

Detection of Resistance in Echinochloa spp. to Three Post-Emergence Herbicides (Penoxsulam, Metamifop, and Quinclorac) Used in China

 and
and 
Abstract
:1. Introduction
2. Materials & Methods
2.1. Plant Materials
2.2. Sensitivity of Different Biotypes to the Three Herbicides
2.3. Development of the RISQ Test
2.3.1. Media Preparation
2.3.2. Seedling Transplantation
2.3.3. Determination of Observation Time
2.3.4. Selection of Discriminating Rates of Herbicides
2.3.5. The Whole-Plant Pot Assay
2.4. Verification
2.5. Statistical Analysis
2.5.1. The Sensitivity of Different Biotypes
2.5.2. RISQ Test
3. Results
3.1. Susceptibility of Different Biotypes to the Three Herbicides
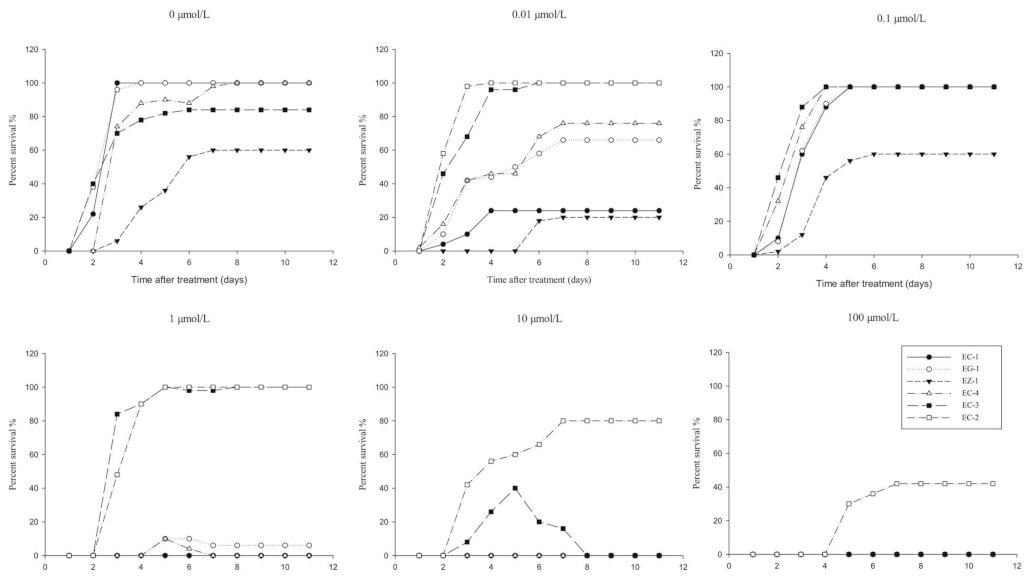
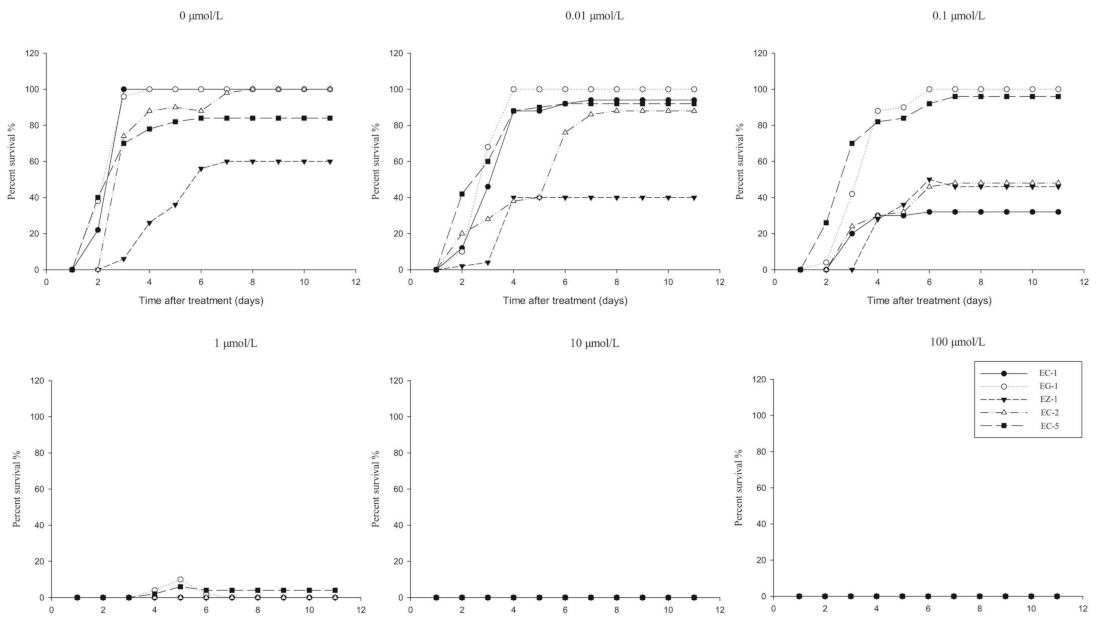
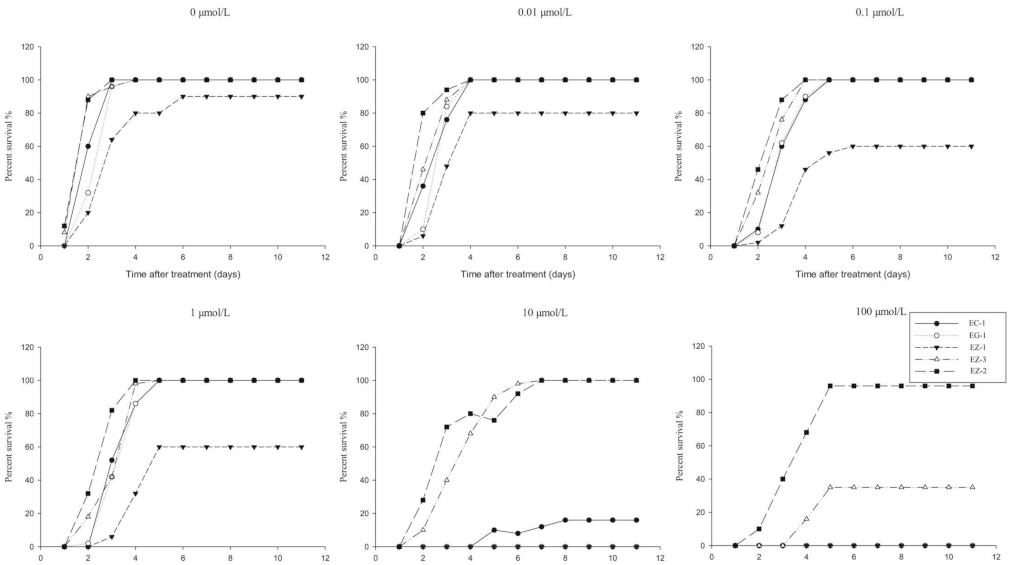
3.2. Development of the RISQ Test
3.2.1. Determination of Observation Time
3.2.2. Selection of Discriminating Rates of Herbicides in the RISQ Test
3.3. Verification
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maun, M.A.; Barrett, S.C.H. The biology of Canada weeds.: 77. Echinochloa-crussgalli (L.) Beauv. Can. J. Plant Sci. 1986, 66, 739–759. [Google Scholar] [CrossRef]
- Lim, S.H.; Kim, H.; Noh, T.K.; Lim, J.S.; Yook, M.J.; Kim, J.W.; Yi, J.H.; Kim, D. Baseline Sensitivity of Echinochloa crus-gall and E. oryzicola to Florpyrauxifen-Benzyl, a New Synthetic Auxin Herbicide, in Korea. Front. Plant Sci. 2021, 12, 656642. [Google Scholar] [CrossRef]
