




Morphological, Anatomical and Chemical Characterization of Ricinus communis L. (Euphorbiaceae)

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
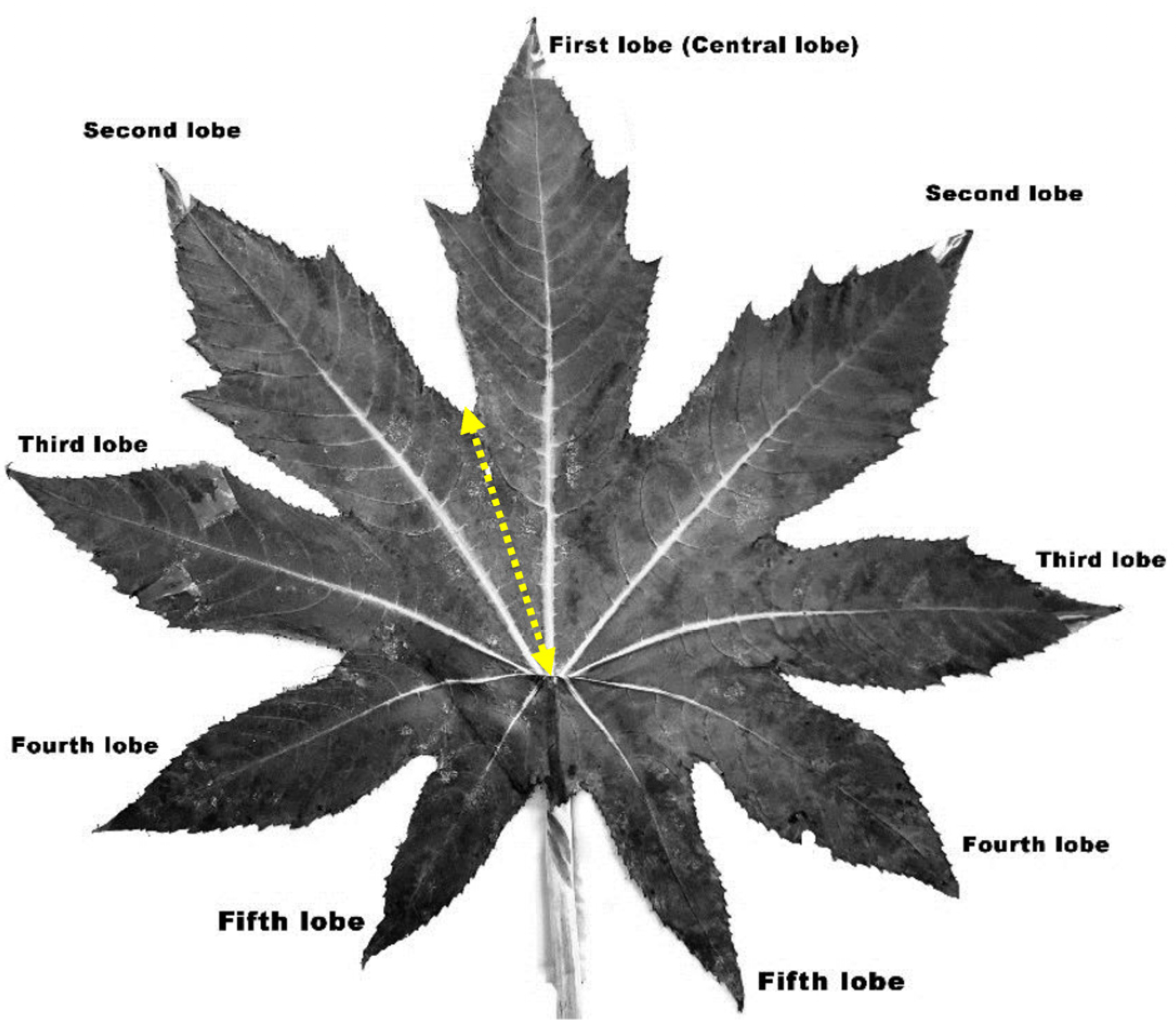
2.1. Plant Materials and Vegetative Morphometry
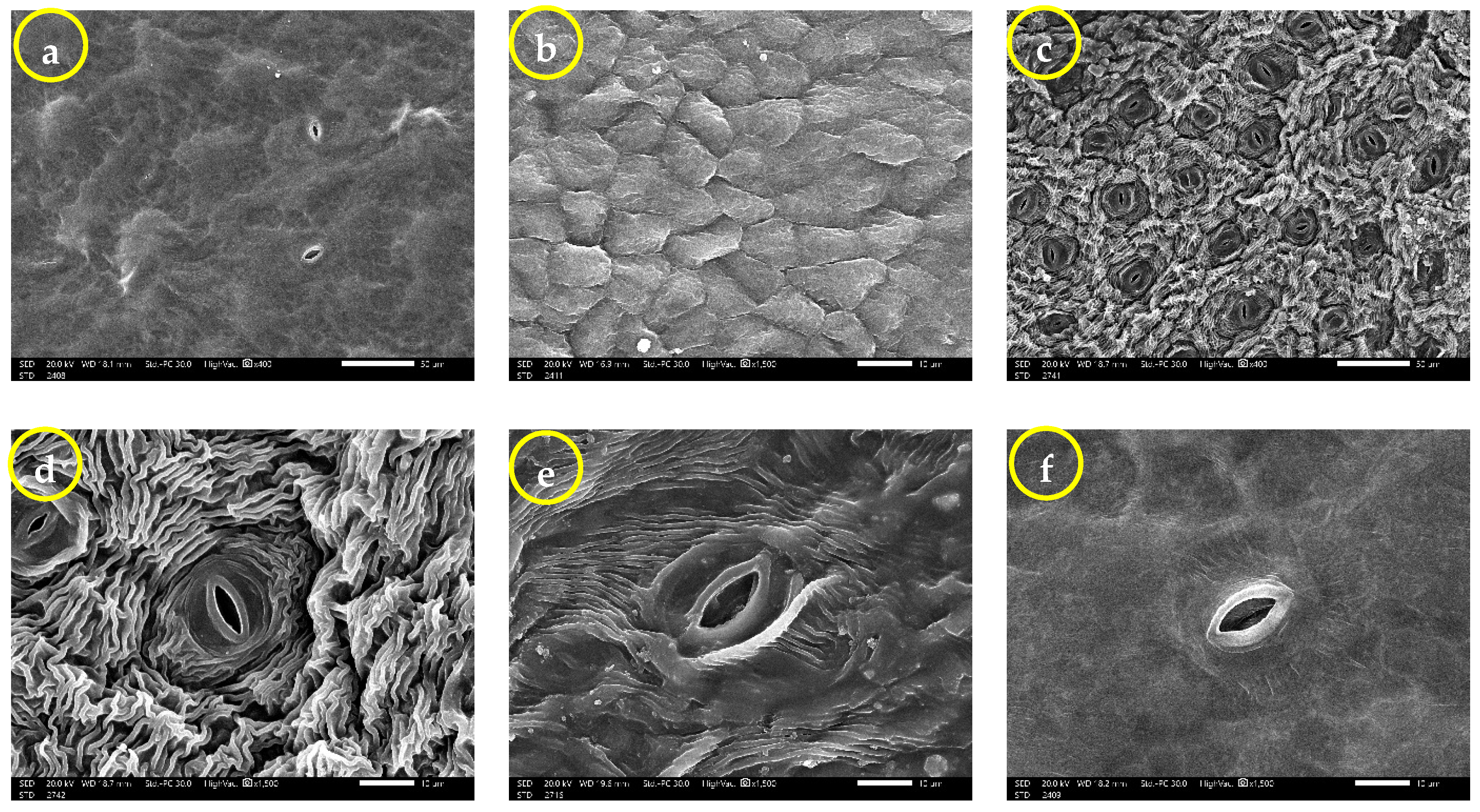
2.2. Leaf Surface Ultrastructure (SEM)
2.3. Soil Sampling and Elemental Analysis
2.4. Gas Chromatography-Mass Spectrometry (GC-MS) Analysis
2.4.1. Preparation of Plant Extracts
2.4.2. GC-MS Analysis Conditions
2.5. Data Analysis
3. Results
3.1. Vegetative Morphometry
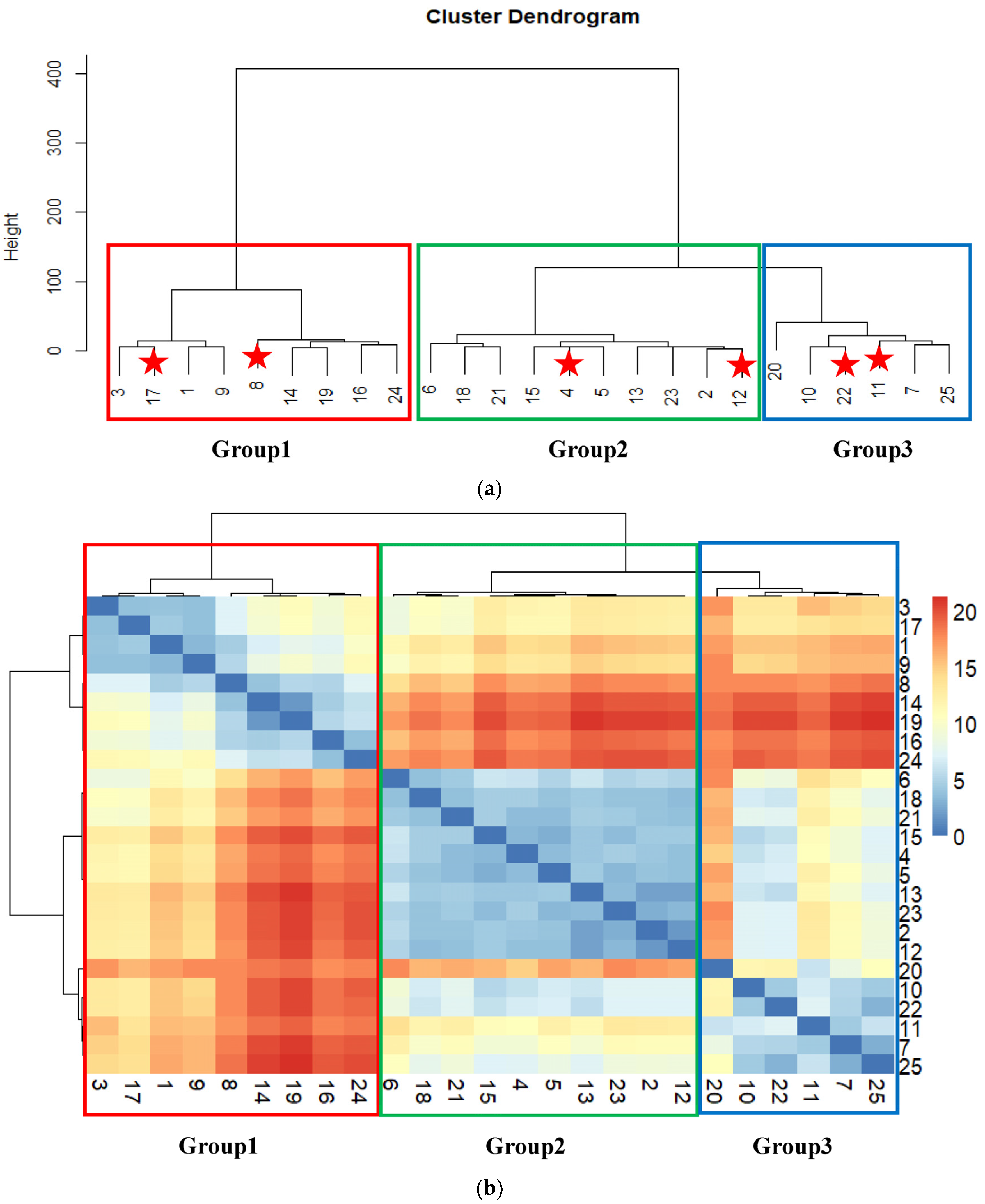
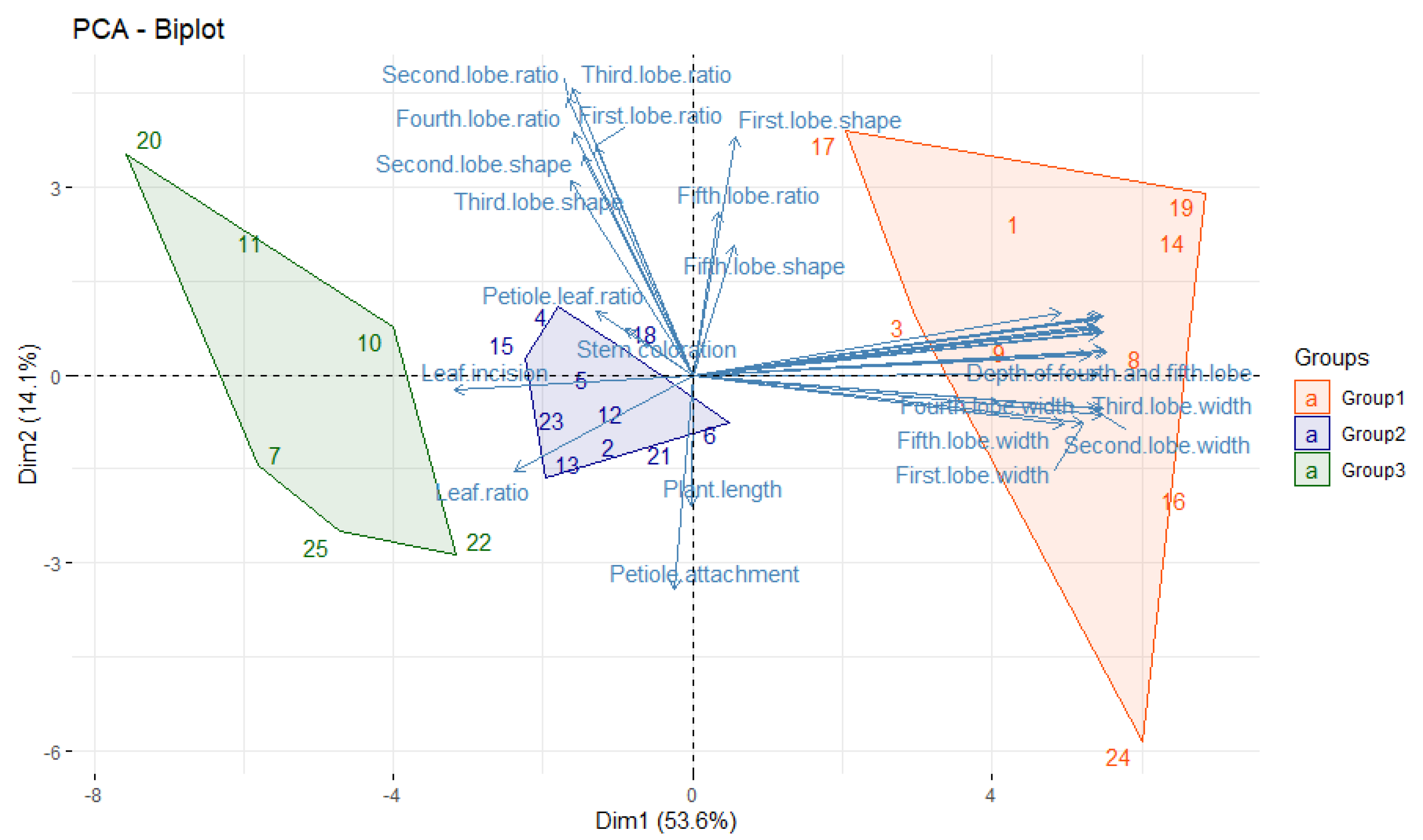
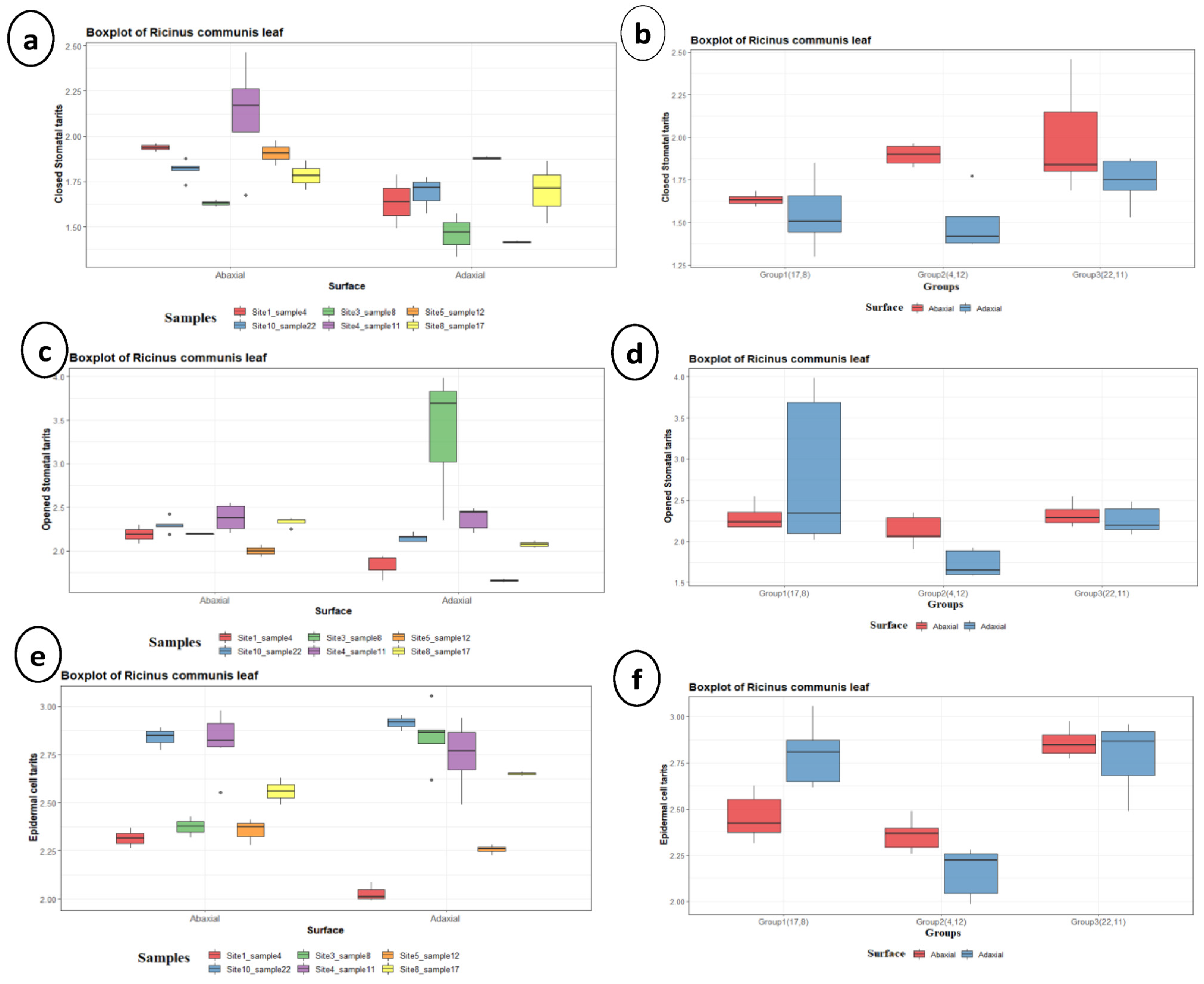
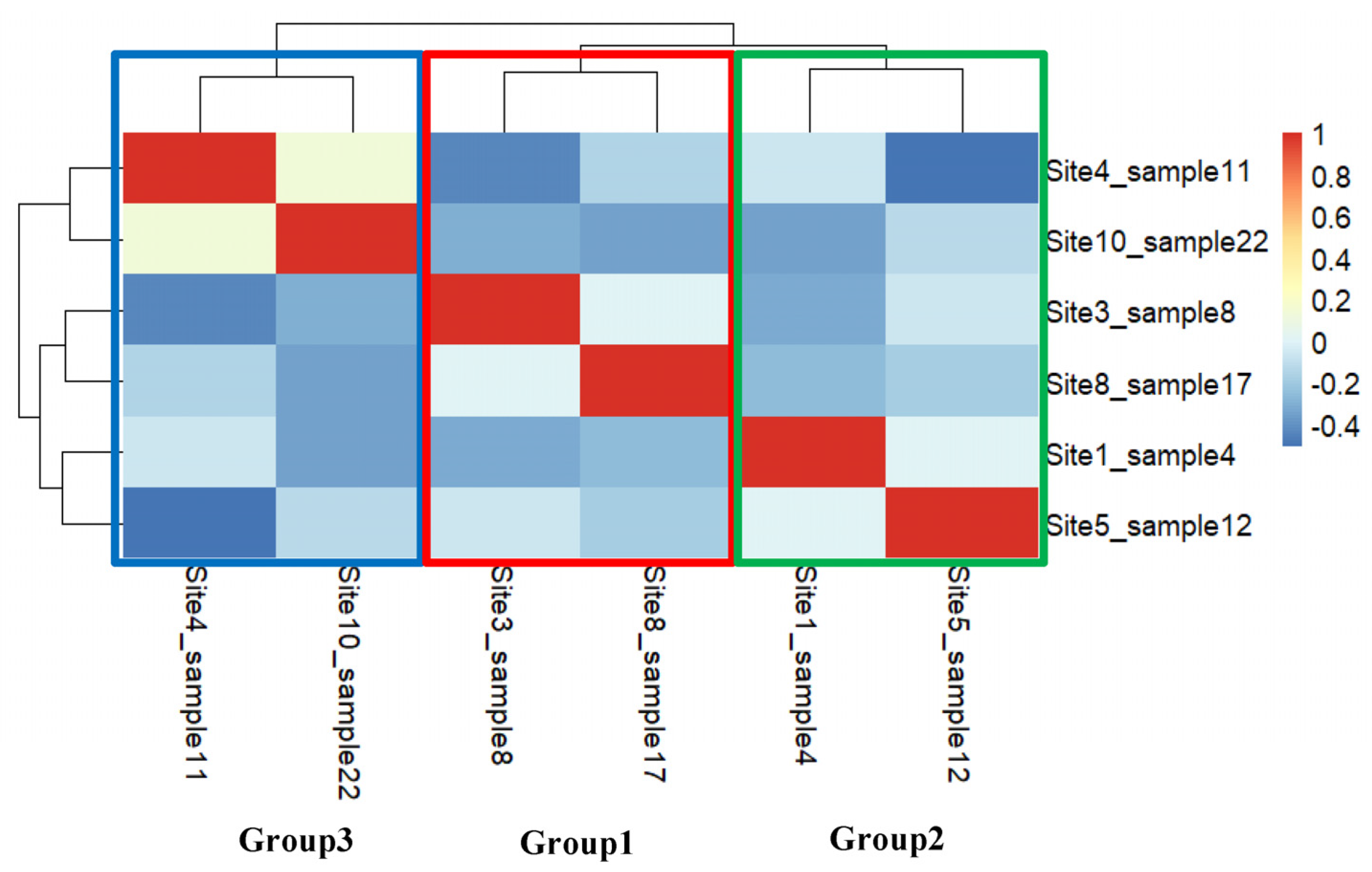
3.2. Multivariate Analysis
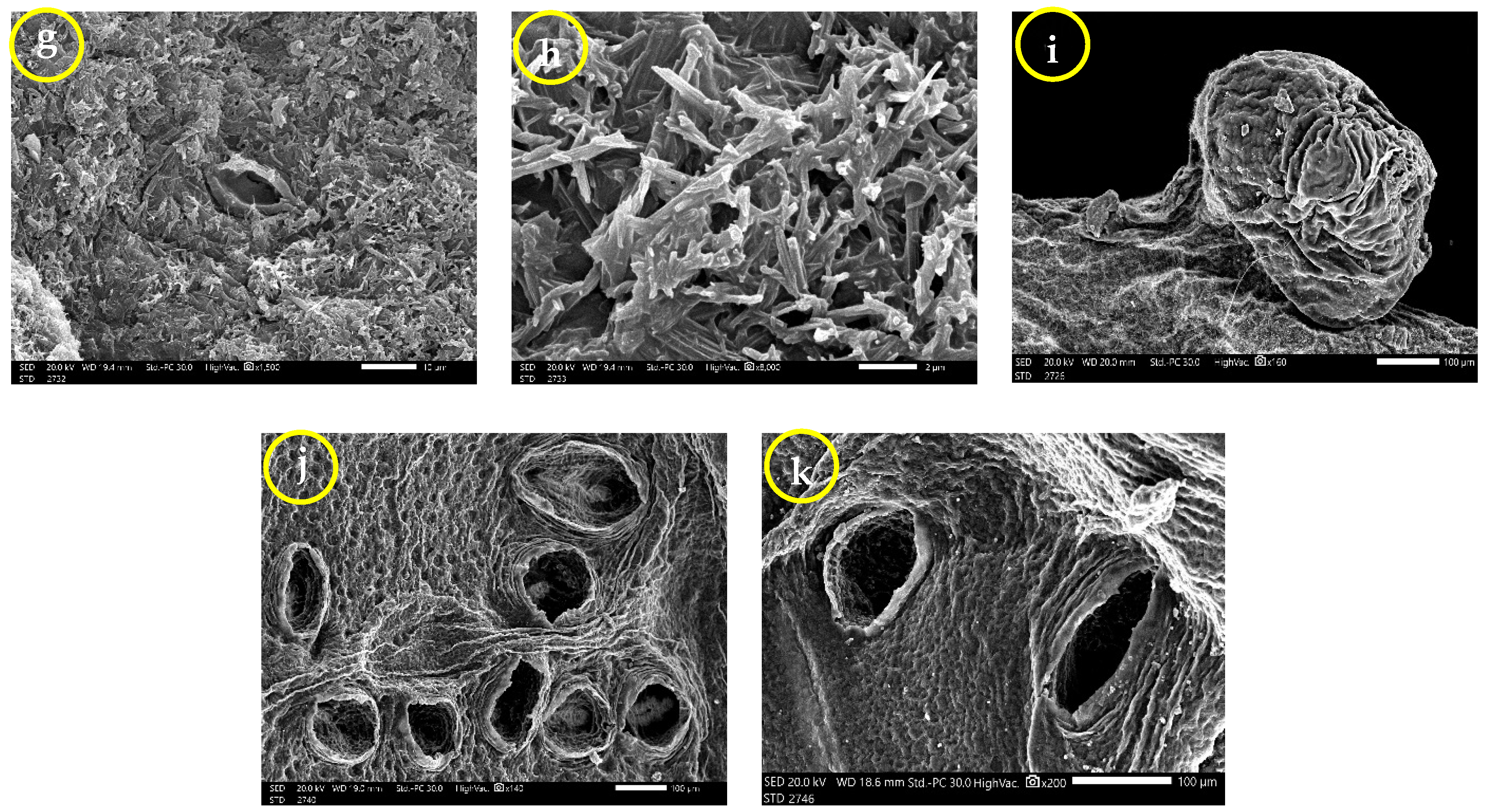
3.3. Leaf Surface Ultrastructure (SEM)
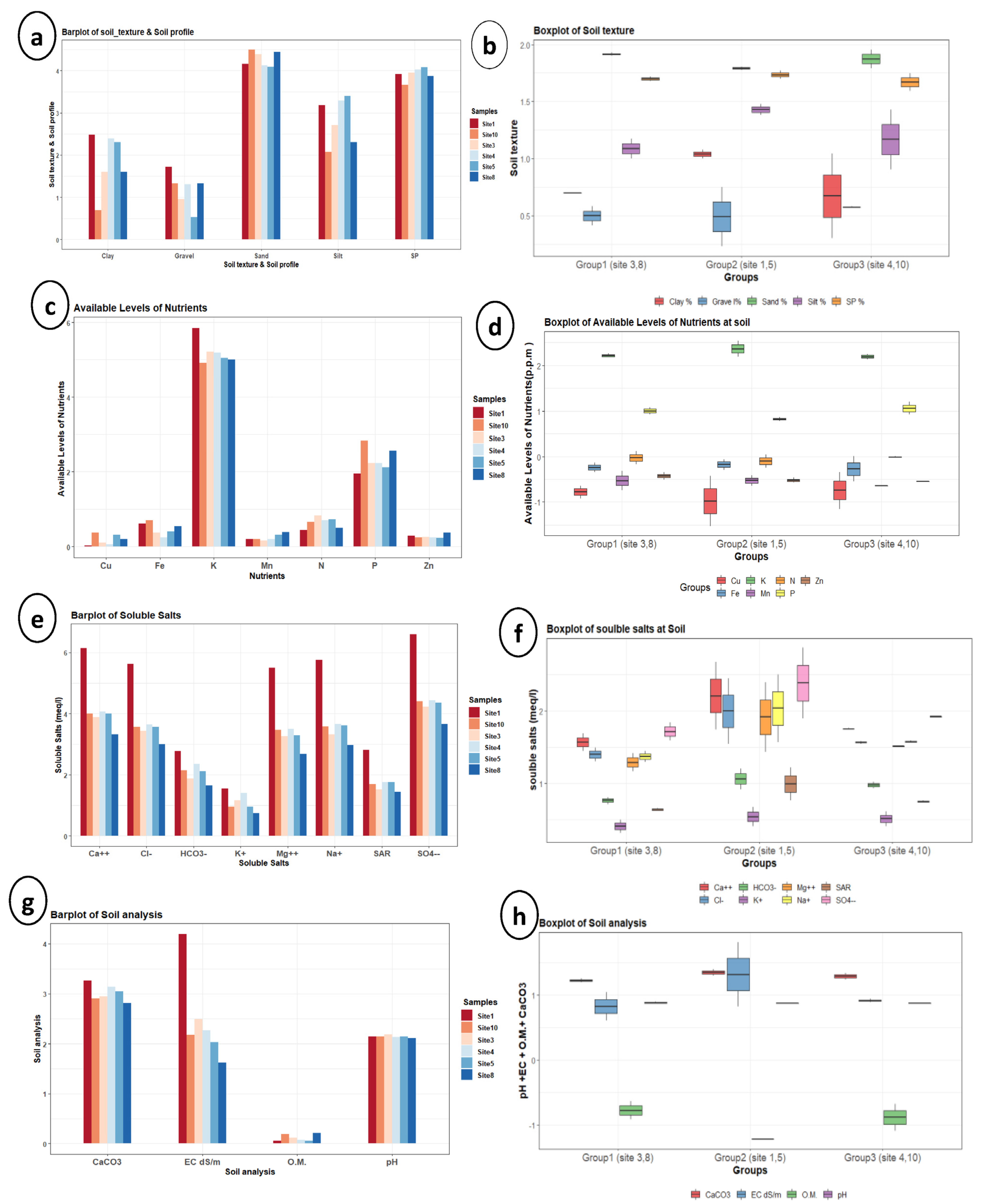
3.4. Soil Analysis
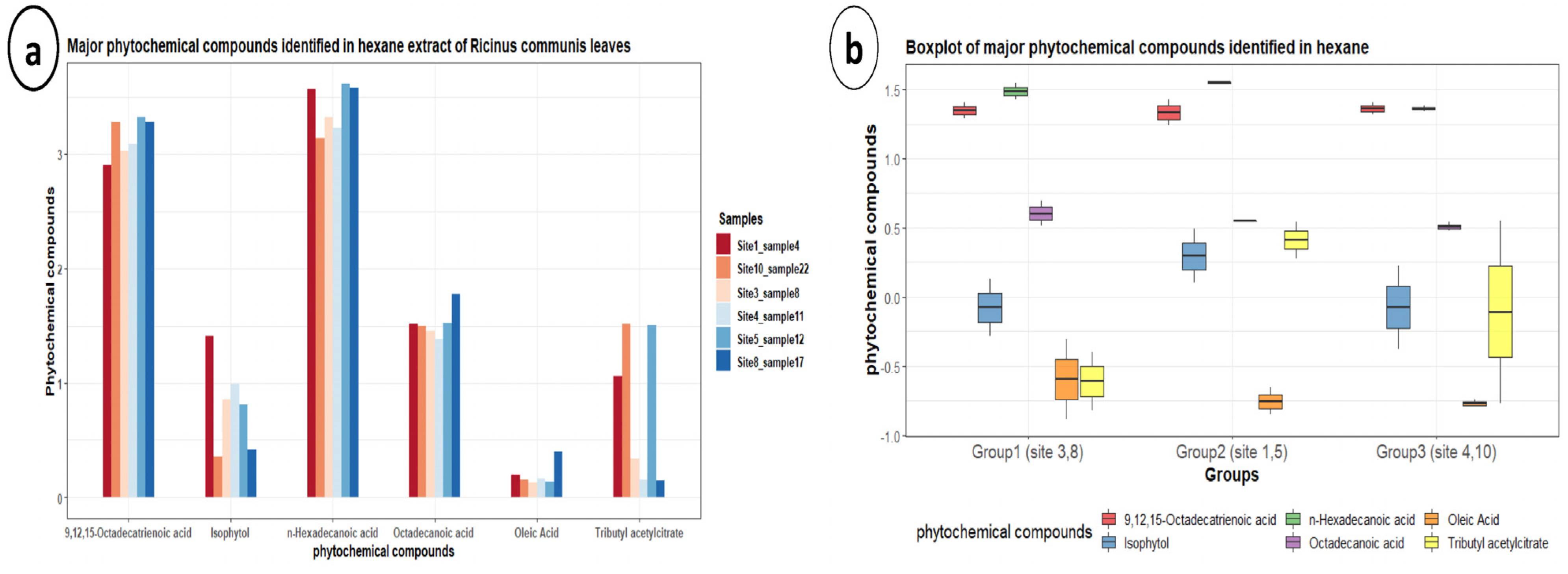
3.5. Gas Chromatography-Mass Spectrometry (GC-MS)
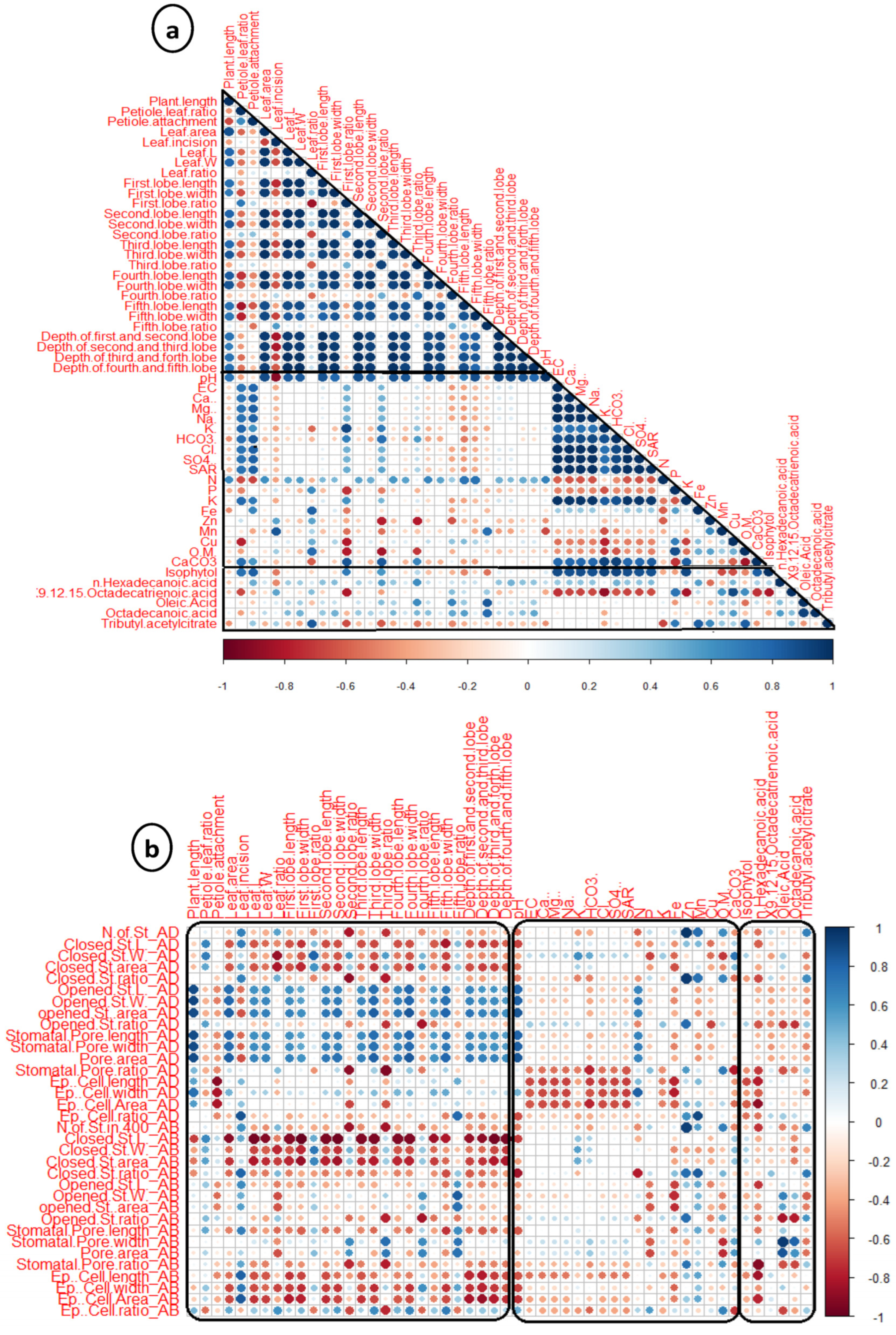
3.6. Correlation Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, H.; Lei, P.; Wang, A.; Liu, S.; Zhao, Y.; Huang, F.; Yu, Z.; Zhu, G.; He, Z.; Tan, D.; et al. Genetic Diversity of Castor Bean (Ricinus communis L.) Revealed by ISSR and RAPD Markers. Agronomy 2021, 11, 457. [Google Scholar] [CrossRef]
- Sahney, M.; Rahi, S.; Kumar, A.; Jaiswal, R. Melissopalynological Studies on Winter Honeys from Allahabad, Uttar Pradesh, India. Palynology 2018, 42, 540–552. [Google Scholar] [CrossRef]
- Yang, S.; Mao, L.; Zheng, Z.; Chen, B.; Li, J. Pollen Atlas for Selected Subfamilies of Euphorbiaceae from Southern China: A Complementary Contribution to Quaternary Pollen Analysis. Palynology 2020, 44, 659–673. [Google Scholar] [CrossRef]
- Kiran, B.R.; Prasad, M.N.V. Ricinus communis L. (Castor Bean), a Potential Multi-Purpose Environmental Crop for Improved and Integrated Phytoremediation. EuroBiotech J. 2017, 1, 101–116. [Google Scholar] [CrossRef]
