
Physiological and Biochemical Mechanisms of Exogenous Melatonin Regulation of Saline–Alkali Tolerance in Oats

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Solution Preparation
2.3. Experiment 1: Effect of MT on Seed Germination of Oats under Soda Saline–Alkali Stress
2.3.1. Seed Germination Index
2.3.2. Morphological and Biomass Indicators
2.3.3. A Comprehensive Evaluation of Different MT Treatments to Improve the Saline–Alkali Tolerance of Oats
2.4. Experiment 2: Study of the Intrinsic Physiological Mechanism of Oat Seed Germination under Saline–Alkali Stress and MT Treatment
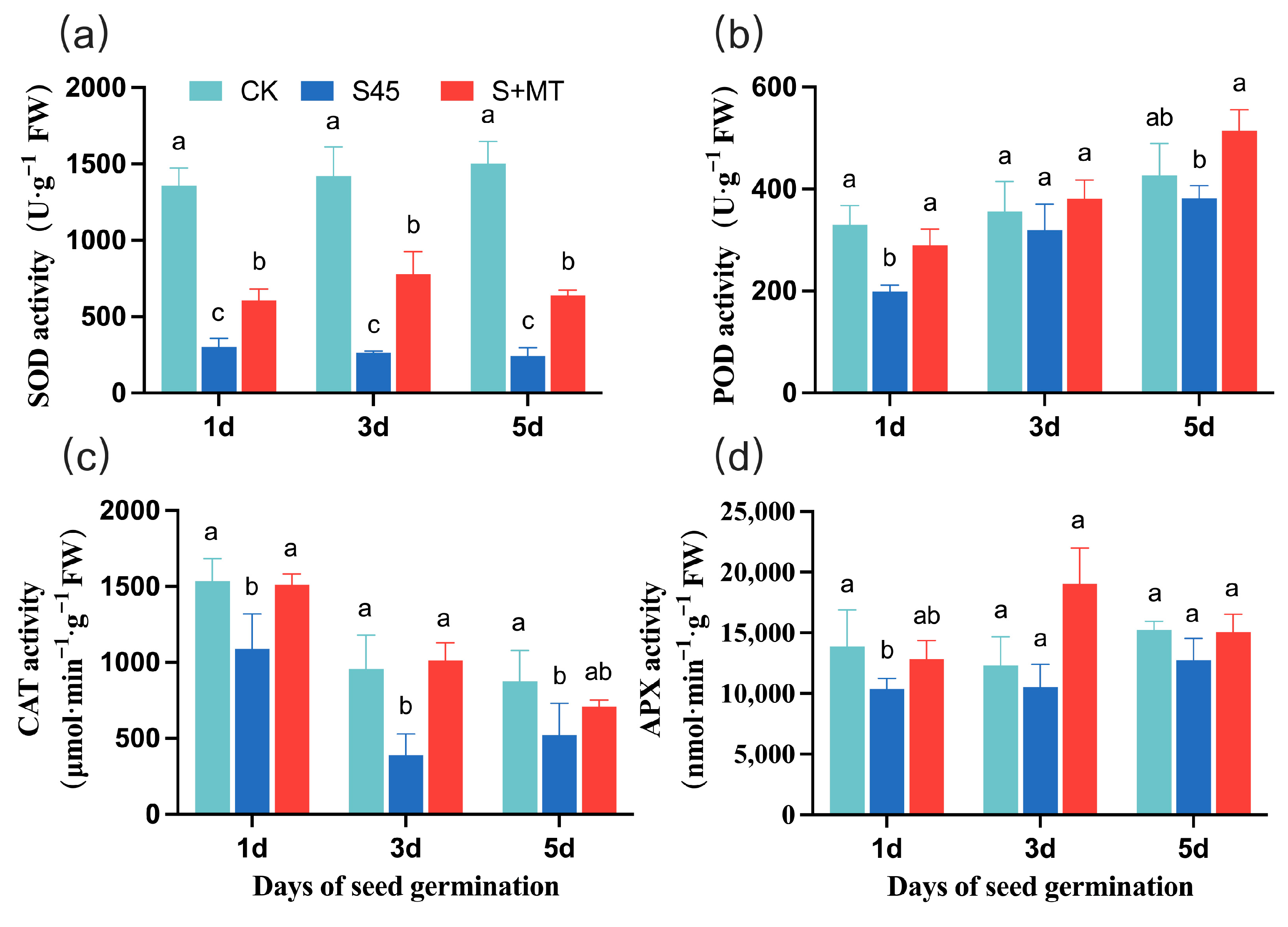
2.4.1. Antioxidant Enzyme Activity
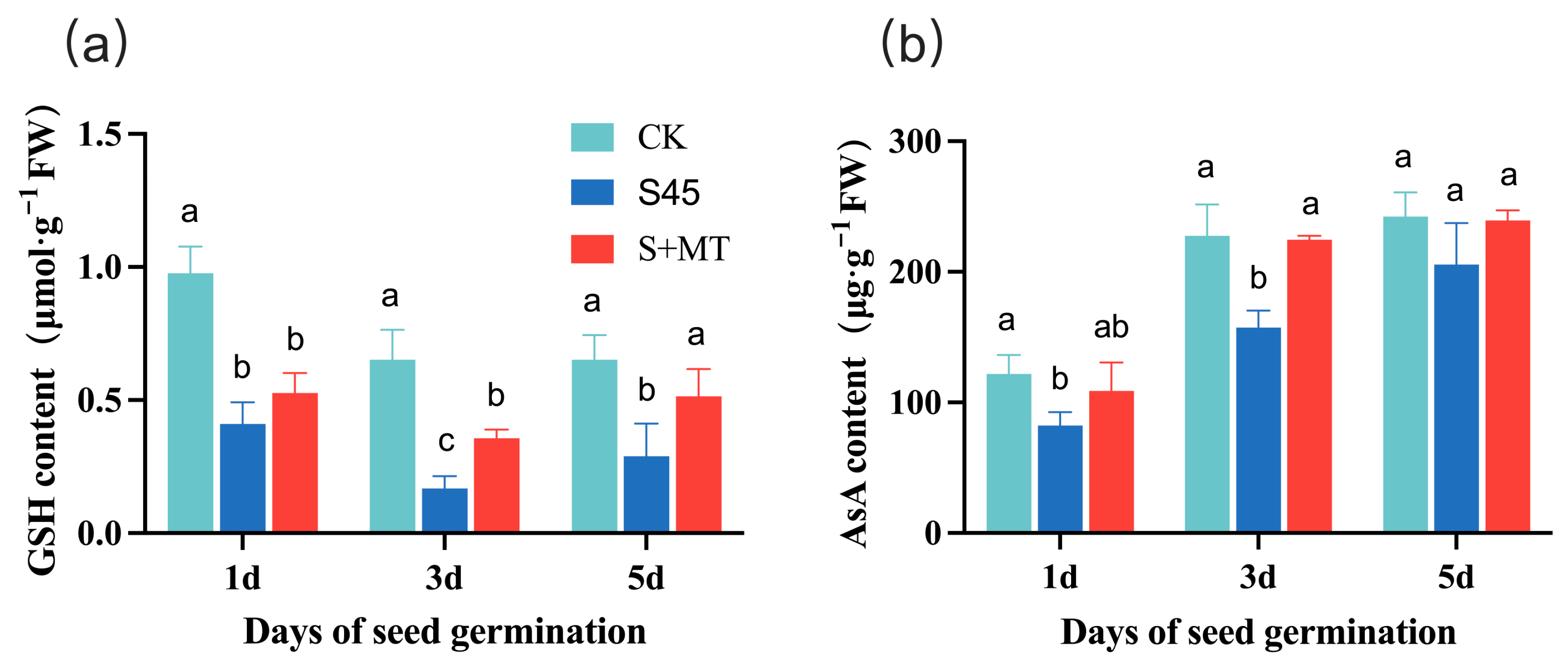
2.4.2. Nonenzymatic Antioxidant Activity
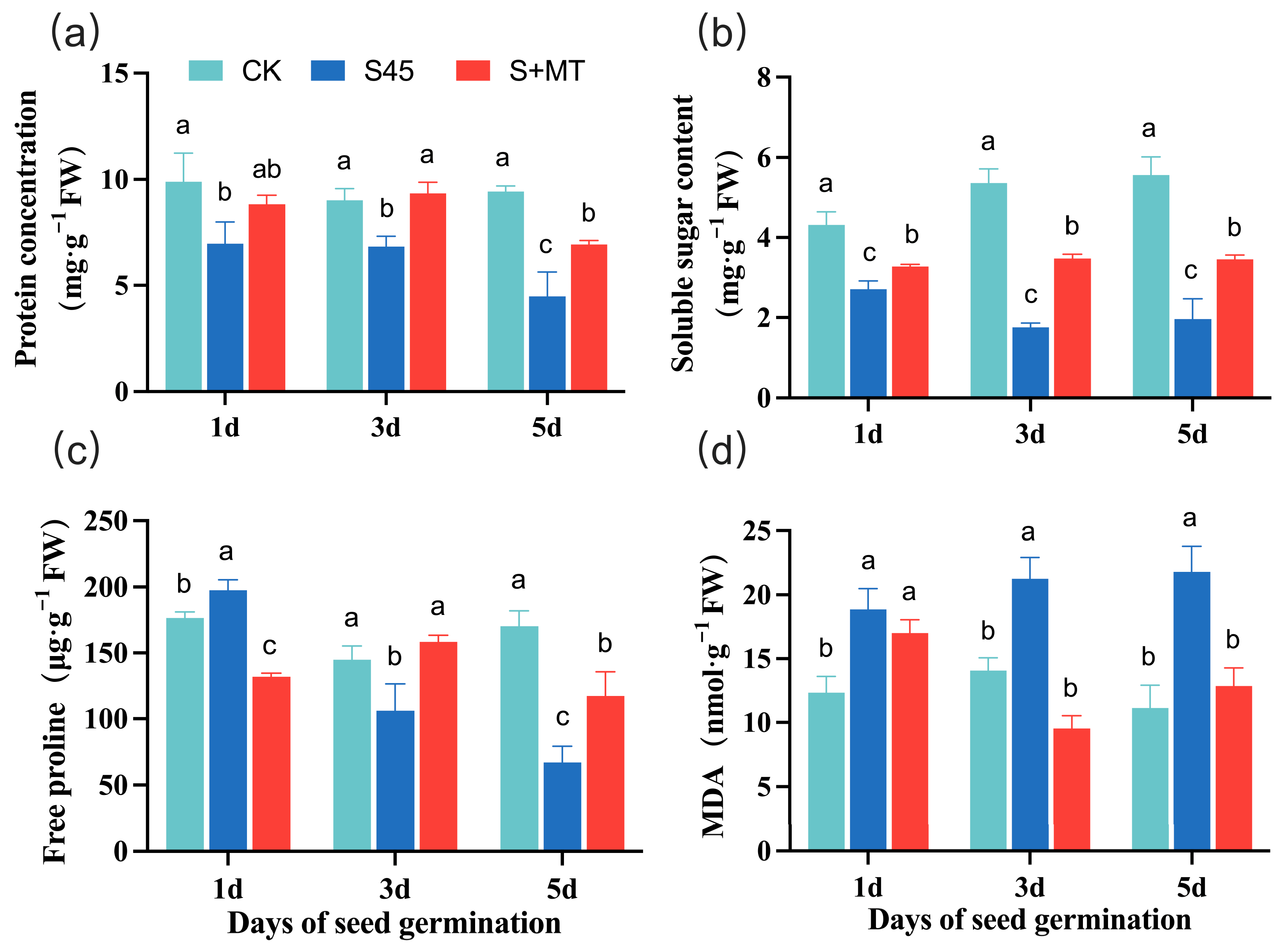
2.4.3. MDA Content
2.4.4. Osmoregulatory Substances
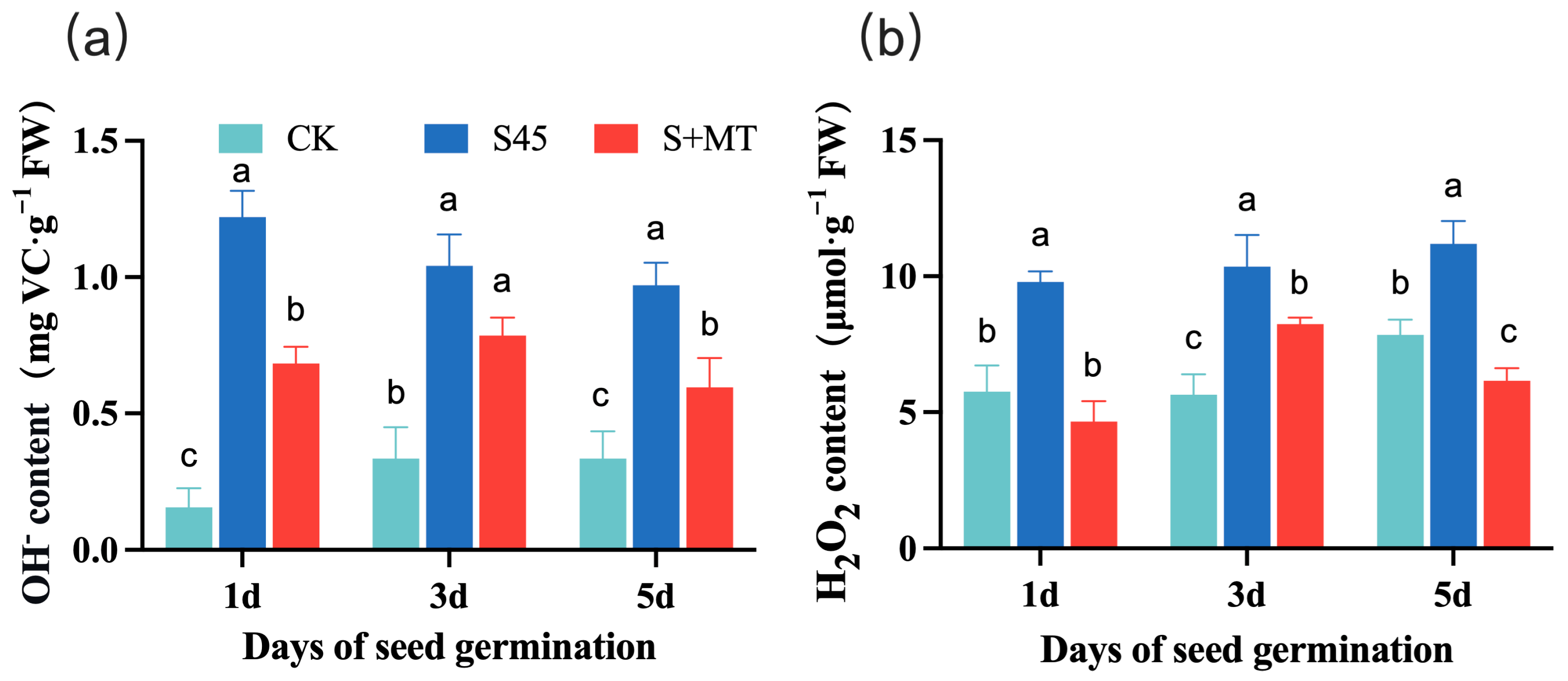
2.4.5. H2O2 Content and OH− Scavenging Capacity
2.5. Statistical Analysis
3. Results
3.1. Effect of Different Concentrations of MT Seed Soaking Treatment on Seed Germination of Oats under Saline–Alkali Stress
3.1.1. Effect on Seed Germination Indicators
3.1.2. Effect on Morphological Biomass
3.1.3. A Comprehensive Evaluation of the Saline–Alkali Tolerance of Oat Seeds during Germination
3.2. Effect of MT Seed Soaking Treatment on Physiological Indicators of Oats under Saline–Alkali Stress
3.2.1. Effect on the Activity of Antioxidant Enzymes
3.2.2. Effect on the Activity of Nonenzymatic Antioxidants
3.2.3. Effects on Osmoregulatory Substances and the Degree of Membrane Peroxidation
3.2.4. Effect on ROS Levels
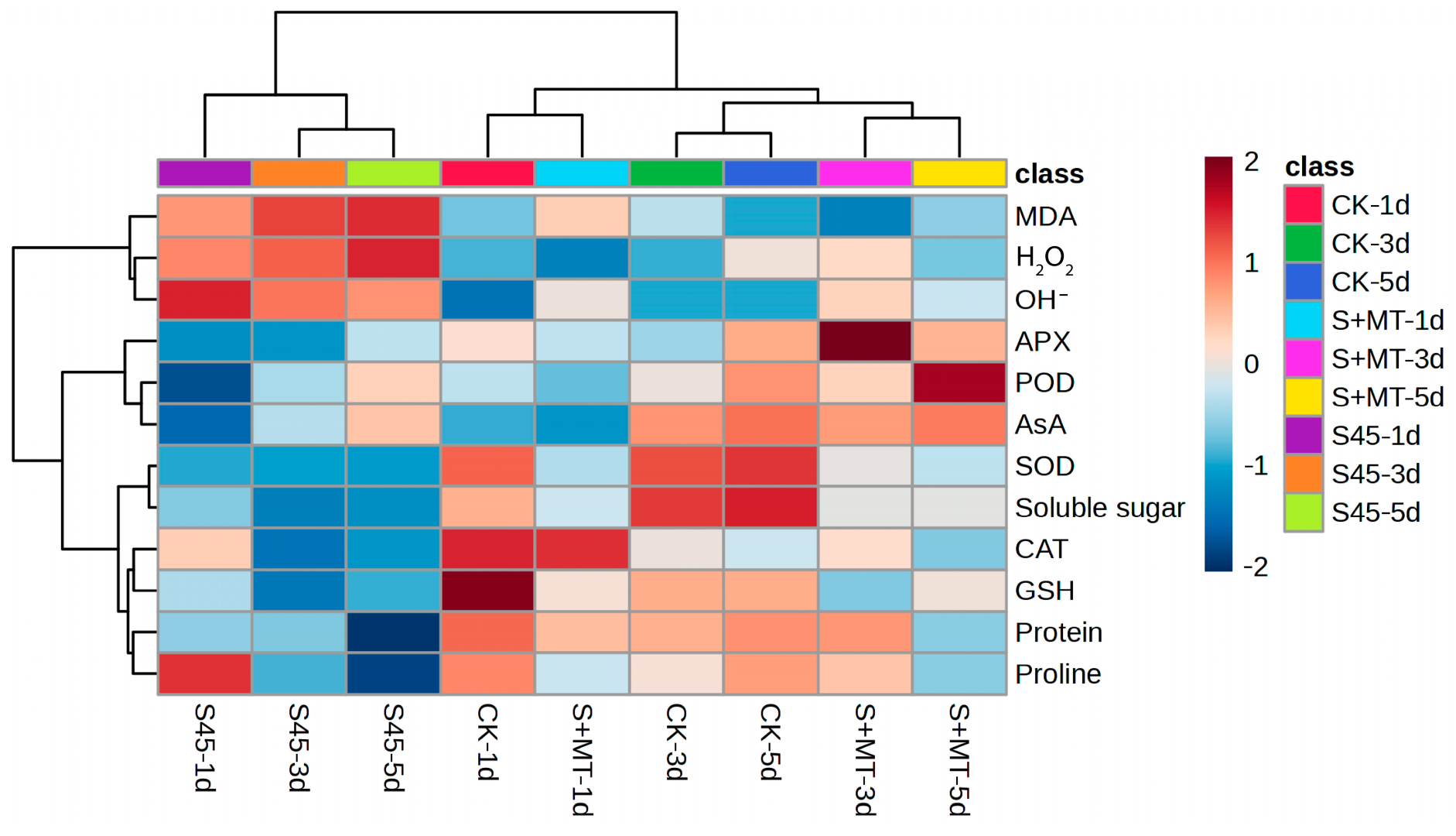
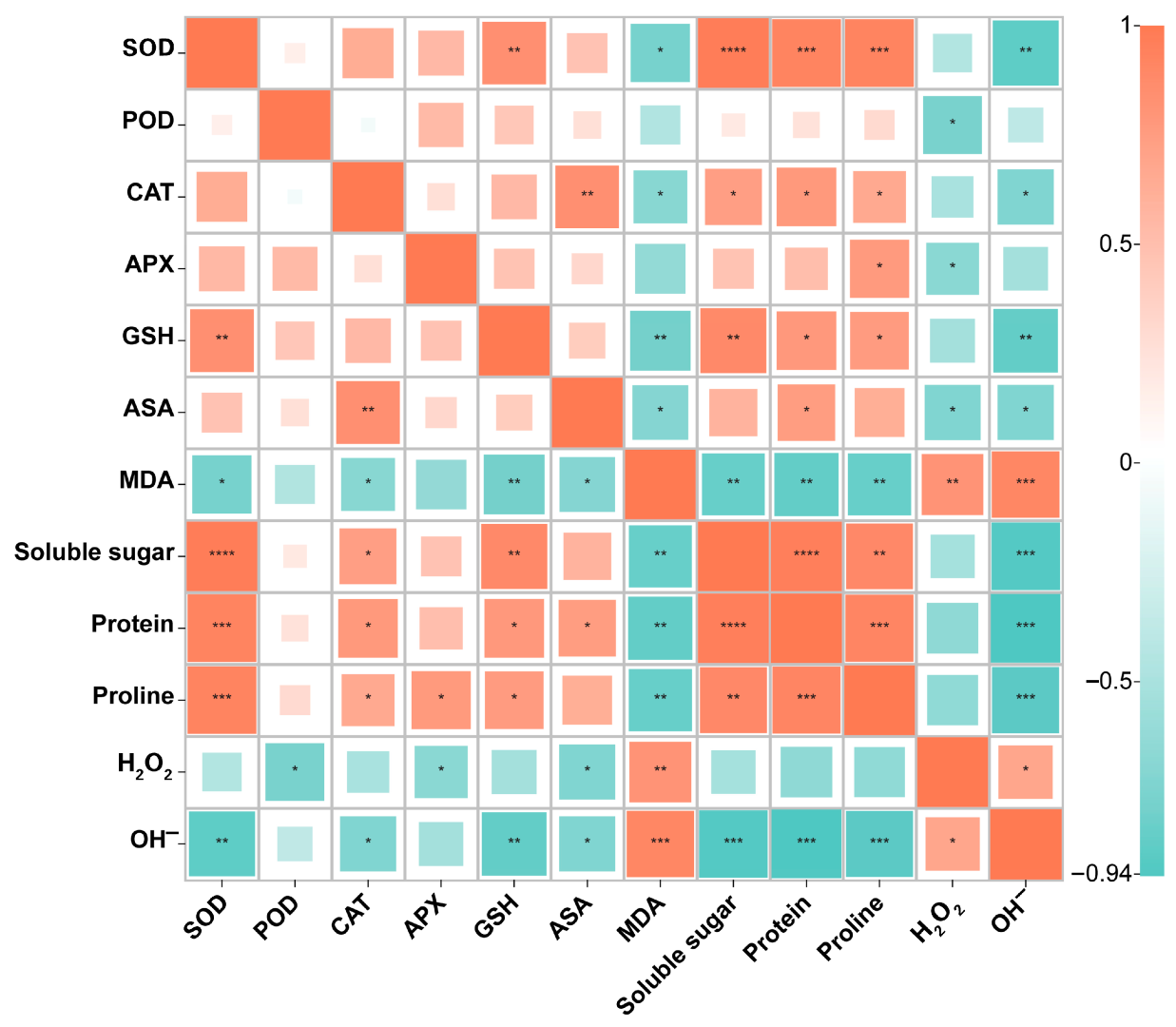
3.2.5. Evaluation of MT under Saline–Alkali Stress Using Hierarchical Cluster Analysis and Correlation Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composition | mg·L−1 |
|---|---|
| Ca (NO3)2·4H2O | 945 |
| KNO3 | 506 |
| NH4NO3 | 80 |
| KH2PO4 | 126 |
| MgSO4 | 241 |
| FeNaEDTA | 36.7 |
| KI | 0.83 |
| H3BO3 | 6.2 |
| MnSO4·H2O | 16.9 |
| ZnSO4·7H2O | 8.6 |
| Na2MoO4·2H2O | 0.25 |
| CuSO4·5H2O | 0.025 |
| CoCl2·6H2O | 0.025 |
| Totals | 1977.53 |
| pH (25 °C) | 5.8 ± 0.2 |
References
- Shabala, S. Learning from halophytes: Physiological basis and strategies to improve abiotic stress tolerance in crops. Ann. Bot. 2013, 112, 1209–1221. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Pu, L.; Han, M.; Zhu, M.; Zhang, R.; Xiang, Y. Soil salinization research in China: Advances and prospects. J. Geogr. Sci. 2014, 24, 943–960. [Google Scholar] [CrossRef]
- Amirinejad, A.-A.; Sayyari, M.; Ghanbari, F.; Kordi, S. Salicylic acid improves salinity-alkalinity tolerance in pepper (Capsicum annuum L.). Adv. Hort. Sci. 2017, 31, 157–163. [Google Scholar] [CrossRef]
- Wang, X.-S.; Ren, H.-L.; Wei, Z.-W.; Wang, Y.-W.; Ren, W.-B. Effects of neutral salt and alkali on ion distributions in the roots, shoots, and leaves of two alfalfa cultivars with differing degrees of salt tolerance. J. Integr. Agric. 2017, 16, 1800–1807. [Google Scholar] [CrossRef]