- Holm, L.G.; Pucknett, D.L.; Pancho, J.V.; Heberger, J.P. The World’s Worst Weeds. Distribution and Biology; University Press of Hawaii: Honolulu, HI, USA, 1977. [Google Scholar]
- Juliano, L.M.; Casimero, M.C.; Llewellyn, R. Multiple herbicide resistance in barnyardgrass (Echinochloa crus-galli) in direct-seeded rice in the Philippines. Int. J. Pest Manag. 2010, 56, 299–307. [Google Scholar] [CrossRef]
- Zhang, N.Y.; Linscombe, S.; Oard, J. Out-crossing frequency and genetic analysis of hybrids between transgenic glufosinate herbicide-resistant rice and the weed, red rice. Euphytica 2003, 130, 35–45. [Google Scholar] [CrossRef]
- Tadeo, J.L.; Sanchez-Brunete, C.; Perez, R.A.; Fernandez, M.D. Analysis of herbicide residues in cereals, fruits and vegetables. J. Chromatogr. A 2000, 882, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Tiryaki, O.; Temur, C. The Fate of Pesticide in the Environment. J. Biol. Environ. Sci. 2010, 4, 29–38. [Google Scholar]
- Quan, L.; Li, S.F.; Tian, S.J.; Xu, H.; Lin, A.Q.; Gu, L. Determination of organochlorine pesticides residue in ginseng root by orthogonal array design soxhlet extraction and gas chromatography. Chromatographia 2004, 59, 89–93. [Google Scholar]
- Beckie, H.J.; Ashworth, M.B.; Flower, K.C. Herbicide Resistance Management: Recent Developments and Trends. Plants 2019, 8, 161. [Google Scholar] [CrossRef] [Green Version]
- Heap, I. The International Herbicide-Resistant Weed Database. Available online: http://www.weedscience.org/Home.aspx (accessed on 13 February 2023).