- Chan, A.P.; Crabtree, J.; Zhao, Q.; Lorenzi, H.; Orvis, J.; Puiu, D.; Melake-Berhan, A.; Jones, K.M.; Redman, J.; Chen, G.; et al. Draft Genome Sequence of the Oilseed Species Ricinus communis. Nat. Biotechnol. 2010, 28, 951–956. [Google Scholar] [CrossRef]
- Perea-Flores, M.J.; Chanona-Pérez, J.J.; Garibay-Febles, V.; Calderón-Dominguez, G.; Terrés-Rojas, E.; Mendoza-Pérez, J.A.; Herrera-Bucio, R. Microscopy Techniques and Image Analysis for Evaluation of Some Chemical and Physical Properties and Morphological Features for Seeds of the Castor Oil Plant (Ricinus communis). Ind. Crops Prod. 2011, 34, 1057–1065. [Google Scholar] [CrossRef]
- Enan, M.; Al-Deeb, M.; Fawzy, N.; Amiri, K. DNA Barcoding of Ricinus communis from Different Geographical Origin by Using Chloroplast Matk and Internal Transcribed Spacers. Am. J. Plant Sci. 2012, 3, 1304–1310. [Google Scholar] [CrossRef]
- Taur, D.J.; Waghmare, M.G.; Bandal, R.S.; Patil, R.Y. Antinociceptive Activity of Ricinus communis L. Leaves. Asian Pac. J. Trop. Biomed. 2011, 1, 139–141. [Google Scholar] [CrossRef]
- Ohishi, K.; Toume, K.; Arai, M.A.; Sadhu, S.K.; Ahmed, F.; Mizoguchi, T.; Itoh, M.; Ishibashi, M. Ricinine: A Pyridone Alkaloid from Ricinus communis That Activates the Wnt Signaling Pathway through Casein Kinase 1α. Bioorg. Med. Chem. 2014, 22, 4597–4601. [Google Scholar] [CrossRef]
- Rakesh, M.R.; Kabra, M.P.; Rajkumar, V.S. Evaluation of Antiulcer Activity of Castor Oil in Rats. Int. J. Res. Ayurveda Pharm. 2011, 2, 1349–1353. [Google Scholar]
- Tunaru, S.; Althoff, T.F.; Nüsing, R.M.; Diener, M.; Offermanns, S. Castor Oil Induces Laxation and Uterus Contraction via Ricinoleic Acid Activating Prostaglandin EP3 Receptors. Proc. Natl. Acad. Sci. USA 2012, 109, 9179–9184. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, J.; Zaib, S.; Farooq, U.; Khan, A.; Bibi, I.; Suleman, S. Antioxidant, Antimicrobial, and Free Radical Scavenging Potential of Aerial Parts of Periploca aphylla and Ricinus communis. Int. Sch. Res. Not. 2012, 2012, 563267. [Google Scholar] [CrossRef]