- Feng, N.; Yu, M.; Li, Y.; Jin, D.; Zheng, D. Prohexadione-calcium alleviates saline-alkali stress in soybean seedlings by improving the photosynthesis and up-regulating antioxidant defense. Ecotoxicol. Environ. Saf. 2021, 220, 112369. [Google Scholar] [CrossRef]
- Yang, L.; Wang, Y.; Yang, K. Klebsiella variicola improves the antioxidant ability of maize seedlings under saline-alkali stress. Peer J. 2021, 9, e11963. [Google Scholar] [CrossRef]
- Bai, Y.; Xiao, S.; Zhang, Z.; Zhang, Y.; Sun, H.; Zhang, K.; Wang, X.; Bai, Z.; Li, C.; Liu, L. Melatonin improves the germination rate of cotton seeds under drought stress by opening pores in the seed coat. Peer J. 2020, 8, e9450. [Google Scholar] [CrossRef]
- Chang, J.; Guo, Y.; Zhang, Z.; Wei, C.; Zhang, Y.; Ma, J.; Yang, J.; Zhang, X.; Li, H. CBF-responsive pathway and phytohormones are involved in melatonin-improved photosynthesis and redox homeostasis under aerial cold stress in watermelon. Acta Physiol. Plant. 2020, 42, 159. [Google Scholar] [CrossRef]
- He, J.; Zhuang, X.; Zhou, J.; Sun, L.; Wan, H.; Li, H.; Lyu, D. Exogenous melatonin alleviates cadmium uptake and toxicity in apple rootstocks. Tree Physiol. 2020, 40, 746–761. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Growth conditions influence the melatonin content of tomato plants. Food Chem. 2013, 138, 1212–1214. [Google Scholar] [CrossRef]
- Ye, J.; Yang, W.; Li, Y.; Wang, S.; Yin, L.; Deng, X. Seed pre-soaking with melatonin improves wheat yield by delaying leaf senescence and promoting root development. Agronomy 2020, 10, 84. [Google Scholar] [CrossRef]
- Silalert, P.; Pattanagul, W. Foliar application of melatonin alleviates the effects of drought stress in rice (Oryza sativa L.) seedlings. Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 12417. [Google Scholar] [CrossRef]
- Yan, F.; Zhang, J.; Li, W.; Ding, Y.; Zhong, Q.; Xu, X.; Wei, H.; Li, G. Exogenous melatonin alleviates salt stress by improving leaf photosynthesis in rice seedlings. Plant Physiol. Biochem. 2021, 163, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Liu, L.; Wang, X.; Wang, Z.; Zhang, H.; Chen, J.; Liu, X.; Wang, Y.; Li, C. Beneficial effects of exogenous melatonin on overcoming salt stress in sugar beets (Beta vulgaris L.). Plants 2021, 10, 886. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Chen, D.; Zhang, X.; Song, L.; Dong, J.; Xu, Q.; Hu, M.; Cheng, Y.; Shen, F.; Wang, W. Mitigation of salt stress response in upland cotton (Gossypium hirsutum) by exogenous melatonin. J. Plant Res. 2021, 134, 857–871. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lv, P.; Yan, D.; Zhang, Z.; Xu, X.; Wang, T.; Wang, Y.; Peng, Z.; Yu, C.; Gao, Y.; et al. Exogenous melatonin improves seed germination of wheat (Triticum aestivum L.) under salt stress. Int. J. Mol. Sci. 2022, 23, 8436. [Google Scholar] [CrossRef]
- Chen, L.; Lu, B.; Liu, L.; Duan, W.; Jiang, D.; Li, J.; Zhang, K.; Sun, H.; Zhang, Y.; Li, C.; et al. Melatonin promotes seed germination under salt stress by regulating ABA and GA3 in cotton (Gossypium hirsutum L.). Plant Physiol. Biochem. 2021, 162, 506–516. [Google Scholar] [CrossRef]