- Ryan, G.F. Resistance of common groundsel to simazine and atrazine. Weed Sci. 1970, 18, 614–616. [Google Scholar] [CrossRef]
- Carey, V.F.; Hoagland, R.E.; Talbert, R.E. Resistance mechanism of propanil-resistant barnyardgrass: 2. In-vivo metabolism of the propanil molecule. Pestic. Sci. 1997, 49, 333–338. [Google Scholar] [CrossRef]
- Cai, X.; Chen, J.; Wang, X.; Gao, H.; Xiang, B.; Dong, L. Mefenacet resistance in multiple herbicide-resistant Echinochloa crus-galli L. populations. Pestic. Biochem. Physiol. 2022, 182, 105038. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Lv, B.; Wang, Q.; Li, J.; Dong, L. A resistance mechanism dependent upon the inhibition of ethylene biosynthesis. Pest Manag. Sci. 2013, 69, 1407–1414. [Google Scholar] [CrossRef] [PubMed]
- Bagavathiannan, M.V.; Norsworthy, J.K.; Smith, K.L.; Neve, P. Modeling the Simultaneous Evolution of Resistance to ALS- and ACCase-Inhibiting Herbicides in Barnyardgrass (Echinochloa crusgalli) in Clearfield (R) Rice. Weed Technol. 2014, 28, 89–103. [Google Scholar] [CrossRef]
- Fang, J.; Zhang, Y.; Liu, T.; Yan, B.; Li, J.; Dong, L. Target-Site and Metabolic Resistance Mechanisms to Penoxsulam in Barnyardgrass (Echinochloa crus-galli (L.) P. Beauv). J. Agric. Food Chem. 2019, 67, 8085–8095. [Google Scholar] [CrossRef]
- Zhao, Z.; Liu, X.; Pan, X.; Napier, R.; Dong, L.; Li, J. Different regulation of auxin homeostasis would be a possible mechanism conferring quinclorac resistance in Echinochloa crusgalli var. zelayensis. Weed Res. 2022, 62, 318–327. [Google Scholar] [CrossRef]
- Burgos, N.R.; Tranel, P.J.; Streibig, J.C.; Davis, V.M.; Shaner, D.; Norsworthy, J.K.; Ritz, C. Review: Confirmation of Resistance to Herbicides and Evaluation of Resistance Levels. Weed Sci. 2013, 61, 4–20. [Google Scholar] [CrossRef]
- Tataridas, A.; Jabran, K.; Kanatas, P.; Oliveira, R.S.; Freitas, H.; Travlos, I. Early detection, herbicide resistance screening, and integrated management of invasive plant species: A review. Pest Manag. Sci. 2022, 78, 3957–3972. [Google Scholar] [CrossRef] [PubMed]
- Kaundun, S.S.; Hutchings, S.J.; Dale, R.P.; Bailly, G.C.; Glanfield, P. Syngenta ‘RISQ’ test: A novel in-season method for detecting resistance to post-emergence ACCase and ALS inhibitor herbicides in grass weeds. Weed Res. 2011, 51, 284–293. [Google Scholar] [CrossRef]
- Kaundun, S.S.; Hutchings, S.-J.; Harris, S.C.; Jackson, L.V.; Shashi-Kiran, R.; Dale, R.P.; McIndoe, E. A Simple In-Season Bioassay for Detecting Glyphosate Resistance in Grass and Broadleaf Weeds Prior to Herbicide Application in the Field. Weed Sci. 2014, 62, 597–607. [Google Scholar] [CrossRef]
- Khammassi, M.; Hajri, H.; Menchari, Y.; Chaabane, H.; Souissi, T. Current status in resistance to ACCase and ALS-inhibiting herbicides in rigid ryegrass populations from cereal crops in North of Tunisia. J. Agric. Sci. 2019, 157, 676–683. [Google Scholar] [CrossRef]
