ± 117 | 1 Feb | 05 Apr | 63 | 7107 ± 890 |
| Grand Champion | 12 Nov | 16 Jan | 65 | 46.5 ± 2.3 | 1028 ± 140 | 745 ± 114 | 2 Feb | 30 Mar | 58 | 7168 ± 776 |
| Highland | 12 Nov | 16 Jan | 61 | 46.5 ± 2.3 | 1113 ± 186 | 745 ± 114 | 1 Feb | 27 Mar | 59 | 7402 ± 853 |
| Magness | 16 Nov | 16 Jan | 68 | 43.9 ± 2.0 | 1113 ± 186 | 719 ± 106 | 28 Jan | 27 Mar | 57 | 7509 ± 1048 |
| Packham´s Triumph | 10 Nov | 16 Jan | 67 | 47.5 ± 2.5 | 1163 ± 31 | 753 ± 117 | 29 Jan | 29 Mar | 60 | 6997 ± 1048 |
| Passe Crassane | 08 Nov | 17 Jan | 70 | 49.2 ± 2.8 | 1100 ± 100 | 774 ± 120 | 31 Jan | 29 Mar | 58 | 6514 ± 673 |
| Pierre Corneille | 10 Nov | 16 Jan | 67 | 47.5 ± 2.5 | 1099 ± 99 | 753 ± 117 | 29 Jan | 24 Mar | 55 | 6519 ± 741 |
| Président Drouard | 08 Nov | 16 Jan | 69 | 48.5 ± 2.8 | 1113 ± 186 | 760 ± 118 | 29 Jan | 25 Mar | 56 | 7289 ± 914 |
| Président Heron | 08 Nov | 16 Jan | 69 | 48.5 ± 2.8 | 1163 ± 30 | 760 ± 118 | 29 Jan | 26 Mar | 57 | 6935 ± 1042 |
| Rogue Red | 08 Nov | 16 Jan | 69 | 48.5 ± 2.8 | 1028 ± 140 | 760 ± 118 | 28 Jan | 27 Mar | 59 | 7170 ± 656 |
| Sirrine | 09 Nov | 16 Jan | 68 | 48.0 ± 2.8 | 1163 ± 30 | 757 ± 118 | 30 Jan | 01 Apr | 62 | 7075 ± 902 |
| Star | 09 Nov | 16 Jan | 68 | 48.0 ± 2.8 | 1100 ± 100 | 757 ± 118 | 1 Feb | 31 Mar | 59 | 6573 ± 637 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fadón, E.; Espiau, M.T.; Errea, P.; Alonso Segura, J.M.; Rodrigo, J. Agroclimatic Requirements of Traditional European Pear (Pyrus communis L.) Cultivars from Australia, Europe, and North America. Agronomy 2023, 13, 518. https://doi.org/10.3390/agronomy13020518
Fadón E, Espiau MT, Errea P, Alonso Segura JM, Rodrigo J. Agroclimatic Requirements of Traditional European Pear (Pyrus communis L.) Cultivars from Australia, Europe, and North America. Agronomy. 2023; 13(2):518. https://doi.org/10.3390/agronomy13020518
Chicago/Turabian StyleFadón, Erica, María Teresa Espiau, Pilar Errea, José Manuel Alonso Segura, and Javier Rodrigo. 2023. "Agroclimatic Requirements of Traditional European Pear (Pyrus communis L.) Cultivars from Australia, Europe, and North America" Agronomy 13, no. 2: 518. https://doi.org/10.3390/agronomy13020518
APA StyleFadón, E., Espiau, M. T., Errea, P., Alonso Segura, J. M., & Rodrigo, J. (2023). Agroclimatic Requirements of Traditional European Pear (Pyrus communis L.) Cultivars from Australia, Europe, and North America. Agronomy, 13(2), 518. https://doi.org/10.3390/agronomy13020518