- Yu, Y.; Deng, L.; Zhou, L.; Chen, G.; Wang, Y. Exogenous melatonin activates antioxidant systems to increase the ability of rice seeds to germinate under high temperature conditions. Plants 2022, 11, 886. [Google Scholar] [CrossRef]
- Zhang, H.-J.; Zhang, N.; Yang, R.-C.; Wang, L.; Sun, Q.-Q.; Li, D.-B.; Cao, Y.-Y.; Weeda, S.; Zhao, B.; Ren, S.; et al. Melatonin promotes seed germination under high salinity by regulating antioxidant systems, ABA and GA4 interaction in cucumber (Cucumis sativus L.). J. Pineal Res. 2014, 57, 269–279. [Google Scholar] [CrossRef]
- Cao, Q.; Li, G.; Cui, Z.; Yang, F.; Jiang, X.; Diallo, L.; Kong, F. Seed priming with melatonin improves the seed germination of waxy maize under chilling stress via promoting the antioxidant system and starch metabolism. Sci. Rep. 2019, 9, 15044. [Google Scholar] [CrossRef]
- Lv, Y.; Pan, J.; Wang, H.; Reiter, R.J.; Li, X.; Mou, Z.; Zhang, J.; Yao, Z.; Zhao, D.; Yu, D. Melatonin inhibits seed germination by crosstalk with abscisic acid, gibberellin, and auxin in Arabidopsis. J. Pineal Res. 2021, 70, e12736. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Wang, H.; Chen, W.; You, Q.; Li, X.; Yu, D. Phytomelatonin inhibits seed germination by regulating germination-related hormone signaling in Arabidopsis. Plant Signal. Behav. 2021, 16, 1970447. [Google Scholar] [CrossRef] [PubMed]
- Butt, M.S.; Tahir-Nadeem, M.; Khan, M.K.I.; Shabir, R.; Butt, M.S. Oat: UNIQUE among the cereals. Eur. J. Nutr. 2008, 47, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Rafique, H.; Dong, R.; Wang, X.; Alim, A.; Aadil, R.M.; Li, L.; Zou, L.; Hu, X. Dietary-nutraceutical properties of oat protein and peptides. Front. Nutr. 2022, 9, 950400. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Ren, C.Z.; Li, P.F.; Ren, T.S. Oat growth and cation absorption characteristics under salt and alkali stress. Ying Yong Sheng Tai Xue Bao 2011, 22, 2875–2882. [Google Scholar]
- Zhao, B.P.; Liu, J.H.; Wu, J.Y.; Liu, H.; Xu, H. Effects of salt stress on plas-malemma permeability, osmolyte accumulation and protective enzyme activities in oat plants. J. Food Agric. Environ. 2013, 11, 696–701. [Google Scholar]
- Zhang, Y.Q.; Wu, N.; Liu, J.L.; Yang, N.N.; Yang, Y.Y. Effects of mild saline-alkali on photosynthetic physiology and yield of different oat cultivars. Southwest China J. Agri. Sci. 2018, 31, 2041–2046. [Google Scholar]
- Xiao, Z.Z.; Sui, X.Q.; Shi, G.Q.; Chen, A.P.; Zhang, B. Effects of different concentrations and immersion durations of exogenous melatonin on seed germination of bromus inermis under drought stress. Acta Agrestia Sin. 2022, 30, 655–660. [Google Scholar] [CrossRef]
- Ukeda, H.; Kawana, D.; Maeda, S.; Sawamura, M. Spectrophotometric assay for superoxide dismutase based on the reduction of highly water-soluble tetrazolium salts by xanthine-xanthine oxidase. Biosci. Biotechnol. Biochem. 1999, 63, 485–488. [Google Scholar] [CrossRef]
