
Plant Growth Promoting Rhizobacteria (PGPR) and Arbuscular Mycorrhizal Fungi Combined Application Reveals Enhanced Soil Fertility and Rice Production

 ,
, 
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation and Application of AMF and PGPR Inoculum
2.2. Soil Characterization
2.3. Soil Nitrogen (N), Extractable Phosphorus (P), Potassium (K), and Boron (B)
2.4. Experimental Design of Pot Trial
2.5. Plant Leaf Analysis
2.6. Agronomic Parameters
2.7. Data Statistical Analysis, Pearson Correlation, and PCA
3. Results
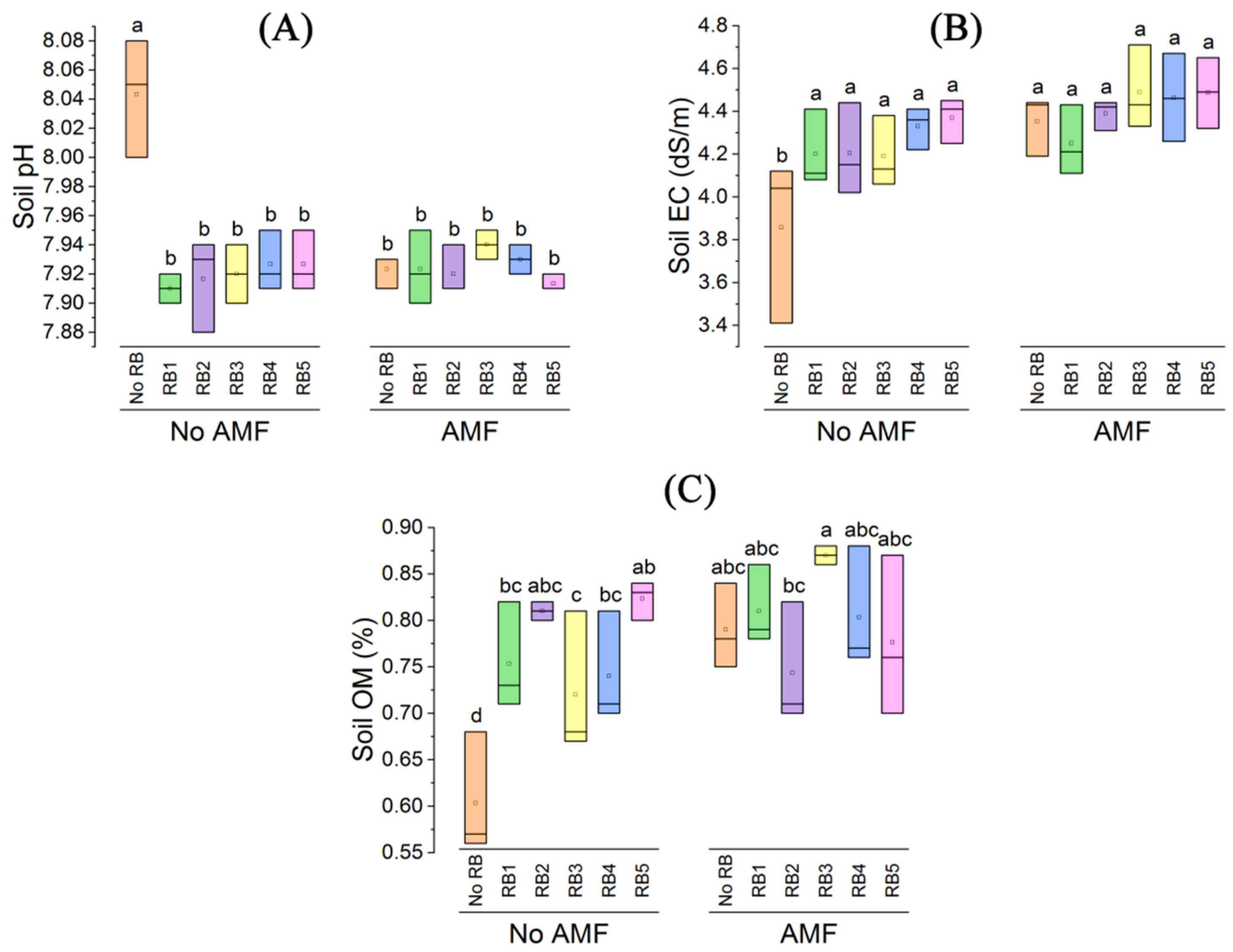
3.1. Soil pH, EC, and Organic Matter
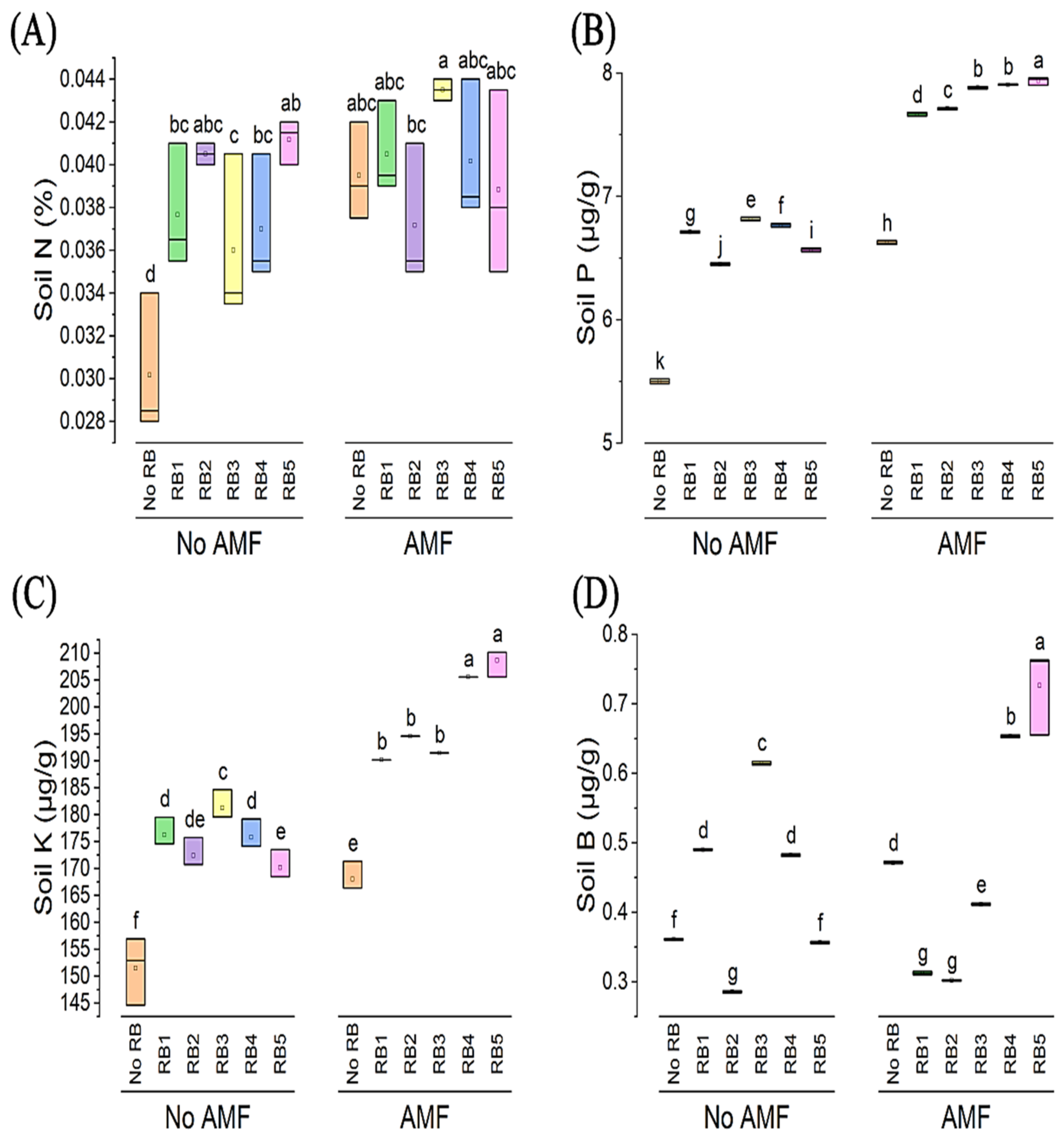
3.2. Soil Analysis
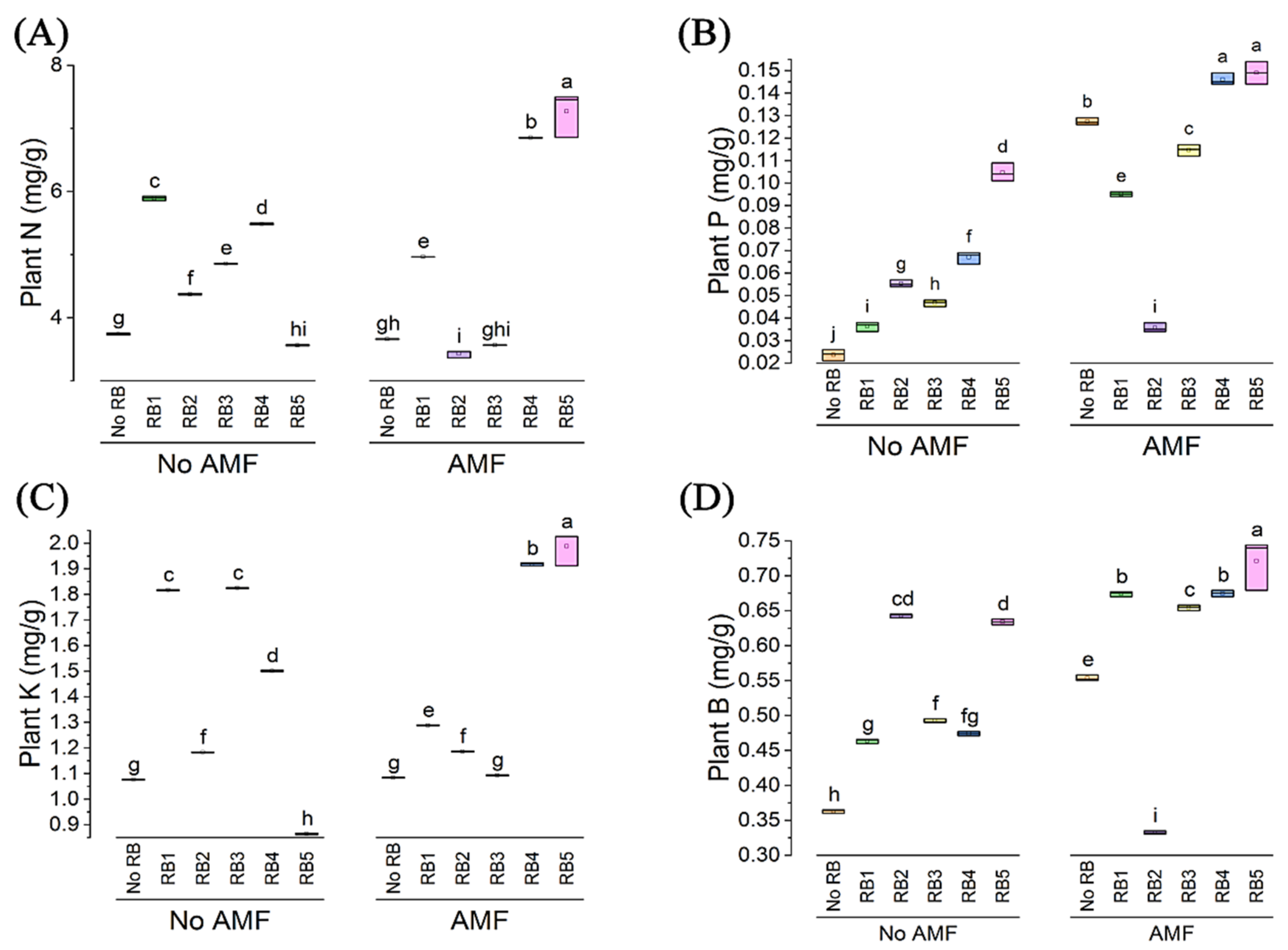
3.3. Plant Leaf Analysis
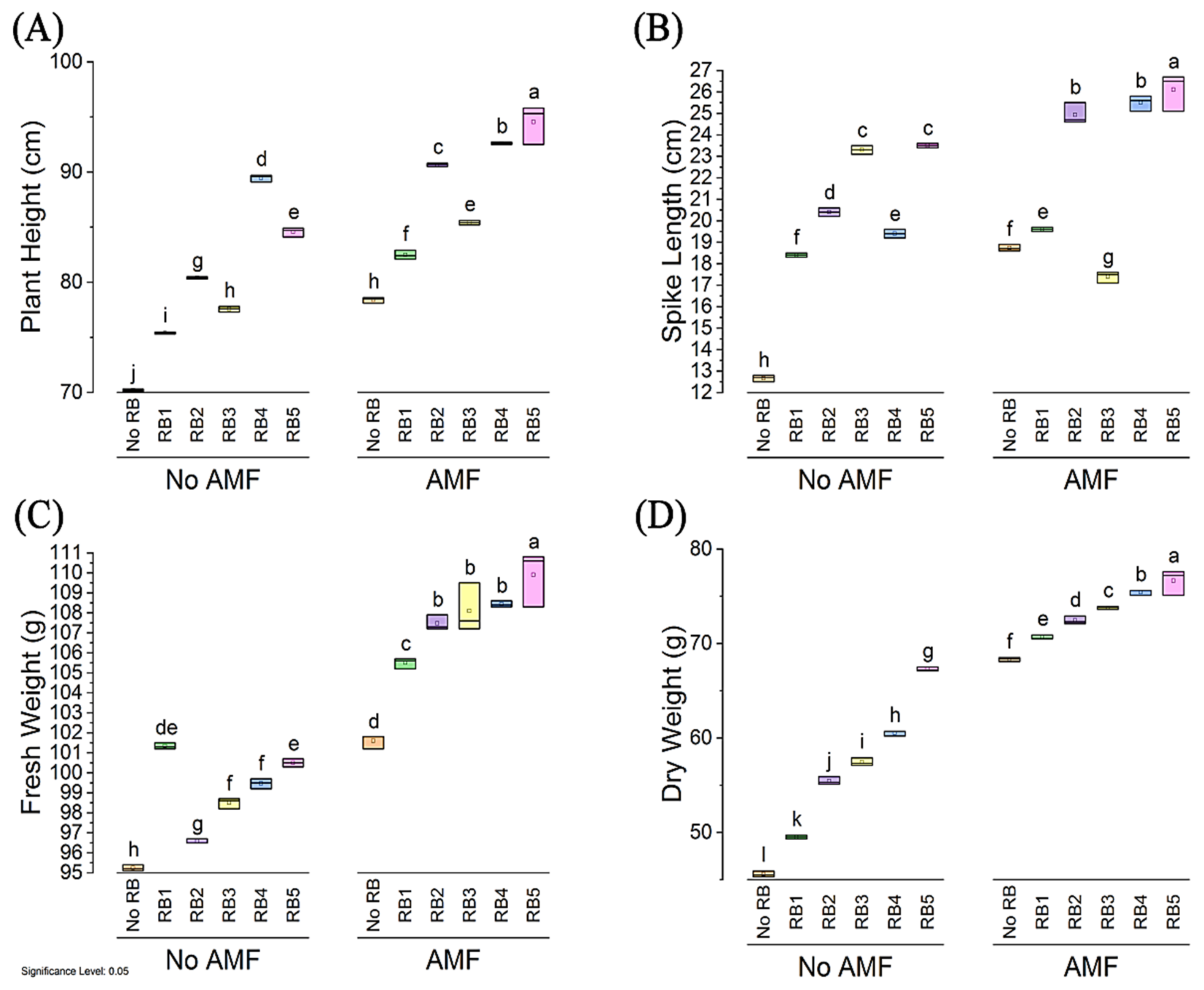
3.4. Morpho-Biochemical Characteristics
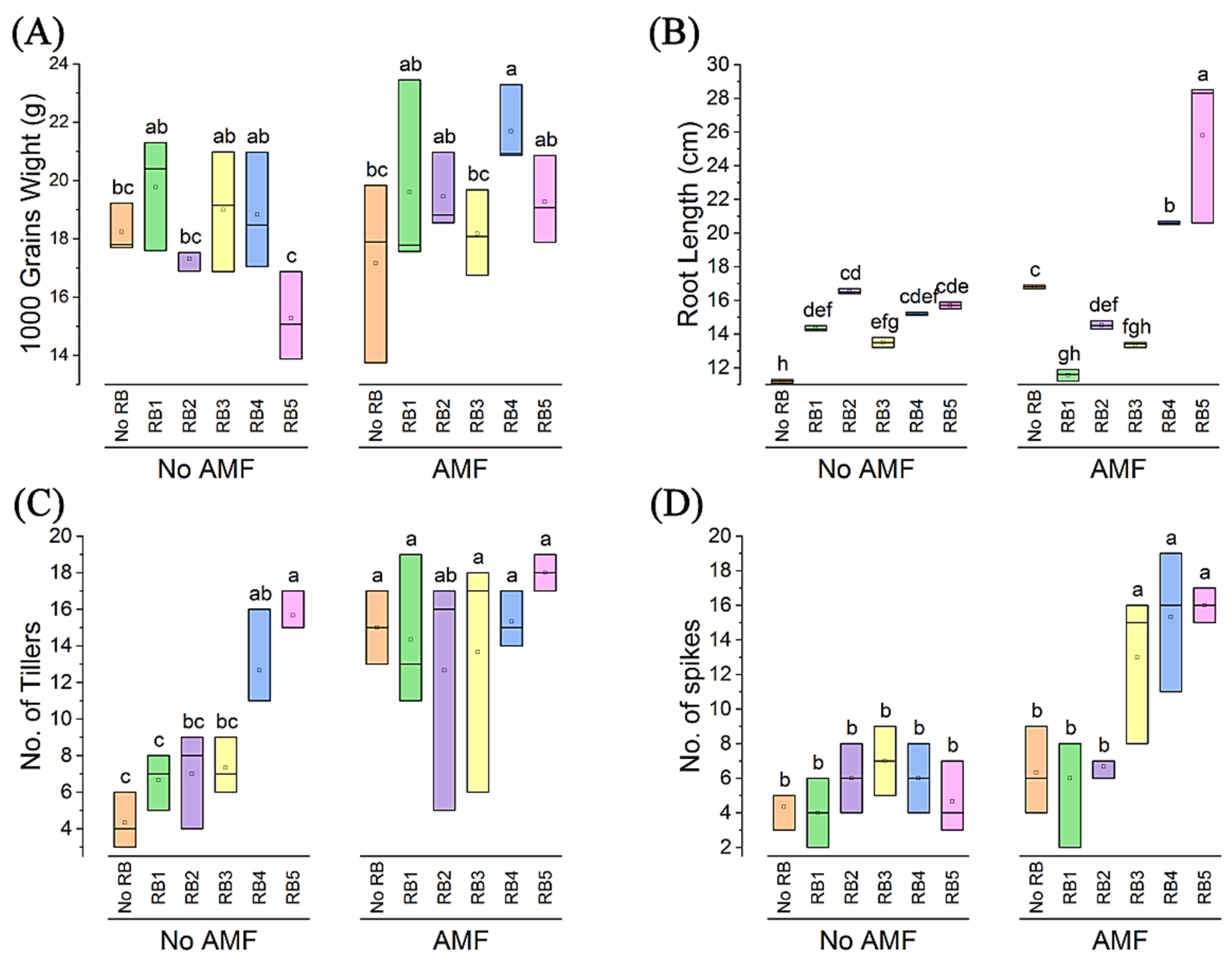
3.5. Root Length, 1000-Grain Weight, No. of Tillers and No. of Spikes
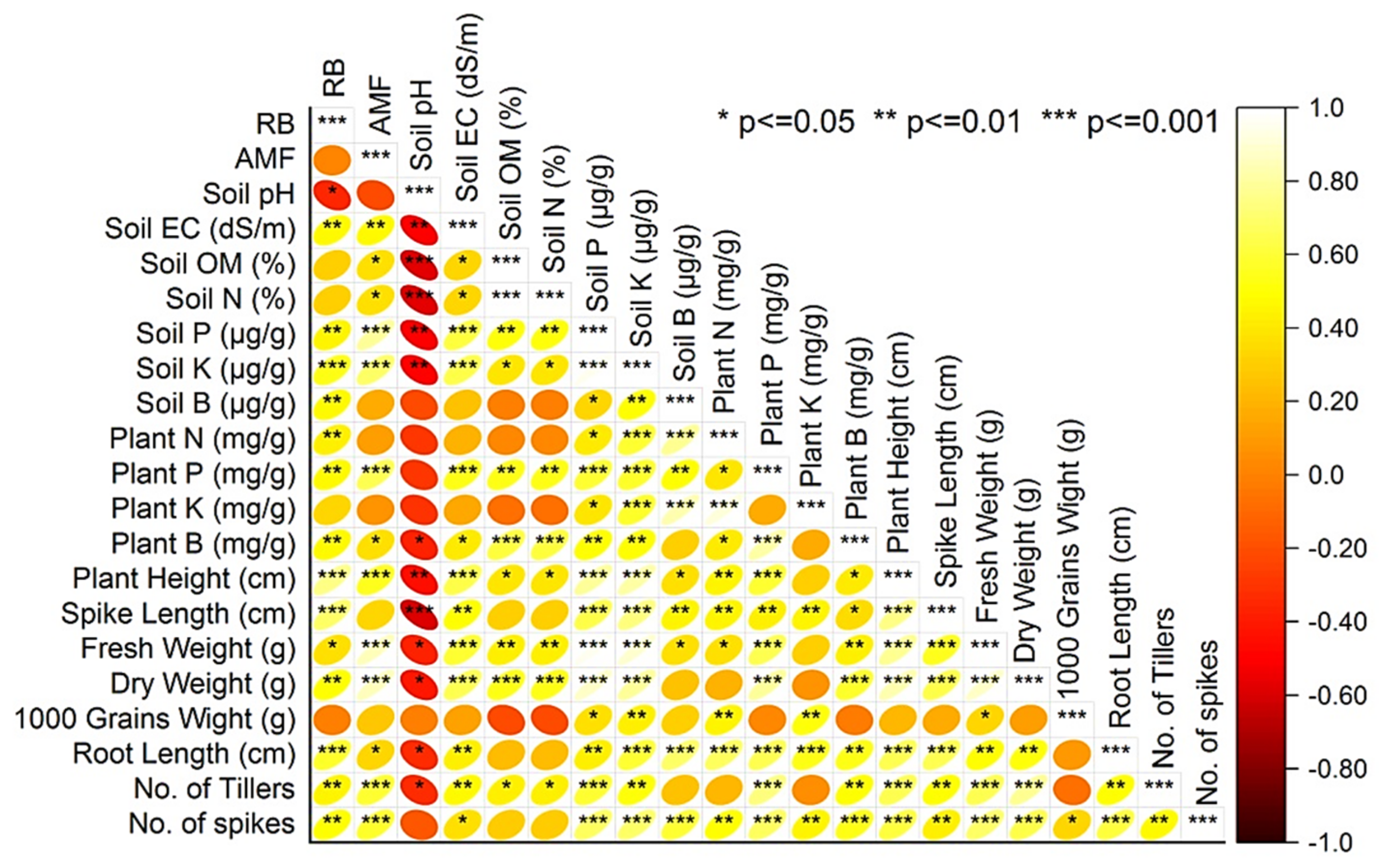
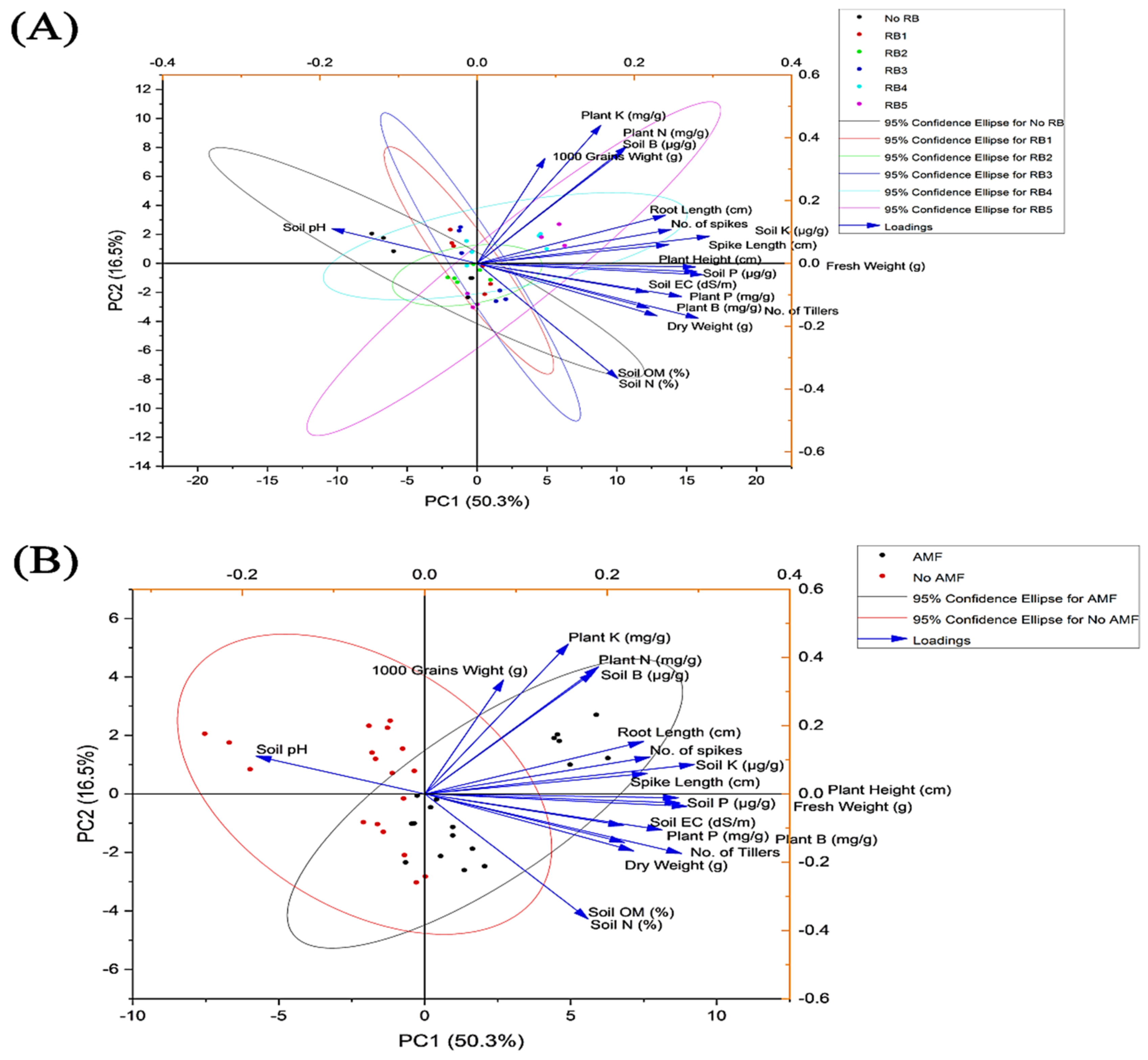
3.6. Pearson’s Correlation among the Variables and Principal Component Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ejaz, W. Agriculture Pak. Economic Survey Report Finance Division Government of Pakistan, Islamabad. 2018. Available online: https://www.finance.gov.pk/survey/chapters_19/2-Agriculture.pdf (accessed on 9 February 2023).
- Wasim, M.P. A Study of Rice in the Major Growing Countries of the World: Their Growth Instability and World Share. Pak. Econ. Soc. Rev. 2002, 40, 153–183. [Google Scholar]
- World Health Organization; Food and Agriculture Organization of the United Nations. Preparation and Use of Food-Based Dietary Guidelines; Report of a Joint FAO/WHO Consultation; WHO: Geneva, Switzerland, 1996.
- Encyclopaedia Britannica. Britannica Book of the Year 2010; Encyclopaedia Britannica, Inc.: Edinburgh, UK, 2010; ISBN 1615353666. [Google Scholar]
- Gangwar, R.K.; Javeria, S.; Yadav, K.; Tyagi, S.; Singh, R. Survey and Surveillance of Major Insect-Pests of Basmati Rice in Western Uttar Pradesh (India). Int. J. Res. Appl. Nat. Soc. Sci. 2015, 3, 1–8. [Google Scholar]
- Santoyo, G.; Urtis-Flores, C.A.; Loeza-Lara, P.D.; Orozco-Mosqueda, M.; Glick, B.R. Rhizosphere Colonization Determinants by Plant Growth-Promoting Rhizobacteria (PGPR). Biology 2021, 10, 475. [Google Scholar] [CrossRef] [PubMed]