- Weiqiang, J.; Roh, S.-W.; Song, J.E.; Bo, A.B.; Cho, K.M.; Park, K.W.; Joo, L. Evaluation of Three in-season Tests for Rapidly Detecting Resistance to Acetolactate Synthase Inhibitors in Schoenoplectus juncoides. Weed Turfgrass Sci. 2021, 10, 13–23. [Google Scholar]
- Koger, C.H.; Shaner, D.L.; Henry, W.B.; Nadler-Hassar, T.; Thomas, W.E.; Wilcut, J.W. Assessment of two nondestructive assays for detecting glyphosate resistance in horseweed (Conyza canadensis). Weed Sci. 2005, 53, 438–445. [Google Scholar] [CrossRef]
- Chung, I.M.; Ahn, J.K.; Yun, S.J. Assessment of allelopathic potential of barnyard grass (Echinochloa crus-galli) on rice (Oryza sativa L.) cultivars. Crop Prot. 2001, 20, 921–928. [Google Scholar] [CrossRef]
- Beckie, H.J.; Tardif, F.J. Herbicide cross resistance in weeds. Crop Prot. 2012, 35, 15–28. [Google Scholar] [CrossRef]
- Delye, C. Weed resistance to acetyl coenzyme A carboxylase inhibitors: An update. Weed Sci. 2005, 53, 728–746. [Google Scholar] [CrossRef]
- Beckie, H.J.; Reboud, X. Selecting for Weed Resistance: Herbicide Rotation and Mixture. Weed Technol. 2009, 23, 363–370. [Google Scholar] [CrossRef]
- Rosenhauer, M. Distribution, Detection and Genetic Background of Herbicide-Resistant Alopecurus myosuroides (Huds.) in Germany. Ph.D. Dissertation, Universität Hohenheim, Stuttgart, Germany, 2015. [Google Scholar]
- Preston, C. Herbicide resistance in weeds endowed by enhanced detoxification: Complications for management. Weed Sci. 2004, 52, 448–453. [Google Scholar] [CrossRef]
- Ghanizadeh, H.; Harrington, K.C.; James, T.K.; Woolley, D.J. A quick test using seeds for detecting dicamba resistance in fathen (Chenopodium album). Aust. J. Crop Sci. 2015, 9, 337–343. [Google Scholar]
- Richter, J.; Powles, S.B. Pollen Expression of Herbicide Target Site Resistance Genes in Annual Ryegrass (Lolium rigidum). Plant Physiol. 1993, 102, 1037–1041. [Google Scholar] [CrossRef] [Green Version]
- Letouze, A.; Gasquez, J. A pollen test to detect ACCase target-site resistance within Alopecurus myosuroides populations. Weed Res. 2000, 40, 151–162. [Google Scholar] [CrossRef]
- Cirujeda, A.; Recasens, J.; Taberner, A. A qualitative quick-test for detection of herbicide resistance to tribenuron-methyl in Papaver rhoeas. Weed Res. 2001, 41, 523–534. [Google Scholar] [CrossRef]
- Cirujeda, A.; Taberner, A. A qualitative quick-test for the detection of herbicide resistance towards tribenuron-methyl in Papaver rhoeas L. on agar medium. In Proceedings of the Actas Congreso 2001 Sociedad Española de Malherbología, León, Spain, 20–22 November 2001. [Google Scholar]
- Concenco, G.; Melo, P.T.B.S.; Andres, A.; Ferreira, E.A.; Galon, L.; Ferreira, F.A.; Silva, A.A. A quick method for detecting resistance of Echinochloa spp. to quinclorac. Planta Daninha 2008, 26, 429–437. [Google Scholar]
- Abdurruhman, A.M.; Uygur, S.; Mathiassen, S.K.; Uygur, N. Identification of sterile wild oat (Avena sterilis L.) resistance to acetolactate synthase (ALS)-inhibiting herbicides using different assay techniques. J. Plant Prot. Res. 2020, 60, 244–252. [Google Scholar]