- Kochba, J.; Lavee, S.; Spiegel-Roy, P. Differences in peroxidase activity and isoenzymes in embryogenic ane non-embryogenic ‘Shamouti’ orange ovular callus lines. Plant Cell Physiol. 1977, 18, 463–467. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Ullah, S.; Kolo, Z.; Egbichi, I.; Keyster, M.; Ludidi, N. Nitric oxide influences glycine betaine content and ascorbate peroxidase activity in maize. S. Afr. J. Bot. 2016, 105, 218–225. [Google Scholar] [CrossRef]
- Nishimoto, S.; Koike, S.; Inoue, N.; Suzuki, T.; Ogasawara, Y. Activation of Nrf2 attenuates carbonyl stress induced by methylglyoxal in human neuroblastoma cells: Increase in GSH levels is a critical event for the detoxification mechanism. Biochem. Biophys. Res. Commun. 2017, 483, 874–879. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Cullen, J.J.; Buettner, G.R. Ascorbic acid: Chemistry, biology and the treatment of cancer. Biochim. Biophys. Acta 2012, 1826, 443–457. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Buysse, J.A.N.; Merckx, R. An improved colorimetric method to quantify sugar content of plant tissue. J. Exp. Bot. 1993, 44, 1627–1629. [Google Scholar] [CrossRef]
- Mansour, M.M.F.; Ali, E.F. Evaluation of proline functions in saline conditions. Phytochemistry 2017, 140, 52–68. [Google Scholar] [CrossRef]
- Campion, E.M.; Loughran, S.T.; Walls, D. Protein quantitation and analysis of purity. Methods Mol. Biol. 2011, 681, 229–258. [Google Scholar] [CrossRef]
- Satterfield, C.N.; Bonnell, A.H. Interferences in titanium sulfate method for hydrogen peroxide. Anal. Chem. 1955, 27, 1174–1175. [Google Scholar] [CrossRef]
- Sutton, H.C.; Winterbourn, C.C. On the participation of higher oxidation states of iron and copper in fenton reactions. Free Radic. Biol. Med. 1989, 6, 53–60. [Google Scholar] [CrossRef]
- Ibrahim, E.A. Seed priming to alleviate salinity stress in germinating seeds. J. Plant Physiol. 2016, 192, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Khoso, M.A.; Hussain, A.; Ritonga, F.N.; Ali, Q.; Channa, M.M.; Alshegaihi, R.M.; Meng, Q.; Ali, M.; Zaman, W.; Brohi, R.D.; et al. WRKY transcription factors (TFs): Molecular switches to regulate drought, temperature, and salinity stresses in plants. Front. Plant Sci. 2022, 13, 1039329. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yang, F.; Ma, H.; Li, J. Evaluation of the saline-alkaline tolerance of rice (Oryza sativa L.) mutants induced by heavy-ion beam mutagenesis. Biology 2022, 11, 126. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Ren, L.; Sun, H.; Qiao, K.; Liu, S.; Zhou, A. Morphological and physiological responses of two willow species from different habitats to salt stress. Sci. Rep. 2020, 10, 18228. [Google Scholar] [CrossRef]
- Cheng, C.; Liu, J.; Wang, Z.; Liu, J.; Wang, Y.; Liao, Y.; Gao, Z.; Lu, Z.; Zhu, B.; Yao, F. Analysis of effect of compound salt stress on seed germination and salt tolerance analysis of pepper (Capsicum annuum L.). J. Vis. Exp. 2022, 189, e64702. [Google Scholar] [CrossRef]
- Hardeland, R. Taxon- and site-specific melatonin catabolism. Molecules 2017, 22, 2015. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.; Kong, K.; Park, S.; Natsagdorj, U.; Kim, Y.S.; Back, K. Molecular cloning of a plant N-acetylserotonin methyltransferase and its expression characteristics in rice. J. Pineal Res. 2011, 50, 304–309. [Google Scholar] [CrossRef]