- Gusain, P.; Bhandari, B.S. Rhizosphere Associated PGPR Functioning. J. Pharmacogn. Phytochem. 2019, 8, 1181–1191. [Google Scholar]
- Delaporte-Quintana, P.; Lovaisa, N.C.; Rapisarda, V.A.; Pedraza, R.O. The Plant Growth Promoting Bacteria GluconacetobacterDiazotrophicus and AzospirillumBrasilense Contribute to the Iron Nutrition of Strawberry Plants through Siderophores Production. Plant Growth Regul. 2020, 91, 185–199. [Google Scholar] [CrossRef]
- Patel, P.; Trivedi, G.; Saraf, M. Iron Biofortification in Mungbean Using Siderophore Producing Plant Growth Promoting Bacteria. Environ. Sustain. 2018, 1, 357–365. [Google Scholar] [CrossRef]
- Uzoh, I.M.; Babalola, O.O. Review on Increasing Iron Availability in Soil and Its Content in Cowpea (Vigna Unguiculata) by Plant Growth Promoting Rhizobacteria. Afr. J. Food Agric. Nutr. Dev. 2020, 20, 15779–15799. [Google Scholar] [CrossRef]
- Billah, M.; Khan, M.; Bano, A.; Hassan, T.U.; Munir, A.; Gurmani, A.R. Phosphorus and Phosphate Solubilizing Bacteria: Keys for Sustainable Agriculture. Geomicrobiol. J. 2019, 36, 904–916. [Google Scholar] [CrossRef]
- Reyes-Castillo, A.; Gerding, M.; Oyarzúa, P.; Zagal, E.; Gerding, J.; Fischer, S. Plant Growth-Promoting Rhizobacteria Able to Improve NPK Availability: Selection, Identification and Effects on Tomato Growth. Chil. J. Agric. Res. 2019, 79, 473–485. [Google Scholar] [CrossRef]
- Sabir, A.; Naveed, M.; Bashir, M.A.; Hussain, A.; Mustafa, A.; Zahir, Z.A.; Kamran, M.; Ditta, A.; Núñez-Delgado, A.; Saeed, Q.; et al. Cadmium Mediated Phytotoxic Impacts in Brassica Napus: Managing Growth, Physiological and Oxidative Disturbances through Combined Use of Biochar and Enterobacter Sp. MN17. J. Environ. Manag. 2020, 265, 110522. [Google Scholar] [CrossRef]
- Coniglio, A.; Mora, V.; Puente, M.; Cassán, F. Azospirillum as Biofertilizer for Sustainable Agriculture: AzospirillumBrasilense AZ39 as a Model of PGPR and Field Traceability. In Microbial Probiotics for Agricultural Systems; Springer: Berlin/Heidelberg, Germany, 2019; pp. 45–70. [Google Scholar]
- Cassán, F.; Coniglio, A.; López, G.; Molina, R.; Nievas, S.; de Carlan, C.L.N.; Donadio, F.; Torres, D.; Rosas, S.; Pedrosa, F.O. Everything You Must Know about Azospirillum and Its Impact on Agriculture and Beyond. Biol. Fertil. Soils 2020, 56, 461–479. [Google Scholar] [CrossRef]