- Zhang, C.J.; Lim, S.H.; Kim, J.W.; Nah, G.; Fischer, A.; Kim, D.S. Leaf chlorophyll fluorescence discriminates herbicide resistance in Echinochloa species. Weed Res. 2016, 56, 424–433. [Google Scholar] [CrossRef]
- Zhang, C.-J.; Kim, D.-S. Using Leaf Chlorophyll Fluorescence for In-Season Diagnosing Herbicide Resistance in Echinochloa Species at Reproductive Growth Stage. Plant Prot. Sci. 2018, 54, 194–202. [Google Scholar] [CrossRef] [Green Version]
- Linn, A.I.; Kosnarova, P.; Soukup, J.; Gerhards, R. Detecting herbicide-resistant Apera spica-venti with a chlorophyll fluorescence agar test. Plant Soil Environ. 2018, 64, 386–392. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Peteinatos, G.; Li, H.; Gerhards, R. Rapid in-season detection of herbicide resistant Alopecurus myosuroides using a mobile fluorescence imaging sensor. Crop Prot. 2016, 89, 170–177. [Google Scholar] [CrossRef]
- Kaiser, Y.I.; Menegat, A.; Gerhards, R. Chlorophyll fluorescence imaging: A new method for rapid detection of herbicide resistance in Alopecurus myosuroides. Weed Res. 2013, 53, 399–406. [Google Scholar] [CrossRef]
- Zhang, T.-J.; Feng, L.; Tian, X.-S.; Yang, C.-H.; Gao, J.-D. Use of chlorophyll fluorescence and P700 absorbance to rapidly detect glyphosate resistance in goosegrass (Eleusine indica). J. Integr. Agric. 2015, 14, 714–723. [Google Scholar] [CrossRef]
- Shaner, D.L.; Nadler-Hassar, T.; Henry, W.B.; Koger, C.H. A rapid in vivo shikimate accumulation assay with excised leaf discs. Weed Sci. 2005, 53, 769–774. [Google Scholar] [CrossRef]
- Pan, L.; Li, J.; Xia, W.; Zhang, D.; Dong, L. An effective method, composed of LAMP and dCAPS, to detect different mutations in fenoxaprop-P-ethyl-resistant American sloughgrass (Beckmannia syzigachne Steud.) populations. Pestic. Biochem. Physiol. 2015, 117, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Li, J.; Zhang, W.-N.; Dong, L. Detection of the I1781L mutation in fenoxaprop-p-ethyl-resistant American sloughgrass (Beckmannia syzigachne Steud.), based on the loop-mediated isothermal amplification method. Pest Manag. Sci. 2015, 71, 123–130. [Google Scholar] [CrossRef]
- Panozzo, S.; Farinati, S.; Sattin, M.; Scarabel, L. Can allele-specific loop-mediated isothermal amplification be used for rapid detection of target-site herbicide resistance in Lolium spp.? Plant Methods 2023, 19, 14. [Google Scholar] [CrossRef] [PubMed]
- Ghanizadeh, H.; Griffiths, A.G.; Buddenhagen, C.E.; Anderson, C.B.; Harrington, K.C. A PCR plus restriction enzyme-based technique for detecting target-enzyme mutations at position Pro-106 in glyphosate-resistant Lolium perenne . PLoS ONE 2021, 16, e0246028. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Wu, Q.; Wan, F.; Guo, J.; Wang, R. Reliable and rapid identification of glyphosate-resistance in the invasive weed Amaranthus palmeri in China. Pest Manag. Sci. 2022, 78, 2173–2182. [Google Scholar] [CrossRef]
