


Deciphering the Impact of Induced Drought in Agriculture Soils: Changes in Microbial Community Structure, Enzymatic and Metabolic Diversity

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
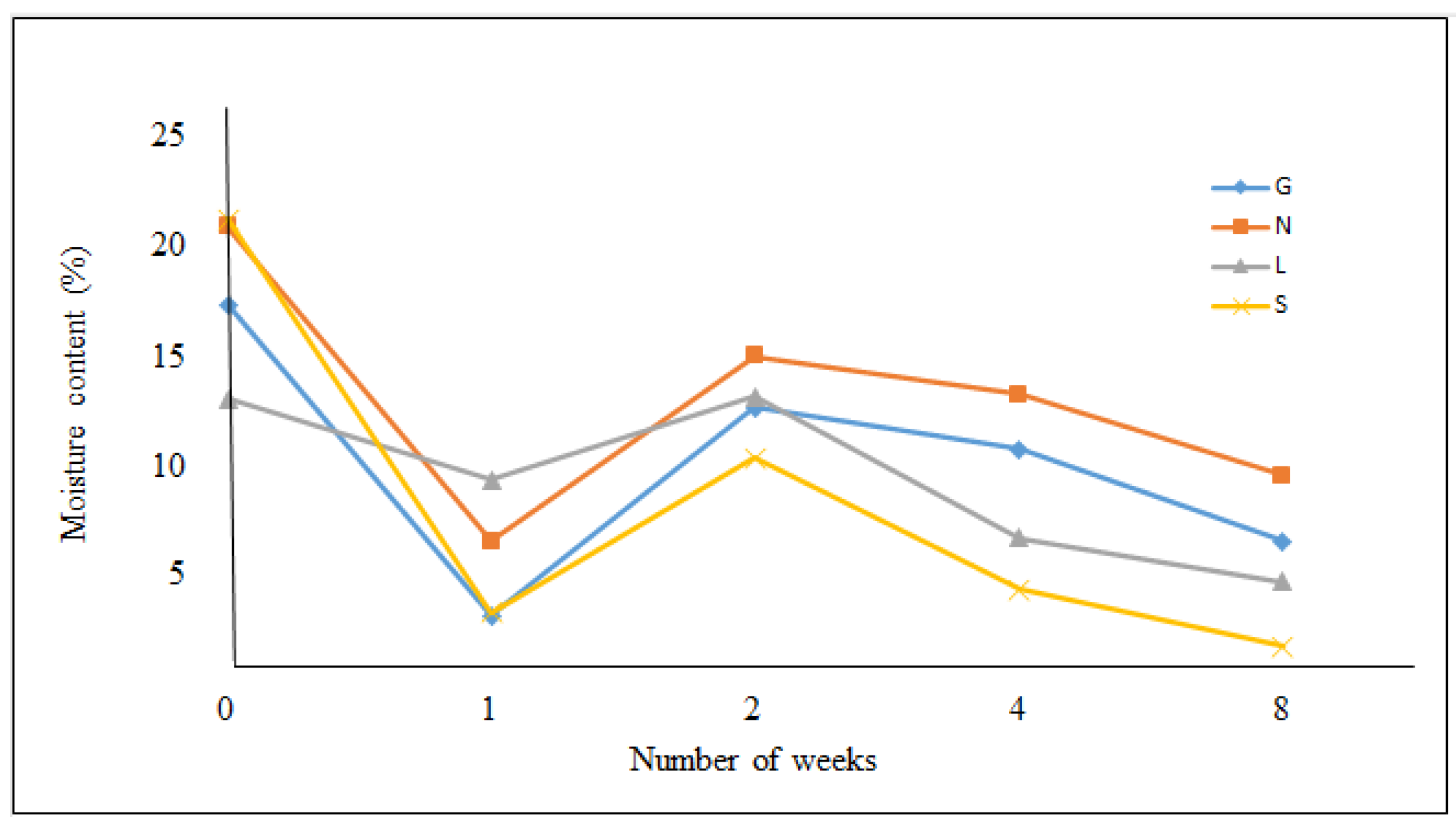
2.1. Sample Collection and Physicochemical Analyses of Soil Samples