- Riaz, M.; Kamran, M.; Fang, Y.; Wang, Q.; Cao, H.; Yang, G.; Deng, L.; Wang, Y.; Zhou, Y.; Anastopoulos, I.; et al. Arbuscular Mycorrhizal Fungi-Induced Mitigation of Heavy Metal Phytotoxicity in Metal Contaminated Soils: A Critical Review. J. Hazard. Mater. 2021, 402, 123919. [Google Scholar] [CrossRef] [PubMed]
- Savolainen, T.; Kytöviita, M.-M. Mycorrhizal Symbiosis Changes Host Nitrogen Source Use. Plant Soil 2021, 471, 643–654. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic Press: Cambridge, MA, USA, 2010; ISBN 0080559344. [Google Scholar]
- Jakobsen, I.; Rosendahl, L. N2 Fixation and Root Respiration in Pea: Effects of VAM and P. Agric. Ecosyst. Environment. 1990, 29, 205–209. [Google Scholar] [CrossRef]
- Schwember, A.R.; Schulze, J.; Del Pozo, A.; Cabeza, R.A. Regulation of Symbiotic Nitrogen Fixation in Legume Root Nodules. Plants 2019, 8, 333. [Google Scholar] [CrossRef]
- Londoño, D.M.M.; Meyer, E.; da Silva, K.J.; Hernández, A.G.; de Armas, R.D.; Soares, L.M.; Stürmer, S.L.; Nodari, R.O.; Soares, C.R.F.S.; Lovato, P.E. Root Colonization and Arbuscular Mycorrhizal Fungal Community Composition in a Genetically Modified Maize, Its Non-Modified Isoline, and a Landrace. Mycorrhiza 2020, 30, 611–621. [Google Scholar] [CrossRef]
- Saxena, D.; Stewart, C.N.; Altosaar, I.; Shu, Q.; Stotzky, G. Larvicidal Cry Proteins from Bacillus Thuringiensis Are Released in Root Exudates of Transgenic B. Thuringiensis Corn, Potato, and Rice but Not of B. Thuringiensis Canola, Cotton, and Tobacco. Plant Physiol. Biochem. 2004, 42, 383–387. [Google Scholar] [CrossRef]
- Staddon, P.L.; Ramsey, C.B.; Ostle, N.; Ineson, P.; Fitter, A.H. Rapid Turnover of Hyphae of Mycorrhizal Fungi Determined by AMS Microanalysis of 14C. Science 2003, 300, 1138–1140. [Google Scholar] [CrossRef]
- Okonji, C.J.; Sakariyawo, O.S.; Okeleye, K.A.; Osunbiyi, A.G.; Ajayi, E.O. Effects of Arbuscular Mycorrhizal Fungal Inoculation on Soil Properties and Yield of Selected Rice Varieties. J. Agric. Sci. 2018, 63, 153–170. [Google Scholar] [CrossRef]
- Cera, A.; Duplat, E.; Montserrat-Martí, G.; Gómez-Bolea, A.; Rodríguez-Echeverría, S.; Palacio, S. Seasonal Variation in AMF Colonisation, Soil and Plant Nutrient Content in Gypsum Specialist and Generalist Species Growing in P-Poor Soils. Plant Soil 2021, 468, 509–524. [Google Scholar] [CrossRef]