- Shokeen, P.; Anand, P.; Murali, Y.K.; Tandon, V. Antidiabetic Activity of 50% Ethanolic Extract of Ricinus communis and Its Purified Fractions. Food Chem. Toxicol. 2008, 46, 3458–3466. [Google Scholar] [CrossRef] [PubMed]
- Worbs, S.; Köhler, K.; Pauly, D.; Avondet, M.A.; Schaer, M.; Dorner, M.B.; Dorner, B.G. Ricinus communis Intoxications in Human and Veterinary Medicine-a Summary of Real Cases. Toxins 2011, 3, 1332–1372. [Google Scholar] [CrossRef] [PubMed]
- Abbes, M.; Montana, M.; Curti, C.; Vanelle, P. Ricin Poisoning: A Review on Contamination Source, Diagnosis, Treatment, Prevention and Reporting of Ricin Poisoning. Toxicon 2021, 195, 86–92. [Google Scholar] [CrossRef]
- Sogan, N.; Kapoor, N.; Singh, H.; Kala, S.; Nayak, A.; Nagpal, B. Larvicidal Activity of Ricinus communis Extract against Mosquitoes. J. Vector Borne Dis. 2018, 55, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Hussein, A.O.; Hameed, I.H.; Jasim, H.; Kareem, M.A. Determination of Alkaloid Compounds of Ricinus communis by Using Gas Chromatography-Mass Spectroscopy (GC-MS). J. Med. Plants Res. 2015, 9, 349–359. [Google Scholar] [CrossRef]
- Elkousy, R.H.; Said, Z.N.A.; Abd El-Baseer, M.A.; Abu El wafa, S.A. Antiviral Activity of Castor Oil Plant (Ricinus communis) Leaf Extracts. J. Ethnopharmacol. 2021, 271, 113878. [Google Scholar] [CrossRef]
- Mboyazi, S.N.; Nqotheni, M.I.; Maliehe, T.S.; Shandu, J.S. In Vitro Antibacterial and in Silico Toxicity Properties of Phytocompounds from Ricinus communis Leaf Extract. Pharmacogn. J. 2020, 12, 977–983. [Google Scholar] [CrossRef]
- Osorio-González, C.S.; Gómez-Falcon, N.; Sandoval-Salas, F.; Saini, R.; Brar, S.K.; Ramírez, A.A. Production of Biodiesel from Castor Oil: A Review. Energies 2020, 13, 2467. [Google Scholar] [CrossRef]
- Layek, U.; Manna, S.S.; Karmakar, P. Pollen Foraging Behaviour of Honey Bee (Apis mellifera L.) in Southern West Bengal, India. Palynology 2020, 44, 114–126. [Google Scholar] [CrossRef]