- Alarcon-Reverte, R.; Hanley, S.; Kaundun, S.S.; Karp, A.; Moss, S.R. A SNaPshot assay for the rapid and simple detection of known point mutations conferring resistance to ACCase-inhibiting herbicides in Lolium spp. Weed Res. 2013, 53, 12–20. [Google Scholar] [CrossRef]
- Benevenuto, J.; Bhakta, M.; Lohr, D.A.; Ferrao, L.F.V.; Resende, M.F.R., Jr.; Kirst, M.; Quesenberry, K.; Munoz, P. Cost-effective detection of genome-wide signatures for 2,4-D herbicide resistance adaptation in red clover. Sci. Rep. 2019, 9, 20037. [Google Scholar] [CrossRef] [Green Version]
- Delye, C.; Causse, R.; Gautier, V.; Poncet, C.; Michel, S. Using next-generation sequencing to detect mutations endowing resistance to pesticides: Application to acetolactate-synthase (ALS)-based resistance in barnyard grass, a polyploid grass weed. Pest Manag. Sci. 2015, 71, 675–685. [Google Scholar] [CrossRef]
- Petit, C.; Bay, G.; Pernin, F.; Delye, C. Prevalence of cross- or multiple resistance to the acetyl-coenzyme A carboxylase inhibitors fenoxaprop, clodinafop and pinoxaden in black-grass (Alopecurus myosuroides Huds.) in France. Pest Manag. Sci. 2010, 66, 168–177. [Google Scholar] [CrossRef]
- Yoon, A.M.; Kim, I.R.; Choi, W. Monoclonal antibody production for CP4 EPSPS detection assays. Korean J. Environ. Biol. 2021, 39, 445–451. [Google Scholar] [CrossRef]
- Jones, E.A.L.; Austin, R.; Dunne, J.C.; Cahoon, C.W.; Jennings, K.M.; Leon, R.G.; Everman, W.J. Utilization of image-based spectral reflectance to detect herbicide resistance in glufosinate-resistant and glufosinate-susceptible plants: A proof of concept. Weed Sci. 2022, 1–11. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Biotypes * | Species | Origin | Susceptibility |
|---|---|---|---|
| EC-1 (JNBX-1) | E. crusgalli | Baixia, Jiangsu province in China (33°49′ N, 119°23′ E) | susceptible to penoxsulam, metamifop, and quinclorac |
| EG-1 (JJXY-1) | E. glabrescens | Minhe, Xiannv, Jiangdu, Jiangsu province in China (32°42′ N, 119°63′ E) | susceptible to penoxsulam, metamifop, and quinclorac |
| EZ-1 (JNX-S) | E. crusgalli var. zelayensis | Academy of Agricultural Sciences, Jiangsu province in China (32°04′ N, 118°86′ E) | resistant to quinclorac |
| EC-2 (AXXZ-6) | E. crusgalli | Xuanzhou, Xuancheng, Anhui province in China (30°47′ N, 118°14′ E) | resistant to penoxsulam |
| EC-3 (AXXZ-8) | E. crusgalli | Xuanzhou, Xuancheng, Anhui province in China(30°47′ N, 118°14′ E) | resistant to penoxsulam |
| EC-4 (JHLS-1) | E. crusgalli | Lianshui, Huaian, Jiangsu province in China (33°25′ N, 119°30′ E) | resistant to penoxsulam |
| EC-5 (AXXZ-2) | E. crusgalli | Xuanzhou, Xuancheng, Anhui province in China(30°56′ N, 110°59′ E) | resistant to penoxsulam |
| EZ-2 (SSX-R) | E. crusgalli var. zelayensis | Xinbing, Songjiang, Shanghai in China (31°03′ N, 121°23′ E) | resistant to quinclorac |
| EZ-3 (JCW-R) | E. crusgalli var. zelayensis | Wujin, Changzhou, Jiangsu province in China (31°70′ N, 119°94′ E) | resistant to quinclorac |