- de Vries, J.; Evers, J.B.; Kuyper, T.W.; van Ruijven, J.; Mommer, L. Mycorrhizal Associations Change Root Functionality: A 3D Modelling Study on Competitive Interactions between Plants for Light and Nutrients. New Phytol. 2021, 231, 1171. [Google Scholar] [CrossRef] [PubMed]
- Newman, E.I. Mycorrhizal Links between Plants: Their Functioning and Ecological Significance. Adv. Ecol. Res. 1988, 18, 243–270. [Google Scholar]
- Hung, L.-L.L.; Sylvia, D.M. Production of Vesicular-Arbuscular Mycorrhizal Fungus Inoculum in Aeroponic Culture. Appl. Environ. Microbiol. 1988, 54, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Chahal, K.; Gupta, V.; Verma, N.K.; Chaurasia, A.; Rana, B. Arbuscular Mycorrhizal (AM) Fungi as a Tool for Sustainable Agricultural System. Mycorrhizal Fungi Util. Agric. For. 2021, 19, 1–12. [Google Scholar] [CrossRef]
- Salomon, M.J. Soil Health in Urban Agriculture with an Emphasis on Arbuscular Mycorrhizal Fungi. Ph.D. Thesis, The University of Adelaide Library, Adelaide, Australia, 2021. [Google Scholar]
- Bhantana, P.; Rana, M.S.; Sun, X.; Moussa, M.G.; Saleem, M.H.; Syaifudin, M.; Shah, A.; Poudel, A.; Pun, A.B.; Bhat, M.A.; et al. Arbuscular Mycorrhizal Fungi and Its Major Role in Plant Growth, Zinc Nutrition, Phosphorous Regulation and Phytoremediation. Symbiosis 2021, 84, 19–37. [Google Scholar] [CrossRef]
- Vafadar, F.; Amooaghaie, R.; Otroshy, M. Effects of Plant-Growth-Promoting Rhizobacteria and Arbuscular Mycorrhizal Fungus on Plant Growth, Stevioside, NPK, and Chlorophyll Content of Stevia Rebaudiana. J. Plant Interact. 2014, 9, 128–136. [Google Scholar] [CrossRef]
- Jiang, F.; Zhang, L.; Zhou, J.; George, T.S.; Feng, G. Arbuscular Mycorrhizal Fungi Enhance Mineralisation of Organic Phosphorus by Carrying Bacteria along Their Extraradical Hyphae. New Phytol. 2021, 230, 304–315. [Google Scholar] [CrossRef]
- Azizi, S.; Tabari Kouchaksaraei, M.; Hadian, J.; FallahNosrat Abad, A.R.; Modarres Sanavi, S.A.M.; Ammer, C.; Bader, M.K.-F. Dual Inoculations of Arbuscular Mycorrhizal Fungi and Plant Growth-Promoting Rhizobacteria Boost Drought Resistance and Essential Oil Yield of Common Myrtle. For. Ecol. Manag. 2021, 497, 119478. [Google Scholar] [CrossRef]
- Charles, P.; Raj, A.D.S.; Kiruba, S. Arbuscular Mycorrhizal Fungi in the Reclamation and Restoration of Soil Fertility. Mycorrhiza News 2006, 18, 13–14. [Google Scholar]