- Allan, G.; Williams, A.; Rabinowicz, P.D.; Chan, A.P.; Ravel, J.; Keim, P. Worldwide Genotyping of Castor Bean Germplasm (Ricinus communis L.) Using AFLPs and SSRs. Genet. Resour. Crop Evol. 2008, 55, 365–378. [Google Scholar] [CrossRef]
- Anjani, K. Purple-Coloured Castor (Ricinus communis L.)-A Rare Multiple Resistant Morphotype. Curr. Sci. 2005, 88, 215–216. [Google Scholar]
- Kallamadi, P.R.; Nadigatla, V.P.R.G.R.; Mulpuri, S. Molecular Diversity in Castor (Ricinus communis L.). Ind. Crops Prod. 2015, 66, 271–281. [Google Scholar] [CrossRef]
- Purseglove, J.W. Castor, Sesame & Safflower By E. A. Weiss London: Leonard Hill Books (1971), pp. 901. Exp. Agric. 1972, 8, 282. [Google Scholar] [CrossRef]
- Chaudhari, B.A.; Patel, M.P.; Patel, J.A.; Makwana, R.R.; Patel, A.M.; Patel, M.K. The Characterization of Castor (Ricinus communis L.) Genotypes for Morphological Traits. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 2482–2492. [Google Scholar] [CrossRef]
- Da Silva, A.R.; Silva, S.A.; Dos Santos, L.A.; De Souza, D.R.; De Melo Araujo, G.; Dantas, J.L.L.; Da Silva Leite, E.; Dantas, A.C.V.L. Characterization and Performance of Castor Bean Lineages and Parents at the UFRB Germplasm Bank. PLoS ONE 2019, 14, e0209335. [Google Scholar] [CrossRef]
- Wahibah, N.N.; Fitmawati; Yahya, V.J.; Perdana, M.A.; Budiono, R. Morphological Variation of Castor Bean (Ricinus communis L.) on Peatland Area in Kepulauan Meranti Riau Indonesia. J. Phys. Conf. Ser. 2020, 1655, 012028. [Google Scholar] [CrossRef]
- Shaheen, A.M. Morphological Variation within Ricinus communis L. Egypt: Fruit, Leaf, Seed and Pollen. Pak. J. Biol. Sci. 2002, 5, 1202–1206. [Google Scholar] [CrossRef]
- Pandey, A.; Agrawal, S.; Patel, A.D.; Pendarkar, D.; Bambhaniya, J.D. Evergreen Conventional and Modern Microscopic Methods for Solving Various Cases of Forensic Botany by Morphological and Histological Study. Pharmacogn. J. 2019, 11, 171–176. [Google Scholar] [CrossRef]
- CSIR. The Wealth of India: A Dictionary of Indian Raw Materials and Industrial Products (Industrial Products—Part I); Council of Scientific & Industrial Research: New Delhi, India, 1972.