| EC-6 (JNLS-1) | E. crusgalli | Lishui, Jiangsu province in China (31°32′ N,118°89′ E) | susceptible to penoxsulam, metamifop, and quinclorac |
| EC-7 (HYYJ-1) | E. crusgalli | Yuanjiang, Hunan province in China (28°84′ N,112°35′ E) | resistant to penoxsulam and quinclorac |
| Herbicides | Herbicide Doses (g a.i./ha) | Biotypes |
|---|---|---|
| penoxsulam | 0, 2.8125, 5.625, 11.25, 22.5, 45 | EC-1, EZ-1, EG-1, EC-2, EC-4 |
| 0, 5.625, 11.25, 22.5, 45, 90 | EC-3 | |
| 0, 3.75, 7.5, 15, 30, 60 | EC-6, EC-7 | |
| metamifop | 0, 12.5, 25, 50, 100, 200 | EC-1, EZ-1, EG-1, EC-2, EC-2 |
| 0, 7.5, 15, 30, 60, 120, 240 | EC-6, EC-7 | |
| quinclorac | 0, 75, 150, 300, 600, 1200 | EC-1, EZ-1 EG-1, EZ-3 |
| 0, 150, 300, 600, 1200, 2400 | EZ-2 | |
| 0, 46.88, 93.75, 187.5, 375, 750, 1500 | EC-6, EC-7 |
| Herbicides | Herbicide Rates (µmol/L) | Tested Biotypes |
|---|---|---|
| penoxsulam | 0, 0.3, 0.6, 0.9, 1.2, 1.5 | EC-1, EZ-1, EG-1, EC-2, EC-3, EC-4 |
| metamifop | 0, 0.3, 0.6, 0.9, 1.2, 1.5 | EC-1, EZ-1, EG-1, EC-2, EC-5 |
| quinclorac | 0, 0.6, 1.2, 2.4, 4.8, 9.6 | EC-1, EZ-1, EG-1, EZ-2, EZ-3 |
| Population | ED50 ± SE (g a.i./ha) | RI | Susceptibility |
|---|---|---|---|
| EC-1 | 3.10 ± 0.32 | 1.00 | S |
| EG-1 | 5.67 ± 1.57 | 1.83 | S |
| EZ-1 | 6.10 ± 0.60 | 1.97 | S |
| EC-4 | 10.40 ± 1.79 | 3.35 | L |
| EC-3 | 58.29 ± 5.68 | 18.80 | H |
| EC-2 | 40.45 ± 4.14 | 13.05 | H |
| EC-6 | <3.75 | <2 | S |
| EC-7 | 28.61 ± 3.53 | 9.23 | M |
| Population | ED50 ± SE (g a.i./ha) | RI | Susceptibility |
|---|---|---|---|
| EC-1 | 15.59 ± 3.81 | 1.00 | S |
| EG-1 | 29.65 ± 3.76 | 1.90 | S |
| EZ-1 | 29.96 ± 1.79 | 1.92 | S |
| EC-2 | 63.09 ± 7.69 | 4.05 | L |
| EC-5 | 51.85 ± 12.43 | 3.33 | L |
| EC-6 | <7.5 | <2 | S |
| EC-7 | 22.10 ± 3.41 | 1.42 | S |
| Population | ED50 ± SE (g a.i./ha) | RI | Susceptibility |
|---|---|---|---|
| EC-1 | 23.89 ± 1.66 | 1.00 | S |
| EG-1 | 42.26 ± 2.80 | 1.77 | S |
| EZ-1 | 94.96 ± 1.29 | 3.97 | L |
| EZ-2 | 2457.52 ± 149.69 | 102.87 | VH |
| EZ-3 | 416.79 ± 3.05 | 17.45 | H |
| EC-6 | <46.88 | <2 | S |
| EC-7 | 354.36 ± 73.33 | 14.83 | H |
| (a) penoxsulam | ||||||||
| Biotypes | Whole-Plant Pot Test (g a.i./ha) | Herbicide Rate in RISQ Test (µmol/L) | p-Value * | |||||
| 22.5 | 0 | 0.3 | 0.6 | 0.9 | 1.2 | 1.5 | ||
| EC-1 | 0 ± 0.00 | 100 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | >0.9999 |
| EG-1 | 6 ± 2.45 | 100 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0.2424 |
| EZ-1 | 0 ± 0.00 | 100 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | >0.9999 |
| EC-2 | 100 ± 0.00 | 100 ± 0.00 | 100 ± 0.00 | 72 ± 5.83 | 54 ± 5.10 | 0 ± 0.00 | 0 ± 0.00 | >0.9999 |
| EC-3 | 100 ± 0.00 | 100 ± 0.00 | 100 ± 0.00 | 52 ± 3.74 | 44 ± 5.10 | 2 ± 2.00 | 0 ± 0.00 | >0.9999 |
| EC-4 | 30 ± 3.16 | 100 ± 0.00 | 22 ± 3.74 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0.1609 |
| (b) metamifop | ||||||||