- Parihar, M.; Rakshit, A.; Meena, V.S.; Gupta, V.K.; Rana, K.; Choudhary, M.; Tiwari, G.; Mishra, P.K.; Pattanayak, A.; Bisht, J.K. The Potential of Arbuscular Mycorrhizal Fungi in C Cycling: A Review. Arch. Microbiol. 2020, 202, 1581–1596. [Google Scholar] [CrossRef]
- Sharpley, A.; Daniel, T.C.; Sims, J.T.; Pote, D.H. Determining Environmentally Sound Soil Phosphorus Levels. J. Soil Water Conserv. 1996, 51, 160–166. [Google Scholar]
- Jenkinson, D.S.; Coleman, K. Calculating the Annual Input of Organic Matter to Soil from Measurements of Total Organic Carbon and Radiocarbon. Eur. J. Soil Sci. 1994, 45, 167–174. [Google Scholar] [CrossRef]
- Beckett, P.H.T. Studies on Soil Potassium: I. Confirmation of the Ratio Law: Measurement of Potassium Potential. J. Soil Sci. 1964, 15, 1–8. [Google Scholar] [CrossRef]
- Hood-Nowotny, R.; Umana, N.H.-N.; Inselbacher, E.; Oswald-Lachouani, P.; Wanek, W. Alternative Methods for Measuring Inorganic, Organic, and Total Dissolved Nitrogen in Soil. Soil Sci. Soc. Am. J. 2010, 74, 1018–1027. [Google Scholar] [CrossRef]
- Sah, R.N.; Brown, P.H. Techniques for Boron Determination and Their Application to the Analysis of Plant and Soil Samples. Plant Soil 1997, 193, 15–33. [Google Scholar] [CrossRef]
- Basson, W.D.; Stanton, D.A.; Böhmer, R.G. Automated Procedure for the Simultaneous Determination of Phosphorus and Nitrogen in Plant Tissue. Analyst 1968, 93, 166–172. [Google Scholar] [CrossRef]
- Emmert, E.M. Field Method for Estimating Nitrate, Phosphate, and Potassium in Plants. Plant Physiol. 1932, 7, 315. [Google Scholar] [CrossRef]
- Williams, D.E.; Vlamis, J. Boron Contamination in Furnace Dry Ashing of Plant Material. Anal. Chem. 1961, 33, 967–968. [Google Scholar] [CrossRef]
- Chen, X.W.; Wu, L.; Luo, N.; Mo, C.H.; Wong, M.H.; Li, H. Arbuscular Mycorrhizal Fungi and the Associated Bacterial Community Influence the Uptake of Cadmium in Rice. Geoderma 2019, 337, 749–757. [Google Scholar] [CrossRef]
- Pramanik, K.; Mitra, S.; Sarkar, A.; Soren, T.; Maiti, T.K. Characterization of Cadmium-Resistant Klebsiella Pneumoniae MCC 3091 Promoted Rice Seedling Growth by Alleviating Phytotoxicity of Cadmium. Environ. Sci. Pollut. Res. 2017, 24, 24419–24437. [Google Scholar] [CrossRef]