- Wang, L.; Feng, C.; Zheng, X.; Guo, Y.; Zhou, F.; Shan, D.; Liu, X.; Kong, J. Plant mitochondria synthesize melatonin and enhance the tolerance of plants to drought stress. J. Pineal Res. 2017, 63, e12429. [Google Scholar] [CrossRef]
- Li, C.; Wang, P.; Wei, Z.; Liang, D.; Liu, C.; Yin, L.; Jia, D.; Fu, M.; Ma, F. The mitigation effects of exogenous melatonin on salinity-induced stress in Malus hupehensis. J. Pineal Res. 2012, 53, 298–306. [Google Scholar] [CrossRef]
- Bajwa, V.S.; Shukla, M.R.; Sherif, S.M.; Murch, S.J.; Saxena, P.K. Role of melatonin in alleviating cold stress in Arabidopsis thaliana. J. Pineal Res. 2014, 56, 238–245. [Google Scholar] [CrossRef]
- Xiao, S.; Liu, L.; Wang, H.; Li, D.; Bai, Z.; Zhang, Y.; Sun, H.; Zhang, K.; Li, C. Exogenous melatonin accelerates seed germination in cotton (Gossypium hirsutum L.). PLoS ONE 2019, 14, e0216575. [Google Scholar] [CrossRef] [PubMed]
- Posmyk, M.M.; Bałabusta, M.; Wieczorek, M.; Sliwinska, E.; Janas, K.M. Melatonin applied to cucumber (Cucumis sativus L.) seeds improves germination during chilling stress. J. Pineal Res. 2009, 46, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, L.; Lu, B.; Ma, T.; Jiang, D.; Li, J.; Zhang, K.; Sun, H.; Zhang, Y.; Bai, Z.; et al. Exogenous melatonin promotes seed germination and osmotic regulation under salt stress in cotton (Gossypium hirsutum L.). PLoS ONE 2020, 15, e0228241. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhao, C.; Zhang, M.; Yuan, F.; Chen, M. Exogenous melatonin improves seed germination in Limonium bicolor under salt stress. Plant Signal. Behav. 2019, 14, 1659705. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Gao, Y.; Tong, S.; Liu, B. Morphological and physiological traits related to the response and adaption of Bolboschoenus planiculmis seedlings grown under salt-alkaline stress conditions. Front. Plant Sci. 2021, 12, 567782. [Google Scholar] [CrossRef]
- Luo, X.; Dai, Y.; Zheng, C.; Yang, Y.; Chen, W.; Wang, Q.; Chandrasekaran, U.; Du, J.; Liu, W.; Shu, K. The ABI4-RbohD/VTC2 regulatory module promotes reactive oxygen species (ROS) accumulation to decrease seed germination under salinity stress. New Phytol. 2021, 229, 950–962. [Google Scholar] [CrossRef]
- Waisel, Y. Salt resistance. In Biology of Halophytes; Waisel, Y., Ed.; Academic Press: Cambridge, MA, USA, 1972; pp. 236–245. [Google Scholar]
- Huang, X.-S.; Wang, W.; Zhang, Q.; Liu, J.-H. A basic helix-loop-helix transcription factor, PtrbHLH, of Poncirus trifoliata confers cold tolerance and modulates peroxidase-mediated scavenging of hydrogen peroxide. Plant Physiol. 2013, 162, 1178–1194. [Google Scholar] [CrossRef]
- Matamoros, M.A.; Loscos, J.; Dietz, K.-J.; Aparicio-Tejo, P.M.; Becana, M. Function of antioxidant enzymes and metabolites during maturation of pea fruits. J. Exp. Bot. 2010, 61, 87–97. [Google Scholar] [CrossRef]
- Pastori, G.M.; Trippi, V.S. Antioxidative protection in a drought-resistant maize strain during leaf senescence. Physiol. Plant. 1993, 87, 227–231. [Google Scholar] [CrossRef]