- Fakhr, M.A.; Mazrou, Y.S.A.; Ellmouni, F.Y.; Elsaied, A.; Elhady, M.; Elkelish, A.; Nour, I.H. Investigating the Phenotypic Plasticity of the Invasive Weed Trianthema portulacastrum L. Plants 2022, 11, 77. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.; Chen, Y.; Ponce, G.; Acevedo, F.E.; Anderson, C.T.; Ali, J.G.; Felton, G.W.; Lynch, J.P. Stomata-Mediated Interactions between Plants, Herbivores, and the Environment. Trends Plant Sci. 2022, 27, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.F.; Cardoso, D.; de Queiroz, L.P. An Updated Plant Checklist of the Brazilian Caatinga Seasonally Dry Forests and Woodlands Reveals High Species Richness and Endemism. J. Arid Environ. 2020, 174, 104079. [Google Scholar] [CrossRef]
- Duruibe, J.O.; Ogwuegbu, M.O.C.; Egwurugwu, J.N. Heavy Metal Pollution and Human Biotoxic Effects. Int. J. Phys. Sci. 2007, 2, 112–118. [Google Scholar] [CrossRef]
- Nazzal, Y.; Rosen, M.A.; Al-Rawabdeh, A.M. Assessment of Metal Pollution in Urban Road Dusts from Selected Highways of the Greater Toronto Area in Canada. Environ. Monit. Assess. 2013, 185, 1847–1858. [Google Scholar] [CrossRef] [PubMed]
- Sameena, P.; Puthur, J.T. Heavy Metal Phytoremediation by Bioenergy Plants and Associated Tolerance Mechanisms. Soil Sediment Contam. 2021, 30, 253–274. [Google Scholar] [CrossRef]
- Huang, J.; Guo, S.; Zeng, G.; Li, F.; Gu, Y.; Shi, Y.; Shi, L.; Liu, W.; Peng, S. A New Exploration of Health Risk Assessment Quantification from Sources of Soil Heavy Metals under Different Land Use. Environ. Pollut. 2018, 243, 49–58. [Google Scholar] [CrossRef]
- Li, C.; Zhou, K.; Qin, W.; Tian, C.; Qi, M.; Yan, X.; Han, W. A Review on Heavy Metals Contamination in Soil: Effects, Sources, and Remediation Techniques. Soil Sediment Contam. 2019, 28, 380–394. [Google Scholar] [CrossRef]
- Farooq, M.A.; Ali, S.; Hameed, A.; Bharwana, S.A.; Rizwan, M.; Ishaque, W.; Farid, M.; Mahmood, K.; Iqbal, Z. Cadmium Stress in Cotton Seedlings: Physiological, Photosynthesis and Oxidative Damages Alleviated by Glycinebetaine. South Afr. J. Bot. 2016, 104, 61–68. [Google Scholar] [CrossRef]
- Siregar, A.S.; Sulistyo, I.; Prayogo, N.A. Heavy Metal Contamination in Water, Sediments and Planiliza Subviridis Tissue in the Donan River, Indonesia. J. Water L. Dev. 2020, 45, 157–164. [Google Scholar] [CrossRef]
- Tyagi, K.; Sharma, S.; Kumar, S.; Ayub, S. Cytological, Morphological and Anatomical Studies of Ricinus communis Linn. Grown under the Influence of Industrial Effluent—A Comparative Study. J. Pharm. Res. 2013, 7, 454–458. [Google Scholar] [CrossRef]
- Bauddh, K.; Singh, K.; Singh, B.; Singh, R.P. Ricinus communis: A Robust Plant for Bio-Energy and Phytoremediation of Toxic Metals from Contaminated Soil. Ecol. Eng. 2015, 84, 640–652. [Google Scholar] [CrossRef]
- Kammerbauer, J.; Dick, T. Monitoring of Urban Traffic Emissions Using Some Physiological Indicators in Ricinus communis L. Plants. Arch. Environ. Contam. Toxicol. 2000, 39, 161–166. [Google Scholar] [CrossRef]
- Boda, R.K.; Majeti, N.V.P.; Suthari, S. Ricinus communis L. (Castor Bean) as a Potential Candidate for Revegetating Industrial Waste Contaminated Sites in Peri-Urban Greater Hyderabad: Remarks on Seed Oil. Environ. Sci. Pollut. Res. 2017, 24, 19955–19964. [Google Scholar] [CrossRef] [PubMed]
- Zarai, Z.; Chobba, I.B.; Mansour, R.B.; Békir, A.; Gharsallah, N.; Kadri, A. Essential Oil of the Leaves of Ricinus communis L.: In Vitro Cytotoxicity and Antimicrobial Properties. Lipids Health Dis. 2012, 11, 102. [Google Scholar] [CrossRef] [PubMed]
- Warra, A.A. Physico-Chemical and GC/MS Analysis of Castor Bean (Ricinus communis L.) Seed Oil. Chem. Mater. Res. 2015, 7, 56–60. [Google Scholar]
- Hammad, H.H.; El-Kateb, H.; Bahnasy, M.I. Growth, Yield and Fatty Acids Composition of Castor Bean (Ricinus communis L.) Cultivars. Middle East J. Agric. Res. 2019, 8, 638–653. [Google Scholar]
- Firn, J.; McGree, J.M.; Harvey, E.; Flores-Moreno, H.; Schütz, M.; Buckley, Y.M.; Borer, E.T.; Seabloom, E.W.; La Pierre, K.J.; MacDougall, A.M.; et al. Leaf Nutrients, Not Specific Leaf Area, Are Consistent Indicators of Elevated Nutrient Inputs. Nat. Ecol. Evol. 2019, 3, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Perdomo, F.A.; Acosta-Osorio, A.A.; Herrera, G.; Vasco-Leal, J.F.; Mosquera-Artamonov, J.D.; Millan-Malo, B.; Rodriguez-Garcia, M.E. Physicochemical Characterization of Seven Mexican Ricinus communis L. Seeds & Oil Contents. Biomass Bioenergy 2013, 48, 17–24. [Google Scholar]
- Hussein, H.M.; Ubaid, J.M.; Hameed, I.H. Inscticidal Activity of Methanolic Seeds Extract of Ricinus communis on Adult of Callosobruchus maculatus (Coleopteran: Brauchidae) and Analysis of Its Phytochemical Composition. Int. J. Pharmacogn. Phytochem. Res. 2016, 8, 1385–1397. [Google Scholar]
- Salimon, J.; Noor, D.A.M.; Nazrizawati, A.; Noraishah, A. Fatty Acid Composition and Physicochemical Properties of Malaysian Castor Bean Ricinus communis L. Seed Oil. Sains Malays. 2010, 39, 761–764. [Google Scholar]
- Nangbes, J.G.; Nvau, J.B.; Buba, W.M.; Zukdimma, A.N. Extraction and Characterization of Castor (Ricinus communis) Seed Oil. Int. J. Eng. Sci. 2013, 2, 105–109. [Google Scholar]
- Yusuf, A.K.; Mamza, P.A.P.; Ahmed, A.S.; Agunwa, U. Extraction and Characterization of Castor Seed Oil from Wild Ricinus communis Linn. Int. J. Sci. Environ. Technol. 2015, 4, 1392–1404. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Singh, G. Plant Systematics: An Integrated Approach; CRC Press: Boca Raton, FL, USA, 2019; ISBN 1000576779. [Google Scholar]
- Ellis, B.; Daly, D.C.; Hickey, L.J.; Johnson, K.R.; Mitchell, J.D.; Wilf, P.; Wing, S.L. Manual of Leaf Architecture; Ithaca: New York, NY, USA, 2009; ISBN 9780801475184. [Google Scholar]
- Ash, A. Manual of Leaf Architecture: Morphological Description and Categorization of Dicotyledonous and Net-Veined Monocotyledonous Angiosperms; Smithsonian Institution: Washington, DC, USA, 1999. [Google Scholar]
- Barthlott, W. Epidermal and Seed Surface Characters of Plants: Systematic Applicability and Some Evolutionary Aspects. Nord. J. Bot. 1981, 1, 345–355. [Google Scholar] [CrossRef]
- Piper, C.S. Soil and Plant Analysis; Scientific Publishers: Jodhpur, India, 2019; ISBN 9386237539. [Google Scholar]
- Walkley, A.J.; Black, I.A. Estimation of Soil Organic Carbon by the Chromic Acid Titration Method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Pansu, M.; Gautheyrou, J. Phosphorus; Springer: Berlin/Heidelberg, Germany, 2006; ISBN 978-3-540-31211-6. [Google Scholar]
- Olsen, S.R.; Sommers, L.E. Phosphorus. Agronomy Monographs. In Methods of Soil Analysis; Academic Press: Madison, WI, USA, 1982; pp. 403–430. ISBN 9780891189770. [Google Scholar]
- Hanway, J.J.; Heidal, H. Soil Analysis Methods as Used in the Iowa State College Soil Testing Laboratory. Iowa State Coll. Agric. Bull. 1952, 57, 1–31. [Google Scholar]
- Tabassum, S.; Amin, F.; Erum, S.; Javed, H.; Kazmi, F.; Nisar, M.F.; Ullah, I.; Murtaza, I.; Ashraf, M. Effect of Hexane and Ethanol Extracts of Ten Basil Genotypes on the Growth of Selected Bacterial Strains. Int. J. Agric. Biol. 2016, 18, 735–740. [Google Scholar] [CrossRef]
- Sbihi, H.M.; Nehdi, I.A.; Mokbli, S.; Romdhani-Younes, M.; Al-Resayes, S.I. Hexane and Ethanol Extracted Seed Oils and Leaf Essential Compositions from Two Castor Plant (Ricinus communis L.) Varieties. Ind. Crops Prod. 2018, 122, 174–181. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2013. Available online: https://www.gbif.org/tool/81287/r-a-language-and-environment-for-statistical-computing (accessed on 30 January 2023).
- Wickham, H. Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016; ISBN 331924275X. [Google Scholar]
- Kolde, R. Pheatmap: Pretty Heatmaps, Version 1.0.8; R Package; R Foundation: Vienna, Austria, 2012.