- Orhan, E.; Esitken, A.; Ercisli, S.; Turan, M.; Sahin, F. Effects of Plant Growth Promoting Rhizobacteria (PGPR) on Yield, Growth and Nutrient Contents in Organically Growing Raspberry. Sci. Hortic. 2006, 111, 38–43. [Google Scholar] [CrossRef]
- Lindhard Pedersen, H. Organic Fruit Growing. K Lind, G Lafer, K Schloffer, G Innerhofer and H Meister. CABI Publishing, Wallingford, Oxon, UK, 2004. 281 Pp ISBN 0–85199–640-X. J. Sci. Food Agric. 2005, 85, 708. [Google Scholar] [CrossRef]
- Li, F.; Zheng, Y.; Tian, J.; Ge, F.; Liu, X.; Tang, Y.; Feng, C. Cupriavidus Sp. Strain Cd02-Mediated PH Increase Favoring Bioprecipitation of Cd2+ in Medium and Reduction of Cadmium Bioavailability in Paddy Soil. Ecotoxicol. Environ. Saf. 2019, 184, 109655. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Fan, M.; Wang, E.; Chen, W.; Wei, G. Interactions of Plant Growth-Promoting Rhizobacteria and Soil Factors in Two Leguminous Plants. Appl. Microbiol. Biotechnol. 2017, 101, 8485–8497. [Google Scholar] [CrossRef]
- Fall, D.; Diouf, D.; Zoubeirou, A.M.; Bakhoum, N.; Faye, A.; Sall, S.N. Effect of Distance and Depth on Microbial Biomass and Mineral Nitrogen Content under Acacia senegal (L.) Willd. Trees. J. Environ. Manag. 2012, 95, S260–S264. [Google Scholar] [CrossRef] [PubMed]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of Arbuscular Mycorrhizal Fungi in Plant Growth Regulation: Implications in Abiotic Stress Tolerance. Front. Plant Sci. 2019, 10, 1068. [Google Scholar] [CrossRef]
- Frey, S.D. Mycorrhizal Fungi as Mediators of Soil Organic Matter Dynamics. Annu. Rev. Ecol. Evol. Syst. 2019, 50, 237–259. [Google Scholar] [CrossRef]
- Zak, D.R.; Pellitier, P.T.; Argiroff, W.A.; Castillo, B.; James, T.Y.; Nave, L.E.; Averill, C.; Beidler, K.V.; Bhatnagar, J.; Blesh, J.; et al. Exploring the Role of Ectomycorrhizal Fungi in Soil Carbon Dynamics. New Phytol. 2019, 223, 33–39. [Google Scholar] [CrossRef]
- Fan, X.; Zhang, S.; Mo, X.; Li, Y.; Fu, Y.; Liu, Z. Effects of Plant Growth-Promoting Rhizobacteria and N Source on Plant Growth and N and P Uptake by Tomato Grown on Calcareous Soils. Pedosphere 2017, 27, 1027–1036. [Google Scholar] [CrossRef]
- Hamel, C. Impact of Arbuscular Mycorrhizal Fungi on N and P Cycling in the Root Zone. Can. J. Soil Sci. 2004, 84, 383–395. [Google Scholar] [CrossRef]