- Khan, A.; Numan, M.; Khan, A.L.; Lee, I.-J.; Imran, M.; Asaf, S.; Al-Harrasi, A. Melatonin: Awakening the defense mechanisms during plant oxidative stress. Plants 2020, 9, 407. [Google Scholar] [CrossRef]
- Zhao, Q.; Chen, S.; Wang, G.; Du, Y.; Zhang, Z.; Yu, G.; Ren, C.; Zhang, Y.; Du, J. Exogenous melatonin enhances soybean (Glycine max (L.) Merr.) seedling tolerance to saline-alkali stress by regulating antioxidant response and DNA damage repair. Physiol. Plant. 2022, 174, e13731. [Google Scholar] [CrossRef] [PubMed]
- Sheikhalipour, M.; Mohammadi, S.A.; Esmaielpour, B.; Zareei, E.; Kulak, M.; Ali, S.; Nouraein, M.; Bahrami, M.K.; Gohari, G.; Fotopoulos, V. Exogenous melatonin increases salt tolerance in bitter melon by regulating ionic balance, antioxidant system and secondary metabolism-related genes. BMC Plant Biol. 2022, 22, 380. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.H.; Alamri, S.; Al-Khaishany, M.Y.; Khan, M.N.; Al-Amri, A.; Ali, H.M.; Alaraidh, I.A.; Alsahli, A.A. Exogenous melatonin counteracts NaCl-induced damage by regulating the antioxidant system, proline and carbohydrates metabolism in tomato seedlings. Int. J. Mol. Sci. 2019, 20, 353. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Song, H.; Zhang, L. New insight into plant saline-alkali tolerance mechanisms and application to breeding. Int. J. Mol. Sci. 2022, 23, 16048. [Google Scholar] [CrossRef] [PubMed]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef]
- Zhao, Q.; Suo, J.; Chen, S.; Jin, Y.; Ma, X.; Yin, Z.; Zhang, Y.; Wang, T.; Luo, J.; Jin, W.; et al. Na2CO3-responsive mechanisms in halophyte Puccinellia tenuiflora roots revealed by physiological and proteomic analyses. Sci. Rep. 2016, 6, 32717. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Lyu, J.; Yu, J.H.; Liu, N.; Li, J.W.; Jin, L.; Jin, N.; Wang, S.Y. Effects of melatonin on photosynthetic properties and osmoregulatory substance contents of cucumber seedlings under salt-alkali stress. Ying Yong Sheng Tai Xue Bao 2022, 33, 1901–1910. [Google Scholar] [CrossRef]
- Kamiab, F. Exogenous melatonin mitigates the salinity damages and improves the growth of pistachio under salinity stress. J. Plant Nutr. 2020, 43, 1468–1484. [Google Scholar] [CrossRef]
- Ding, F.; Liu, B.; Zhang, S. Exogenous melatonin ameliorates cold-induced damage in tomato plants. Sci. Hortic. 2017, 219, 264–271. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, L.; Li, H.; Zhang, S.; Fu, X.; Zhai, X.; Yang, N.; Shen, J.; Li, R.; Li, D. Exogenous melatonin promotes the salt tolerance by removing active oxygen and maintaining ion balance in wheat (Triticum aestivum L.). Front. Plant Sci. 2022, 12, 787062. [Google Scholar] [CrossRef]
- Luo, C.; Min, W.; Akhtar, M.; Lu, X.; Bai, X.; Zhang, Y.; Tian, L.; Li, P. Melatonin enhances drought tolerance in rice seedlings by modulating antioxidant systems, osmoregulation, and corresponding gene expression. Int. J. Mol. Sci. 2022, 23, 12075. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, Y.; Sun, P.; Chen, G.; Xin, J. Interactive effects of salt and alkali stresses on growth, physiological responses and nutrient (N, P) removal performance of Ruppia maritima. Ecol. Eng. 2017, 104, 177–183. [Google Scholar] [CrossRef]