- Viscosi, V.; Cardini, A. Leaf Morphology, Taxonomy and Geometric Morphometrics: A Simplified Protocol for Beginners. PLoS ONE 2011, 6, e25630. [Google Scholar] [CrossRef] [PubMed]
- Kassambara, A.; Mundt, F. Factoextr: Extract and Visualize the Results of Multivariate Data Analyses. 2020. Available online: https://cran.r-project.org/web/packages/factoextra/index.html (accessed on 30 January 2023).
- Soetewey, A. Correlation Coefficient and Correlation Test in R. Available online: https://www.statsandr.com/blog/correlationcoefficient-and-correlation-test-in-r/ (accessed on 20 February 2022).
- Murphy, M.R.C.; Jordan, G.J.; Brodribb, T.J. Differential Leaf Expansion Can Enable Hydraulic Acclimation to Sun and Shade. Plant Cell Environ. 2012, 35, 1407–1418. [Google Scholar] [CrossRef] [PubMed]
- Xiong, D.; Wang, D.; Liu, X.; Peng, S.; Huang, J.; Li, Y. Leaf Density Explains Variation in Leaf Mass per Area in Rice between Cultivars and Nitrogen Treatments. Ann. Bot. 2016, 117, 963–971. [Google Scholar] [CrossRef]
- James, J.J.; Tiller, R.L.; Richards, J.H. Multiple Resources Limit Plant Growth and Function in a Saline-Alkaline Desert Community. J. Ecol. 2005, 93, 113–126. [Google Scholar] [CrossRef]
- Meier, I.C.; Leuschner, C. Leaf Size and Leaf Area Index in Fagus sylvatica Forests: Competing Effects of Precipitation, Temperature, and Nitrogen Availability. Ecosystems 2008, 11, 655–669. [Google Scholar] [CrossRef]
- Liu, F.; Yang, W.; Wang, Z.; Xu, Z.; Liu, H.; Zhang, M.; Liu, Y.; An, S.; Sun, S. Plant Size Effects on the Relationships among Specific Leaf Area, Leaf Nutrient Content, and Photosynthetic Capacity in Tropical woody Species. Acta Oecologica 2010, 36, 149–159. [Google Scholar] [CrossRef]
- Bano, A.; Fatima, M. Salt Tolerance in Zea mays (L). Following Inoculation with Rhizobium and Pseudomonas. Biol. Fertil. Soils 2009, 45, 405–413. [Google Scholar] [CrossRef]
- Sathee, L.; Jain, V. Interaction of Elevated CO2 and Form of Nitrogen Nutrition Alters Leaf Abaxial and Adaxial Epidermal and Stomatal Anatomy of Wheat Seedlings. Protoplasma 2022, 3, 703–716. [Google Scholar] [CrossRef]
- Bravo, S.; Amorós, J.A.; Pérez-De-Los-Reyes, C.; García, F.J.; Moreno, M.M.; Sánchez-Ormeño, M.; Higueras, P. Influence of the Soil PH in the Uptake and Bioaccumulation of Heavy Metals (Fe, Zn, Cu, Pb and Mn) and Other Elements (Ca, K, Al, Sr and Ba) in Vine Leaves, Castilla-La Mancha (Spain). J. Geochem. Explor. 2017, 174, 79–83. [Google Scholar] [CrossRef]
- Mohamed, I.A.A.; Shalby, N.; MA El-Badri, A.; Saleem, M.H.; Khan, M.N.; Nawaz, M.A.; Qin, M.; Agami, R.A.; Kuai, J.; Wang, B. Stomata and Xylem Vessels Traits Improved by Melatonin Application Contribute to Enhancing Salt Tolerance and Fatty Acid Composition of Brassica napus L. Plants. Agronomy 2020, 10, 1186. [Google Scholar] [CrossRef]
- Galal, T.M.; Essa, B.; Al-Yasi, H. Heavy Metals Uptake and Its Impact on the Growth Dynamics of the Riparian Shrub Ricinus communis L. along Egyptian Heterogenic Habitats. Environ. Sci. Pollut. Res. 2021, 28, 37158–37171. [Google Scholar] [CrossRef]
- Azmat, R.; Saleem, A.; Ahmed, W.; Qayyum, A.; El-Serehy, H.A.; Ali, S. The Investigation of the Impact of Toxicity of Metals on Oxygen-Evolving Complex in Spinacia oleracea. Antioxidants 2022, 11, 1802. [Google Scholar] [CrossRef] [PubMed]
- Abdel Latef, A.A.H.; Zaid, A.; Abo-Baker, A.B.A.E.; Salem, W.; Abu Alhmad, M.F. Mitigation of Copper Stress in Maize by Inoculation with Paenibacillus polymyxa and Bacillus circulans. Plants 2020, 9, 1513. [Google Scholar] [CrossRef] [PubMed]
- Yruela, I. Copper in Plants. Braz. J. Plant Physiol. 2005, 17, 145–156. [Google Scholar] [CrossRef]
- delos Angeles, M.D.; Serino, C.B.M.; Granada, K.B. Leaf Morpho-Anatomical Responses of Mangifera indica L. and Ficus benjamina L. to Air Pollution in Selected Areas of Cebu City, Philippines. J. Nat. Stud. 2018, 17, 44–53. [Google Scholar]
- Morsy, A.A.; Youssef, A.M.; Mosallam, H.A.M.; Hashem, A.M. Assessment of Selected Species along Al-Alamein-Alexandria International Desert Road, Egypt. J. Appl. Sci. Res. 2008, 4, 1276–1284. [Google Scholar]
- Marzouk, R.I.; El-darier, S.M.; Nour, I.H.; Kamal, S.A. Numerical Taxonomic Study of Marrubium L. (Lamiaceae) in Egypt. Catrina 2015, 13, 25–35. [Google Scholar]
- Halmy, M.W.A.; Fawzy, M.; Ahmed, D.A.; Saeed, N.M.; Awad, M.A. Monitoring and Predicting the Potential Distribution of Alien Plant Species in Arid Ecosystem Using Remotely-Sensed Data. Remote Sens. Appl. Soc. Environ. 2019, 13, 69–84. [Google Scholar] [CrossRef]
- Shankar, V.G.; Venkata Ramana Rao, P.; Reddy, A.V. Inheritance of Certain Morphological Characters and Fusarium Wilt Resistance in Castor, Ricinus communis L. Sabrao J. Breed. Genet. 2010, 42, 57–64. [Google Scholar]
- Santha, E.B.M.; Prasad, L.; Yadav, P.; Bee, H. Defense Responses to Fusarium oxysporum f. sp. ricini Infection in Castor (Ricinus communis L.) Cultivars. Indian Phytopathol. 2019, 72, 647–656. [Google Scholar] [CrossRef]
- Toscano, S.; Branca, F.; Romano, D.; Ferrante, A. An Evaluation of Different Parameters to Screen Ornamental Shrubs for Salt Spray Tolerance. Biology 2020, 9, 250. [Google Scholar] [CrossRef]
- Lacerda, C.F.; Assis Júnior, J.O.; Lemos Filho, L.C.A.; de Oliveira, T.S.; Guimarães, F.V.A.; Gomes-Filho, E.; Prisco, J.T.; Bezerra, M.A. Morpho-Physiological Responses of Cowpea Leaves to Salt Stress. Braz. J. Plant Physiol. 2006, 18, 455–465. [Google Scholar] [CrossRef]
- Matesanz, S.; Horgan-Kobelski, T.; Sultan, S.E. Phenotypic Plasticity and Population Differentiation in an Ongoing Species Invasion. PLoS ONE 2012, 7, e44955. [Google Scholar] [CrossRef]
- Cookson, S.J.; Radziejwoski, A.; Granier, C. Cell and Leaf Size Plasticity in Arabidopsis: What Is the Role of Endoreduplication? Plant Cell Environ. 2006, 29, 1273–1283. [Google Scholar] [CrossRef] [PubMed]
- Barthlott, W.; Neinhuis, C.; Cutler, D.; Ditsch, F.; Meusel, I.; Theisen, I.; Wilhelmi, H. Classification and Terminology of Plant Epicuticular Waxes. Bot. J. Linn. Soc. 1998, 126, 237–260. [Google Scholar] [CrossRef]
- Reddy, P. Influence of Nutrient Rich Organic Wastes in Wastelands Reclamation; GRIN Verlag: München, Germany, 2014. [Google Scholar]
- Öncel, I.; Keleş, Y.; Üstün, A.S. Interactive Effects of Temperature and Heavy Metal Stress on the Growth and Some Biochemical Compounds in Wheat Seedlings. Environ. Pollut. 2000, 107, 315–320. [Google Scholar] [CrossRef]
- Jeschke, W.; Wolf, O. Effect of NaCI Salinity on Growth, Development, Ion Distribution, and Ion Translocation in Castor Bean (Ricinus communis L.). J. Plant Physiol. 1988, 132, 45–53. [Google Scholar] [CrossRef]
- Papazoglou, E.G.; Alexopoulou, E.; Papadopoulos, G.K.; Economou-Antonaka, G. Tolerance to Drought and Water Stress Resistance Mechanism of Castor Bean. Agronomy 2020, 10, 1580. [Google Scholar] [CrossRef]
- Zhou, H.; Zhou, G.; He, Q.; Zhou, L.; Ji, Y.; Zhou, M. Environmental Explanation of Maize Specific Leaf Area under Varying Water Stress Regimes. Environ. Exp. Bot. 2020, 171, 103932. [Google Scholar] [CrossRef]
- Elliott-kingston, C.; Haworth, M.; Yearsley, J.M.; Batke, S.P.; Lawson, T.; Mcelwain, J.C.; Elliott-kingston, C. Does Size Matter? Atmospheric CO2 May Be a Stronger Driver of Stomatal Closing Rate than Stomatal Size in Taxa That Diversified under Low CO2. Front. Plant Sci. 2016, 7, 1253. [Google Scholar] [CrossRef] [PubMed]
- Ullah, S.; Khan, M.N.; Lodhi, S.S.; Ahmed, I.; Tayyab, M.; Mehmood, T.; Din, I.U.; Khan, M.; Sohail, Q.; Akram, M. Targeted Metabolomics Reveals Fatty Acid Abundance Adjustments as Playing a Crucial Role in Drought-Stress Response and Post-Drought Recovery in Wheat. Front. Genet. 2022, 13, 972696. [Google Scholar] [CrossRef] [PubMed]