- Sagar, A.; Rathore, P.; Ramteke, P.W.; Ramakrishna, W.; Reddy, M.S.; Pecoraro, L. Plant Growth Promoting Rhizobacteria, Arbuscular Mycorrhizal Fungi and Their Synergistic Interactions to Counteract the Negative Effects of Saline Soil on Agriculture: Key Macromolecules and Mechanisms. Microorganisms 2021, 9, 1491. [Google Scholar] [CrossRef] [PubMed]
- Hussain, B.; Ashraf, M.N.; Shafeeq-ur-Rahman; Abbas, A.; Li, J.; Farooq, M. Cadmium Stress in Paddy Fields: Effects of Soil Conditions and Remediation Strategies. Sci. Total Environ. 2021, 754, 142188. [Google Scholar] [CrossRef] [PubMed]
- Masood, S.; Zhao, X.Q.; Shen, R.F. Bacillus Pumilus Promotes the Growth and Nitrogen Uptake of Tomato Plants under Nitrogen Fertilization. Sci. Hortic. 2020, 272, 109581. [Google Scholar] [CrossRef]
- Jha, Y.; Subramanian, R.B. Paddy Plants Inoculated with PGPR Show Better Growth Physiology and Nutrient Content under Saline Conditions. Chil. J. Agric. Res. 2013, 73, 213–219. [Google Scholar] [CrossRef]
- Khosravi, A.; Zarei, M.; Ronaghi, A. Effect of PGPR, Phosphate Sources and Vermicompost on Growth and Nutrients Uptake by Lettuce in a Calcareous Soil. J. Plant Nutr. 2018, 41, 80–89. [Google Scholar] [CrossRef]
- Nautiyal, C.S.; Srivastava, S.; Chauhan, P.S.; Seem, K.; Mishra, A.; Sopory, S.K. Plant Growth-Promoting Bacteria Bacillus Amyloliquefaciens NBRISN13 Modulates Gene Expression Profile of Leaf and Rhizosphere Community in Rice during Salt Stress. Plant Physiol. Biochem. 2013, 66, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Shah, G.; Jan, M.; Afreen, M.; Anees, M.; Rehman, S.; Daud, M.K.; Malook, I.; Jamil, M. Halophilic Bacteria Mediated Phytoremediation of Salt-Affected Soils Cultivated with Rice. J. Geochem. Explor. 2017, 174, 59–65. [Google Scholar] [CrossRef]
- Turan, M.; Gulluce, M.; Şahin, F. Effects of Plant-Growth-Promoting Rhizobacteria on Yield, Growth, and Some Physiological Characteristics of Wheat and Barley Plants. Commun. Soil Sci. Plant Anal. 2012, 43, 681739. [Google Scholar] [CrossRef]
- Mäder, P.; Kaiser, F.; Adholeya, A.; Singh, R.; Uppal, H.S.; Sharma, A.K.; Srivastava, R.; Sahai, V.; Aragno, M.; Wiemken, A.; et al. Inoculation of root microorganisms for sustainable wheat–rice and wheat–black gram rotations in India. Soil Biol. Biochem. 2011, 43, 609–619. [Google Scholar] [CrossRef]