- Mannino, G.; Pernici, C.; Serio, G.; Gentile, C.; Bertea, C.M. Melatonin and phytomelatonin: Chemistry, biosynthesis, metabolism, distribution and bioactivity in plants and animals-an overview. Int. J. Mol. Sci. 2021, 22, 9996. [Google Scholar] [CrossRef] [PubMed]
- Purushothaman, A.; Sheeja, A.A.; Janardanan, D. Hydroxyl radical scavenging activity of melatonin and its related indolamines. Free Radic. Res. 2020, 54, 373–383. [Google Scholar] [CrossRef]






| Treatment | GR (%) | GP (%) | GI | VI | |
|---|---|---|---|---|---|
| CK | 78.89 ± 1.92 | 68.89 ± 3.85 | 40 ± 2.39 | 502.3 ± 33.31 | |
| S45 + MT25 | 52.22 ± 1.92 | 45.56 ± 3.85 | 21.73 ± 0.44 | 217.67 ± 19.38 | |
| S45 + MT25 | 62.22 ± 3.85 ns | 54.44 ± 1.92 ns | 28.43 ± 0.67 ns | 307.24 ± 17.82 ns | |
| T12 | S45 + MT50 | 47.78 ± 9.62 ns | 38.89 ± 10.72 ns | 19.55 ± 4.45 ns | 192.9 ± 63.51 ns |
| S45 + MT75 | 63.33 ± 6.67 ns | 56.67 ± 5.77 ns | 29.49 ± 1.89 ns | 297.8 ± 15.02 ns | |
| S45 + MT100 | 70 ± 3.33 ns | 67.78 ± 1.92 ns | 34.62 ± 1.39 # | 389.06 ± 14.45 # | |
| S45 + MT150 | 58.89 ± 5.09 ns | 53.33 ± 0 ns | 28.92 ± 1.85 ns | 319.05 ± 27.87 ns | |
| S45 + MT200 | 57.78 ± 3.85 ns | 50 ± 12.02 ns | 25.06 ± 5.1 ns | 252.8 ± 67.58 ns | |
| CK | 86.67 ± 5.77 | 85.56 ± 7.7 | 46.55 ± 4.51 | 555.13 ± 56.97 | |
| S45 + MT0 | 52.22 ± 8.39 | 48.89 ± 11.71 | 22.7 ± 4.36 | 190.67 ± 42.77 | |
| S45 + MT25 | 71.11 ± 16.44 ns | 68.89 ± 15.4 ns | 33.7 ± 6.98 ns | 363.25 ± 83.56 * | |
| T24 | S45 + MT50 | 71.11 ± 12.62 ns | 64.44 ± 9.62 ns | 29.11 ± 4.46 ns | 308.45 ± 50.94 ns |
| S45 + MT75 | 66.67 ± 3.33 ns | 63.33 ± 6.67 ns | 29.88 ± 2.39 ns | 320.41 ± 37.37 ns | |
| S45 + MT100 | 85.56 ± 6.94 *** | 83.33 ± 8.82 ** | 38.44 ± 5.38 *** | 453.75 ± 71.55 **** | |
| S45 + MT150 | 74.44 ± 1.92 ns | 72.22 ± 3.85 ns | 32.85 ± 1.8 ns | 353.98 ± 15.39 * | |
| S45 + MT200 | 76.67 ± 10 * | 75.56 ± 8.39 * | 36.05 ± 6.18 * | 383.65 ± 83.1 ** | |
| Treatment | GL (cm) | RL (cm) | FW (mg) | DW (mg) | |
|---|---|---|---|---|---|
| CK | 12.55 ± 0.09 | 7.29 ± 0.34 | 162.93 ± 3.29 | 14.9 ± 0.42 | |
| S45 + MT0 | 10.01 ± 0.68 | 2.9 ± 0.58 | 111.99 ± 6.69 | 12.25 ± 0.59 | |
| S45 + MT25 | 10.81 ± 0.55 ns | 3.67 ± 0.52 ns | 120.15 ± 6.55 ns | 13.65 ± 0.75 ns | |
| T12 | S45 + MT50 | 9.7 ± 1.23 ns | 3.07 ± 0.96 ns | 102.09 ± 18.49 ns | 11.72 ± 2.17 ns |
| S45 + MT75 | 10.11 ± 0.51 ns | 3.83 ± 0.98 ns | 108.68 ± 5.42 ns | 12.55 ± 0.53 ns | |
| S45 + MT100 | 11.24 ± 0.03 ns | 4.83 ± 0.08 ns | 123.35 ± 4.08 ns | 13.65 ± 0.47 ns | |
| S45 + MT150 | 11.02 ± 0.31 ns | 4.17 ± 0.2 ns | 114.51 ± 4.18 ns | 13.35 ± 0.57 ns | |
| S45 + MT200 | 10.01 ± 0.65 ns | 3.26 ± 0.51 ns | 104.64 ± 0.74 ns | 12.32 ± 0.03 ns | |
| CK | 11.92 ± 0.07 | 8.45 ± 1.8 | 150.26 ± 13.68 | 14.17 ± 1.3 | |
| S45 + MT0 | 8.37 ± 0.47 | 2.91 ± 0.74 | 90.58 ± 2.74 | 10.22 ± 0.29 | |
| S45 + MT25 | 10.75 ± 0.24 *** | 6.11 ± 0.4 ** | 120.8 ± 5.94 ** | 12.75 ± 0.54 * | |