- Yaghoubian, I.; Antar, M.; Ghassemi, S.; Modarres-Sanavy, S.A.M.; Smith, D.L. The Effects of Hydro-Priming and Colonization with Piriformospora indica and Azotobacter chroococcum on Physio-Biochemical Traits, Flavonolignans and Fatty Acids Composition of Milk Thistle (Silybum marianum) under Saline Conditions. Plants 2022, 11, 1281. [Google Scholar] [CrossRef] [PubMed]
- Aparna, V.; Dileep, K.V.; Mandal, P.K.; Karthe, P.; Sadasivan, C.; Haridas, M. Anti-Inflammatory Property of n-Hexadecanoic Acid: Structural Evidence and Kinetic Assessment. Chem. Biol. Drug Des. 2012, 80, 434–439. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Sample No | Coordination | Location |
|---|---|---|---|
| 1 | 1–4 | 31°02′40.3″ N 29°42′13.9″ E | Mehwar Al-Ta’ameir, about 700 m from kilo 26 wastewater treatment plant, and 26 km from West Alexandria. |
| 2 | 5–7 | 31°00′00.0″ N 29°36′35.2″ E | 36 km from Alexandria–Matrouh International Coastal Road, and 200 m from Amoun Resort, Sidi Kirayr. |
| 3 | 8–9 | 30°59′19.3″ N 29°35′27.2″ E | 39 km from Alexandria–Matrouh International Coastal Road, Sidi Kirayr. |
| 4 | 10–11 | 30°49′19.5″ N 29°11′47.8″ E | Omayed Biosphere Reserve, Alexandria Desert Road. |
| 5 | 12–13 | 30°49′16.4″ N 29°11′49.9″ E | 1 km from East Omayed biosphere reserve, Alexandria Desert Road. |
| 6 | 14–15 | 30°52′07.4″ N 29°20′20.3″ E | International Coastal Road, 5 km west of El Hammam Central Hospital, El Hammam. |
| 7 | 16 | 30°52′50.7″ N 29°22′07.9″ E | International Coastal Road, 2 km west of El Marwa Resort, El Hammam, Egypt. |
| 8 | 17–19 | 31°00′43.6″ N 29°38′04.7″ E | 175 m from South Sidi Kirayr Bridge. |
| 9 | 20–21 | 30°56′57.6″ N 29°41′27.5″ E | 3 km from Borg-El Arab International Airport, Borg El-Arab. |
| 10 | 22–23 | 30°56′50.7″ N 29°50′31.9″ E | Cairo–Alexandria Desert Road, 7 km south of Amaria General Hospital, second Al Amaria. |
| 11 | 24–25 | 30°56′14.3″ N 29°50′57.9″ E | Cairo–Alexandria Desert Road, 11 km south of Amaria General Hospital, second Al Amaria. |
| Variable | Mean ± StDev | SE Mean | Min. | Q1 | Median | Q3 | Max. | IQR |
|---|---|---|---|---|---|---|---|---|
| Plant length (m) | 2.144 ± 0.725 | 0.145 | 1.02 | 1.57 | 2.03 | 2.7 | 3.72 | 1.13 |
| Petiole length/leaf length ratio | 0.9756 ± 0.2785 | 0.0557 | 0.49 | 0.755 | 1.01 | 1.155 | 1.54 | 0.4 |
| Petiole attachment (%) | 18.302 ± 2.838 | 0.568 | 11.76 | 16.31 | 17.65 | 20.805 | 23 | 4.495 |
| Leaf area (cm2) | 178.7 ± 90.9 | 18.2 | 52.9 | 112.1 | 151.8 | 251 | 352.9 | 138.9 |
| Leaf incision (%) | 63.796 ± 4.454 | 0.891 | 54.82 | 61.525 | 63.77 | 65.755 | 74.4 | 4.23 |
| Leaf length (cm) | 19.351 ± 4.722 | 0.944 | 11.6 | 15.805 | 18.42 | 23.38 | 27.49 | 7.575 |
| Leaf width (cm) | 20.57 ± 5.33 | 1.07 | 12.3 | 16.82 | 19.62 | 25.38 | 30.52 | 8.55 |
| Leaf length/width ratio | 0.9444 ± 0.03318 | 0.00664 | 0.88 | 0.92 | 0.94 | 0.97 | 1 | 0.05 |
| First lobe length (cm) | 13.51 ± 3.366 | 0.673 | 8.22 | 10.775 | 13.63 | 16.495 | 20.06 | 5.72 |
| First lobe width (cm) | 4.579 ± 1.217 | 0.243 | 2.17 | 3.615 | 4.53 | 5.515 | 7 | 1.9 |
| First lobe length/width ratio | 3.0224 ± 0.3283 | 0.0657 | 2.43 | 2.785 | 3 | 3.205 | 3.97 | 0.42 |
| Second lobe length (cm) | 12.505 ± 3.161 | 0.632 | 7.55 | 10.09 | 11.83 | 15.04 | 18.84 | 4.95 |
| Second lobe width (cm) | 3.864 ± 1.019 | 0.204 | 1.97 | 3.26 | 3.69 | 4.62 | 5.51 | 1.36 |
| Second lobe length/width ratio | 3.3036 ± 0.2736 | 0.0547 | 2.75 | 3.15 | 3.3 | 3.46 | 4.15 | 0.31 |
| Third lobe length (cm) | 10.855 ± 2.924 | 0.585 | 6.64 | 8.765 | 10.03 | 13.445 | 16.62 | 4.68 |
| Third lobe width (cm) | 3.075 ± 0.893 | 0.179 | 1.72 | 2.52 | 2.92 | 3.865 | 4.72 | 1.345 |
| Third lobe length/width ratio | 3.6092 ± 0.29 | 0.058 | 2.94 | 3.39 | 3.66 | 3.825 | 4.17 | 0.435 |
| Fourth lobe length (cm) | 8.916 ± 2.588 | 0.518 | 4.93 | 7.14 | 8.34 | 11.095 | 13.69 | 3.955 |
| Fourth lobe width (cm) | 2.39 ± 0.757 | 0.151 | 1.25 | 1.93 | 2.26 | 2.77 | 3.78 | 0.84 |
| Fourth lobe length/width ratio | 3.8452 ± 0.378 | 0.0756 | 3.08 | 3.54 | 3.79 | 4.16 | 4.59 | 0.62 |
| Fifth lobe length (cm) | 6.933 ± 2.187 | 0.437 | 3.5 | 5.045 | 6.46 | 9.135 | 10.21 | 4.09 |
| Fifth lobe width (cm) | 1.739 ± 0.583 | 0.117 | 0.88 | 1.225 | 1.72 | 2.195 | 2.8 | 0.97 |
| Fifth lobe length/width ratio | 4.103 ± 0.508 | 0.102 | 3.29 | 3.725 | 4.06 | 4.44 | 5.33 | 0.715 |
| Depth between first and second lobe (cm) | 4.948 ± 1.807 | 0.207 | 1.5 | 3.682 | 4.617 | 6.029 | 9.19 | 2.348 |
| Depth between second and third lobe (cm) | 4.68 ± 1.69 | 0.194 | 1.559 | 3.524 | 4.435 | 5.472 | 8.902 | 1.948 |
| Depth between third and fourth lobe (cm) | 3.922 ± 1.489 | 0.171 | 1.313 | 2.697 | 3.643 | 4.797 | 7.609 | 2.099 |
| Depth between fourth and fifth lobe (cm) | 3.028 ± 1.182 | 0.136 | 0.98 | 2.043 | 2.817 | 3.848 | 6.204 | 1.805 |
| Dimension 1 | r | Dimension 2 | r |
|---|---|---|---|
| Leaf area (cm2) | 0.987 | Third lobe length/width ratio | 0.818 |
| Depth of third and fourth lobe (cm) | 0.984 | Second lobe length/width ratio | 0.793 |
| Second lobe width (cm) | 0.980 | Fourth lobe length/width ratio | 0.691 |
| Third lobe width (cm) | 0.980 | First lobe shape | 0.679 |
| Fourth lobe length (cm) | 0.980 | First lobe length/width ratio | 0.651 |
| Third lobe length (cm) | 0.978 | Second lobe shape | 0.626 |
| Depth of fourth and fifth lobe (cm) | 0.9745 | Third lobe shape | 0.555 |
| Leaf width (cm) | 0.973 | Fifth lobe length/width ratio | 0.463 |
| Leaf length (cm) | 0.970 | Petiole attachment (%) | −0.613 |
| First lobe length (cm) | 0.970 | ||
| Second lobe length (cm) | 0.970 | ||
| Fourth lobe width (cm) | 0.967 | ||
| Depth of second and third lobe (cm) | 0.966 | ||
| Depth of first and second lobe (cm) | 0.953 | ||
| Fifth lobe length (cm) | 0.948 | ||
| First lobe width (cm) | 0.932 | ||
| Fifth lobe width (cm) | 0.887 | ||
| Blade class | 0.878 | ||
| Leaf incision (%) | −0.568 | ||
| Leaf length/width ratio | −0.425 | ||
| Eigenvalue | 17.68 | Eigenvalue | 4.640 |
| Variance % | 53.59 | Variance % | 14.06 |
| Cumulative variance % | 53.59 | Cumulative variance % | 67.66 |
| Variable | Group 1 | Group 2 | Group 3 |
|---|---|---|---|
| Plant length (m) | 2.207 ± 0.721 a | 1.931 ± 0.72 a | 2.403 ± 0.764 a |
| Petiole length/leaf length ratio | 0.91 ± 0.2776 a | 0.962 ± 0.262 a | 1.097 ± 0.316 a |
| Petiole attachment (%) | 17.824 ± 2.417 a | 19.165 ± 2.671 a | 17.58 ± 3.74 a |
| Leaf area (cm2) | 283.7 ± 50.2 a | 145.57 ± 17.9 b | 76.61 ± 22.31 c |
| Leaf incision (%) | 60.75 ± 3.64 b | 64.74 ± 4.19 ab | 66.8 ± 3.62 a |
| Leaf length (cm) | 24.523 ± 1.79 a | 18.393 ± 1.409 b | 13.19 ± 1.676 c |
| Leaf width (cm) | 26.493 ± 2.318 a | 19.281 ± 1.459 b | 13.813 ± 1.622 c |
| Leaf length/width ratio | 0.92333 ± 0.01936 a | 0.958 ± 0.02974 a | 0.9533 ± 0.0427 a |
| First lobe length (cm) | 17.168 ± 1.489 a | 12.832 ± 1.016 b | 9.152 ± 1.081 b |
| First lobe width (cm) | 5.801 ± 0.575 a | 4.412 ± 0.549 b | 3.023 ± 0.624 c |
| First lobe length/width ratio | 2.991 ± 0.303 a | 2.968 ± 0.2523 a | 3.16 ± 0.475 a |
| Second lobe length (cm) | 15.961 ± 1.587 a | 11.839 ± 0.699 b | 8.432 ± 0.861 c |
| Second lobe width (cm) | 4.937 ± 0.496 a | 3.672 ± 0.319 b | 2.577 ± 0.526 c |
| Second lobe length/width ratio | 3.2778 ± 0.2954 a | 3.275 ± 0.1203 a | 3.39 ± 0.426 a |
| Third lobe length (cm) | 14.174 ± 1.402 a | 10.009 ± 0.645 b | 7.285 ± 0.769 c |
| Third lobe width (cm) | 4.047 ± 0.541 a | 2.832 ± 0.243 b | 2.023 ± 0.334 c |