- Saboor, A.; Ali, M.A.; Husain, S.; Tahir, M.S.; Irfan, M.; Bilal, M.; Baig, K.S.; Datta, R.; Ahmed, N.; Danish, S.; et al. Regulation of Phosphorus and Zinc Uptake in Relation to Arbuscular Mycorrhizal Fungi for Better Maize Growth. Agronomy 2021, 11, 2322. [Google Scholar] [CrossRef]
- Akhtar, M.J.; Asghar, H.N.; Shahzad, K.; Arshad, M. Role of plant growth promoting rhizobacteria applied in combination with compost and mineral fertilizers to improve growth and yield of wheat (Triticum aestivum L.). Pak. J. Bot. 2009, 41, 381–390. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Values |
|---|---|
| Soil pH | 8.2 |
| Soil EC (dS m−1) | 1.1 |
| Soil organic matter (%) | 0.8 |
| Soil nitrogen (%) | 0.04 |
| Plant available phosphorous (%) | 6.5 |
| Extractable potassium (%) | 180 |
| Soil B (%) | 0.39 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, D.; Saeed, M.; Ali, M.N.H.A.; Raheel, M.; Ashraf, W.; Hassan, Z.; Hassan, M.Z.; Farooq, U.; Hakim, M.F.; Rao, M.J.; et al. Plant Growth Promoting Rhizobacteria (PGPR) and Arbuscular Mycorrhizal Fungi Combined Application Reveals Enhanced Soil Fertility and Rice Production. Agronomy 2023, 13, 550. https://doi.org/10.3390/agronomy13020550
Chen D, Saeed M, Ali MNHA, Raheel M, Ashraf W, Hassan Z, Hassan MZ, Farooq U, Hakim MF, Rao MJ, et al. Plant Growth Promoting Rhizobacteria (PGPR) and Arbuscular Mycorrhizal Fungi Combined Application Reveals Enhanced Soil Fertility and Rice Production. Agronomy. 2023; 13(2):550. https://doi.org/10.3390/agronomy13020550
Chicago/Turabian StyleChen, Delai, Munawar Saeed, Mian Noor Hussain Asghar Ali, Muhammad Raheel, Waqas Ashraf, Zeshan Hassan, Muhammad Zeeshan Hassan, Umar Farooq, Muhammad Fahad Hakim, Muhammad Junaid Rao, and et al. 2023. "Plant Growth Promoting Rhizobacteria (PGPR) and Arbuscular Mycorrhizal Fungi Combined Application Reveals Enhanced Soil Fertility and Rice Production" Agronomy 13, no. 2: 550. https://doi.org/10.3390/agronomy13020550
APA StyleChen, D., Saeed, M., Ali, M. N. H. A., Raheel, M., Ashraf, W., Hassan, Z., Hassan, M. Z., Farooq, U., Hakim, M. F., Rao, M. J., Naqvi, S. A. H., Moustafa, M., Al-Shehri, M., & Negm, S. (2023). Plant Growth Promoting Rhizobacteria (PGPR) and Arbuscular Mycorrhizal Fungi Combined Application Reveals Enhanced Soil Fertility and Rice Production. Agronomy, 13(2), 550. https://doi.org/10.3390/agronomy13020550