| T24 | S45 + MT50 | 10.58 ± 0.16 *** | 5.41 ± 1.29 * | 126.64 ± 8.11 *** | 13.23 ± 0.43 ** |
| S45 + MT75 | 10.7 ± 0.43 *** | 5.32 ± 0.18 * | 120.4 ± 6.43 ** | 12.42 ± 0.43 ns | |
| S45 + MT100 | 11.79 ± 0.21 **** | 6.06 ± 0.11 ** | 146.19 ± 4.63 **** | 14.48 ± 0.23 **** | |
| S45 + MT150 | 10.78 ± 0.16 **** | 5.67 ± 0.48 ** | 119.31 ± 0.44 ** | 12.6 ± 0.12 * | |
| S45 + MT200 | 10.58 ± 0.59 **** | 5.16 ± 0.86 ns | 124.45 ± 4.13 *** | 12.01 ± 0 *** | |
| Treatment | GR | GP | GI | VI | GL | RL | FW | DW | D Value | Rank |
|---|---|---|---|---|---|---|---|---|---|---|
| S45 + MT0 + T12 | 0.12 | 0.15 | 0.12 | 0.10 | 0.48 | 0.00 | 0.39 | 0.48 | 0.20 | 12 |
| S45 + MT0 + T24 | 0.12 | 0.23 | 0.17 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.06 | 14 |
| S45 + MT25 + T12 | 0.38 | 0.35 | 0.47 | 0.44 | 0.71 | 0.24 | 0.53 | 0.80 | 0.47 | 8 |
| S45 + MT25 + T24 | 0.62 | 0.68 | 0.75 | 0.66 | 0.70 | 1.01 | 0.54 | 0.59 | 0.65 | 3 |
| S45 + MT50 + T12 | 0.00 | 0.00 | 0.00 | 0.01 | 0.39 | 0.05 | 0.21 | 0.35 | 0.08 | 13 |
| S45 + MT50 + T24 | 0.62 | 0.57 | 0.51 | 0.45 | 0.65 | 0.79 | 0.65 | 0.71 | 0.57 | 5 |
| S45 + MT75 + T12 | 0.41 | 0.40 | 0.53 | 0.41 | 0.51 | 0.30 | 0.33 | 0.55 | 0.41 | 10 |
| S45 + MT75 + T24 | 0.50 | 0.55 | 0.55 | 0.49 | 0.68 | 0.77 | 0.54 | 0.52 | 0.53 | 6 |
| S45 + MT100 + T12 | 0.59 | 0.65 | 0.80 | 0.75 | 0.84 | 0.61 | 0.59 | 0.80 | 0.69 | 2 |
| S45 + MT100 + T24 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 0.99 | 1 |
| S45 + MT150 + T12 | 0.29 | 0.32 | 0.50 | 0.49 | 0.78 | 0.40 | 0.43 | 0.73 | 0.46 | 9 |
| S45 + MT150 + T24 | 0.71 | 0.75 | 0.70 | 0.62 | 0.71 | 0.88 | 0.52 | 0.56 | 0.64 | 4 |
| S45 + MT200 + T12 | 0.26 | 0.25 | 0.29 | 0.24 | 0.48 | 0.11 | 0.25 | 0.49 | 0.27 | 11 |
| S45 + MT200 + T24 | 0.76 | 0.83 | 0.87 | 0.73 | 0.65 | 0.72 | 0.61 | −2.4 | 0.50 | 7 |
| Wi | 0.14 | 0.13 | 0.08 | 0.31 | 0.04 | 0.03 | 0.19 | 0.07 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Xu, W.; Ren, C.; Zhan, C.; Wang, C.; Li, J.; Ren, Q.; Liang, X.; Wei, L.; Xiang, D.; et al. Physiological and Biochemical Mechanisms of Exogenous Melatonin Regulation of Saline–Alkali Tolerance in Oats. Agronomy 2023, 13, 1327. https://doi.org/10.3390/agronomy13051327
Wang Q, Xu W, Ren C, Zhan C, Wang C, Li J, Ren Q, Liang X, Wei L, Xiang D, et al. Physiological and Biochemical Mechanisms of Exogenous Melatonin Regulation of Saline–Alkali Tolerance in Oats. Agronomy. 2023; 13(5):1327. https://doi.org/10.3390/agronomy13051327
Chicago/Turabian StyleWang, Qiang, Weiwei Xu, Changzhong Ren, Chao Zhan, Chunlong Wang, Junwei Li, Qinyong Ren, Xiaotian Liang, Liming Wei, Dabing Xiang, and et al. 2023. "Physiological and Biochemical Mechanisms of Exogenous Melatonin Regulation of Saline–Alkali Tolerance in Oats" Agronomy 13, no. 5: 1327. https://doi.org/10.3390/agronomy13051327
APA StyleWang, Q., Xu, W., Ren, C., Zhan, C., Wang, C., Li, J., Ren, Q., Liang, X., Wei, L., Xiang, D., Wang, J., & Guo, L. (2023). Physiological and Biochemical Mechanisms of Exogenous Melatonin Regulation of Saline–Alkali Tolerance in Oats. Agronomy, 13(5), 1327. https://doi.org/10.3390/agronomy13051327