| Third lobe length/width ratio | 3.57 ± 0.358 a | 3.607 ± 0.2132 a | 3.672 ± 0.33 a |
| Fourth lobe length (cm) | 11.871 ± 1.122 a | 8.121 ± 0.689 b | 5.81 ± 0.845 c |
| Fourth lobe width (cm) | 3.198 ± 0.517 a | 2.183 ± 0.1863 b | 1.523 ± 0.315 c |
| Fourth lobe length/width ratio | 3.801 ± 0.403 a | 3.812 ± 0.3083 a | 3.967 ± 0.48 a |
| Fifth lobe length (cm) | 9.416 ± 0.586 a | 6.249 ± 1.029 b | 4.35 ± 0.681 c |
| Fifth lobe width (cm) | 2.303 ± 0.395 a | 1.641 ± 0.338 b | 1.055 ± 0.142 c |
| Fifth lobe length/width ratio | 4.252 ± 0.63 a | 3.923 ± 0.415 a | 4.178 ± 0.426 a |
| Depth between first and second lobe (cm) | 6.744 ± 0.866 a | 4.494 ± 0.488 b | 3.052 ± 0.494 c |
| Depth between second and third lobe (cm) | 6.322 ± 0.772 a | 4.3113 ± 0.3129 b | 2.88 ± 0.459 c |
| Depth between third and fourth lobe (cm) | 5.403 ± 0.567 a | 3.5332 ± 0.2429 b | 2.373 ± 0.394 c |
| Depth between fourth and fifth lobe (cm) | 4.2 ± 0.512 a | 2.7431 ± 0.1295 b | 1.766 ± 0.303 c |
| Characters | Group 1 (Samples 8 and 17) | Group 2 (Samples 4 and 12) | Group 3 (Samples 11 and 22) | |||
|---|---|---|---|---|---|---|
| Abaxial | Adaxial | Abaxial | Adaxial | Abaxial | Adaxial | |
| Number of stomata | 14.67 ± 5.77 a | 13.75 ± 1.258 a | 16.5 ± 0.707 a | 15 ± 1.41 a | 25 ± 13 a | 17.67 ± 7.23 a |
| Closed stomata length (µm) | 9.095 ± 1.337 bc | 8.567 ± 1.934 c | 13.5 ± 1.238 ab | 8.28 ± 3.21 c | 16.68 ± 4.9 a | 12.22 ± 1.315 abc |
| Closed stomata width (µm) | 4.719 ± 0.395 a | 3.889 ± 1.345 a | 5.525 ± 0.602 a | 3.737 ± 1.469 a | 6.37 ± 3.1 a | 4.478 ± 1.261 a |
| Closed stomata length/width ratio | 1.948 ± 0.406 b | 2.288 ± 0.44 ab | 2.451 ± 0.1606 ab | 2.2152 ± 0.0771 ab | 2.957 ± 1.096 a | 2.87 ± 0.635 a |
| Closed stomata area (µm2) | 29.4 ± 2.26 a b | 26.18 ± 15.42 b | 60.51 ± 9.43 a | 21.81 ± 12.36 b | 88.2 ± 72.9 a | 40.45 ± 14.81 ab |
| Opened stomata length (µm) | 16.78 ± 1.516 a | 48.9 ± 53.7 a | 14.26 ± 3.61 a | 8.652 ± 2.046 b | 19.263 ± 3.033 a | 16.922 ± 3.069 a |
| Opened stomata width (µm) | 9.323 ± 2.19 ab | 26 ± 25.6 a | 6.919 ± 1.473 b | 4.567 ± 1.076 c | 9.177 ± 1.37 ab | 8.577 ± 1.765 ab |
| Opened stomata length/width ratio | 1.859 ± 0.337 a | 1.666 ± 0.294 a | 2.076 ± 0.358 a | 1.921 ± 0.321 a | 2.114 ± 0.276 a | 2.006 ± 0.322 a |
| Opened stomata area (µm2) | 122.3 ± 37.4 a | 2099 ± 2996 a | 80.6 ± 36.9 a | 27.31 ± 11.71 b | 132.6 ± 38 a | 110 ± 43.2 a |
| Stomatal pore length (µm) | 11.07 ± 2.99 a | 41.2 ± 48.1 a | 9.91 ± 2.86 a | 5.212 ± 1.202 b | 13.27 ± 2.975 a | 11.766 ± 2.614 a |
| Stomatal pore width (µm) | 5.004 ± 2.176 a | 12.97 ± 12.99 a | 4.021 ± 1.12 ab | 2.319 ± 1.255 b | 3.611 ± 0.937 ab | 4.127 ± 1.374 a |
| Stomatal pore length/width ratio | 2.411 ± 0.663 b | 2.621 ± 0.735 ab | 2.491 ± 0.336 b | 2.555 ± 0.816 b | 3.808 ± 0.894 a | 3 ± 0.631 ab |
| Stomatal pore area (µm2) | 38.8 ± 26.9 a | 743 ± 1063 a | 29.06 ± 14.69 a | 8.29 ± 5.11 b | 33.36 ± 18.13 a | 35.25 ± 18.73 a |
| Epidermal cell length (µm) | 25.4 ± 3.07 bc | 30.57 ± 5.12 ab | 22.42 ± 2.7 c | 13.909 ± 1.818 d | 35.32 ± 4.51 a | 31.94 ± 5.65 ab |
| Epidermal cell width (µm) | 11.47 ± 3.68 bc | 21.26 ± 5 a | 10.491 ± 2.224 c | 10.75 ± 2.97 c | 17.83 ± 3.96 a | 17.06 ± 5.43 ab |
| Epidermal cell length/width ratio | 2.369 ± 0.663 a | 1.494 ± 0.393 ab | 2.224 ± 0.54 a | 1.389 ± 0.453 b | 2.054 ± 0.428 ab | 1.977 ± 0.52 ab |
| Epidermal cell area (µm2) | 260.4 ± 85.1 b | 600.9 ± 246.8 a | 199.8 ± 44.1 bc | 123.7 ± 38.5 c | 674.6 ± 121.5 a | 623.7 ± 200.5 a |
| Variable | Mean ± StDev | SE Mean | Min. | Q1 | Median | Q3 | Max. | IQR |
|---|---|---|---|---|---|---|---|---|
| Gravel (%) | 3.533 ± 1.317 c | 0.538 | 1.7 | 2.375 | 3.75 | 4.25 | 5.6 | 1.875 |
| Sand (%) | 73.5 ± 13.05 abc | 5.33 | 60 | 61.5 | 72 | 86.25 | 90 | 24.75 |
| Silt (%) | 19 ± 9.25 c | 3.78 | 8 | 9.5 | 19.5 | 27.75 | 30 | 18.25 |
| Clay (%) | 7.5 ± 4.04 c | 1.65 | 2 | 4.25 | 7.5 | 11.25 | 12 | 7 |
| SP (%) | 50.67 ± 6.98 abc | 2.85 | 39 | 45.75 | 51 | 56.75 | 59 | 11 |
| pH | 7.5483 ± 0.2046 c | 0.0835 | 7.3 | 7.39 | 7.55 | 7.6475 | 7.91 | 0.2575 |
| EC (dS/m) | 17.3 ± 23.75 c | 9.7 | 4.05 | 5.96 | 8.25 | 24.71 | 65.55 | 18.75 |
| Ca++ (meq/L) | 118.2 ± 169.8 abc | 69.3 | 28 | 43.8 | 55 | 159.5 | 464 | 115.8 |
| Mg++ (meq/L) | 63.1 ± 89.8 abc | 36.7 | 14.6 | 23.1 | 29.5 | 86.3 | 246 | 63.1 |
| Na+ (meq/L) | 79.1 ± 115.8 abc | 47.3 | 19.6 | 25.9 | 36.5 | 108 | 315 | 82.1 |
| K+ (meq/L) | 3.217 ± 0.999 c | 0.408 | 2.1 | 2.475 | 2.9 | 4.25 | 4.7 | 1.775 |
| HCO3− (meq/L) | 9.18 ± 3.8 c | 1.55 | 5.2 | 6.17 | 8.45 | 11.88 | 16 | 5.7 |
| Cl− (meq/L) | 73 ± 100.6 abc | 41.1 | 20.2 | 28.3 | 35.3 | 98.4 | 278 | 70.1 |
| SO4−− (meq/L) | 181 ± 272 ab | 111 | 39 | 61 | 80 | 248 | 736 | 187 |
| SAR | 7.1 ± 4.75 c | 1.94 | 4.2 | 4.5 | 5.65 | 8.52 | 16.7 | 4.02 |
| N (ppm) | 0.93 ± 0.275 c | 0.112 | 0.56 | 0.643 | 0.975 | 1.15 | 1.3 | 0.508 |
| P (ppm) | 9.65 ± 3.7 c | 1.51 | 6 | 6.97 | 8.3 | 13 | 16 | 6.03 |
| K (ppm) | 192.2 ± 78.5 a | 32.1 | 137 | 146 | 167 | 225.8 | 348 | 79.8 |
| Fe (ppm) | 0.643 ± 0.279 c | 0.114 | 0.28 | 0.408 | 0.62 | 0.907 | 1.02 | 0.5 |
| Zn++ (ppm) | 0.3217 ± 0.0679 c | 0.0277 | 0.27 | 0.2775 | 0.295 | 0.3675 | 0.45 | 0.09 |
| Mn++ (ppm) | 0.2883 ± 0.1158 c | 0.0473 | 0.18 | 0.2175 | 0.23 | 0.405 | 0.48 | 0.1875 |
| Cu++ (ppm) | 0.2133 ± 0.1724 c | 0.0704 | 0.03 | 0.06 | 0.175 | 0.3925 | 0.46 | 0.3325 |
| O.M. (%) | 0.1267 ± 0.0758 c | 0.0309 | 0.06 | 0.06 | 0.1 | 0.215 | 0.23 | 0.155 |
| CaCO3 (%) | 19.67 ± 3.41 c | 1.39 | 15.7 | 16.9 | 19 | 22.75 | 25 | 5.85 |
| Rt | Phytochemical Compounds | M.F | M.W | Mean ± StDev | SE Mean | Min. | Q1 | Median | Q3 | Max. | IQR |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 10.97 | Isophytol | C20H40O | 296 | 1.383 ± 0.965c | 0.394 | 0.42 | 0.495 | 1.3 | 2.03 | 3.08 | 1.535 |
| 13.53 | n-Hexadecanoic acid | C16H32O2 | 256 | 29.78 ± 6.15a | 2.51 | 22.04 | 23.66 | 30.67 | 35.21 | 36.25 | 11.55 |
| 16.88 | 9,12,15-Octadecatrienoic acid | C18H30O2 | 278 | 22.65 ± 3.92b | 1.6 | 17.31 | 18.98 | 23.26 | 25.97 | 26.86 | 6.99 |
| 15.27 | Oleic acid | C18H34O2 | 282 | 0.22 ± 0.1361c | 0.0556 | 0.13 | 0.1375 | 0.17 | 0.2875 | 0.49 | 0.15 |
| 17.10 | Octadecanoic acid | C18H36O2 | 284 | 3.63 ± 0.676c | 0.276 | 3 | 3.203 | 3.5 | 3.912 | 4.94 | 0.71 |
| 19.24 | Tributyl acetylcitrate | C20H34O8 | 402 | 1.552 ± 1.54c | 0.629 | 0.15 | 0.165 | 1.145 | 3.25 | 3.55 | 3.085 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nour, I.H.; Alhadead, K.; Ellmouni, F.Y.; Badr, R.; Saad, T.I.; EL-Banhawy, A.; Abdel Rahman, S.M. Morphological, Anatomical and Chemical Characterization of Ricinus communis L. (Euphorbiaceae). Agronomy 2023, 13, 985. https://doi.org/10.3390/agronomy13040985
Nour IH, Alhadead K, Ellmouni FY, Badr R, Saad TI, EL-Banhawy A, Abdel Rahman SM. Morphological, Anatomical and Chemical Characterization of Ricinus communis L. (Euphorbiaceae). Agronomy. 2023; 13(4):985. https://doi.org/10.3390/agronomy13040985
Chicago/Turabian StyleNour, Iman H., Khadiga Alhadead, Faten Y. Ellmouni, Reem Badr, Tamannouha I. Saad, Ahmed EL-Banhawy, and Salwa M. Abdel Rahman. 2023. "Morphological, Anatomical and Chemical Characterization of Ricinus communis L. (Euphorbiaceae)" Agronomy 13, no. 4: 985. https://doi.org/10.3390/agronomy13040985
APA StyleNour, I. H., Alhadead, K., Ellmouni, F. Y., Badr, R., Saad, T. I., EL-Banhawy, A., & Abdel Rahman, S. M. (2023). Morphological, Anatomical and Chemical Characterization of Ricinus communis L. (Euphorbiaceae). Agronomy, 13(4), 985. https://doi.org/10.3390/agronomy13040985