| Biotypes | Whole-Plant Pot Test (g a.i./ha) | Herbicide Rate in RISQ Test (µmol/L) | p-Value ** | |||||
| 100 ± 0.00 | 0 | 0.3 | 0.6 | 0.9 | 1.2 | 1.5 | ||
| EC-1 | 0 ± 0.00 | 100 ± 0.00 | 34 ± 5.19 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | >0.9999 |
| EG-1 | 0 ± 0.00 | 100 ± 0.00 | 14 ± 2.45 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | >0.9999 |
| EZ-1 | 0 ± 0.00 | 100 ± 0.00 | 30 ± 3.16 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | >0.9999 |
| EC-2 | 32 ± 3.74 | 100 ± 0.00 | 66 ± 5.10 | 28 ± 3.74 | 2 ± 2.00 | 0 ± 0.00 | 0 ± 0.00 | 0.6753 |
| EC-5 | 16 ± 2.45 | 100 ± 0.00 | 70 ± 3.16 | 6 ± 2.45 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | 0.1997 |
| (c) quinclorac | ||||||||
| Biotypes | Whole-Plant Pot Test (g a.i./ha) | Herbicide Rate in RISQ Test (µmol/L) | p-Value *** | |||||
| 300 | 0 | 0.6 | 1.2 | 2.4 | 4.8 | 9.6 | ||
| EC-1 | 0 ± 0.00 | 100 ± 0.00 | 78 ± 3.74 | 26 ± 2.45 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | >0.9999 |
| EG-1 | 0 ± 0.00 | 100 ± 0.00 | 92 ± 3.74 | 26 ± 4.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | >0.9999 |
| EZ-1 | 0 ± 0.00 | 100 ± 0.00 | 72 ± 3.74 | 22 ± 2.00 | 0 ± 0.00 | 0 ± 0.00 | 0 ± 0.00 | >0.9999 |
| EZ-2 | 100 ± 0.00 | 100 ± 0.00 | 100 ± 0.00 | 100 ± 0.00 | 100 ± 0.00 | 100 ± 0.00 | 100 ± 0.00 | >0.9999 |
| EZ-3 | 50 ± 0.32 | 100 ± 0.00 | 100 ± 0.00 | 88 ± 3.74 | 52 ± 3.74 | 2 ± 2.45 | 0 ± 0.00 | >0.9999 |
| Population | Characteristics | Origin | RISQ Test | |||
|---|---|---|---|---|---|---|
| Untreated | Penoxsulam 0.3 µmol/L | Metamifop 0.6 µmol/L | Quinclorac 2.4 µmol/L | |||
| EC-6 | suspected resistant field | Lishui, Jiangsu province in China | 100 ± 0.00 | 2 ± 0.20 | 3 ± 0.20 | 18 ± 0.66 |
| EC-7 | suspected resistant field | Yuanjiang, Hunan province in China | 100 ± 0.00 | 64 ± 0.60 | 6 ± 0.40 | 62 ± 0.58 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Wu, M.; Bao, S.; Li, J.; Liu, D.; Dong, L.; Li, J. Detection of Resistance in Echinochloa spp. to Three Post-Emergence Herbicides (Penoxsulam, Metamifop, and Quinclorac) Used in China. Agronomy 2023, 13, 841. https://doi.org/10.3390/agronomy13030841
Zhang Y, Wu M, Bao S, Li J, Liu D, Dong L, Li J. Detection of Resistance in Echinochloa spp. to Three Post-Emergence Herbicides (Penoxsulam, Metamifop, and Quinclorac) Used in China. Agronomy. 2023; 13(3):841. https://doi.org/10.3390/agronomy13030841
Chicago/Turabian StyleZhang, Yangyang, Miao Wu, Shuijing Bao, Jiamin Li, Dingrong Liu, Liyao Dong, and Jun Li. 2023. "Detection of Resistance in Echinochloa spp. to Three Post-Emergence Herbicides (Penoxsulam, Metamifop, and Quinclorac) Used in China" Agronomy 13, no. 3: 841. https://doi.org/10.3390/agronomy13030841
APA StyleZhang, Y., Wu, M., Bao, S., Li, J., Liu, D., Dong, L., & Li, J. (2023). Detection of Resistance in Echinochloa spp. to Three Post-Emergence Herbicides (Penoxsulam, Metamifop, and Quinclorac) Used in China. Agronomy, 13(3), 841. https://doi.org/10.3390/agronomy13030841