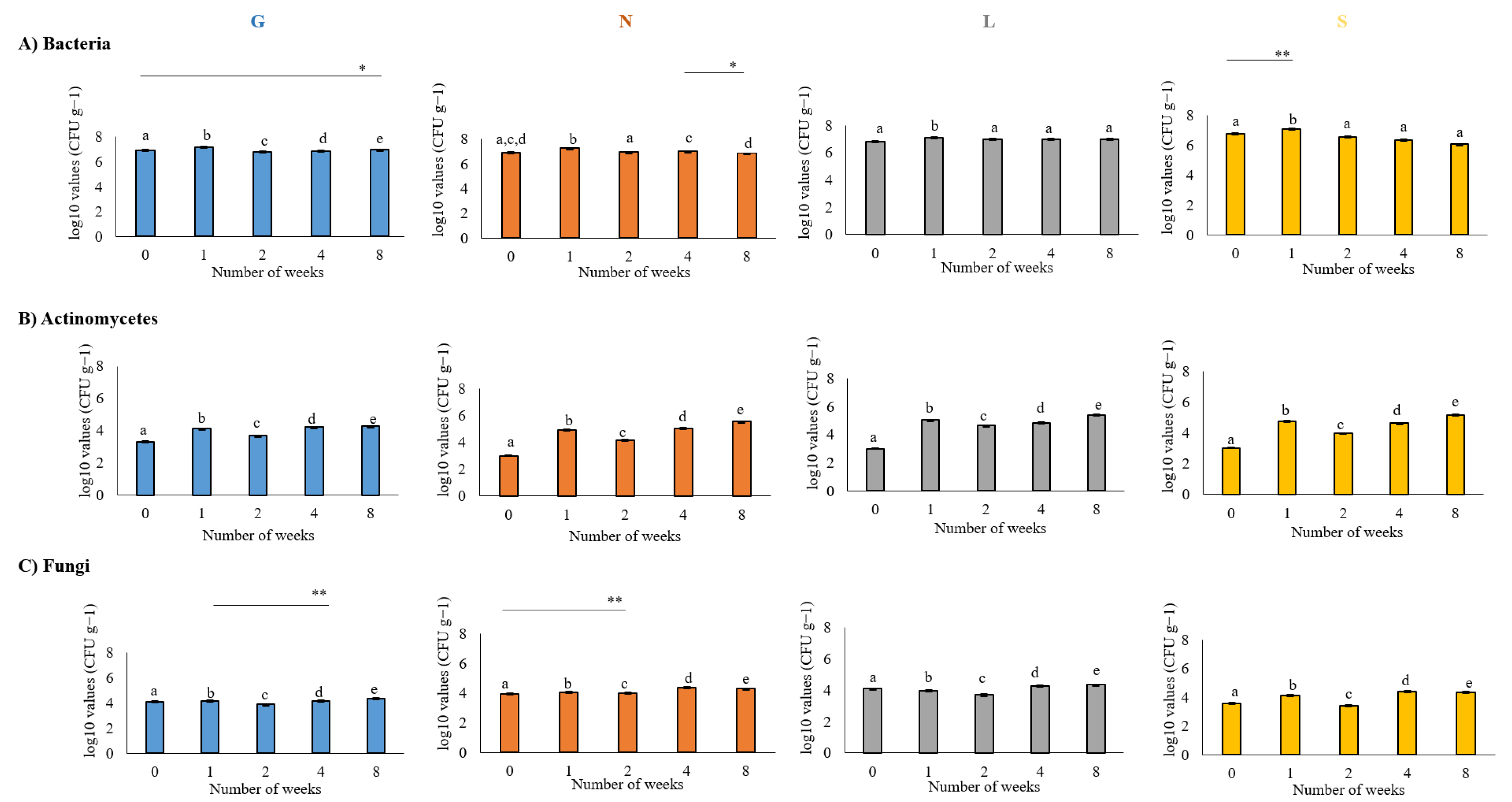
2.2. Determination of the Number of Bacteria, Actinomycetes and Fungi in Soil Samples
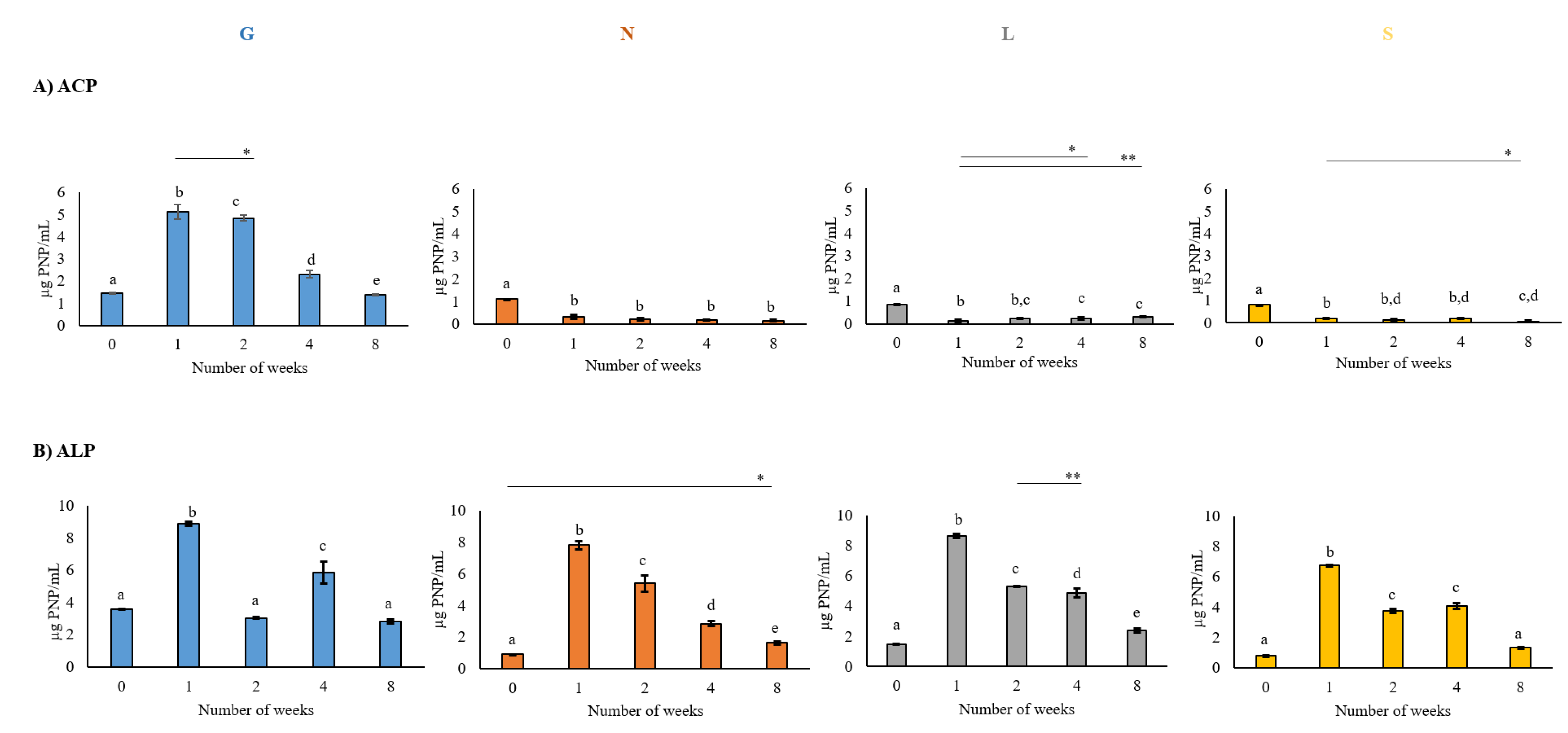
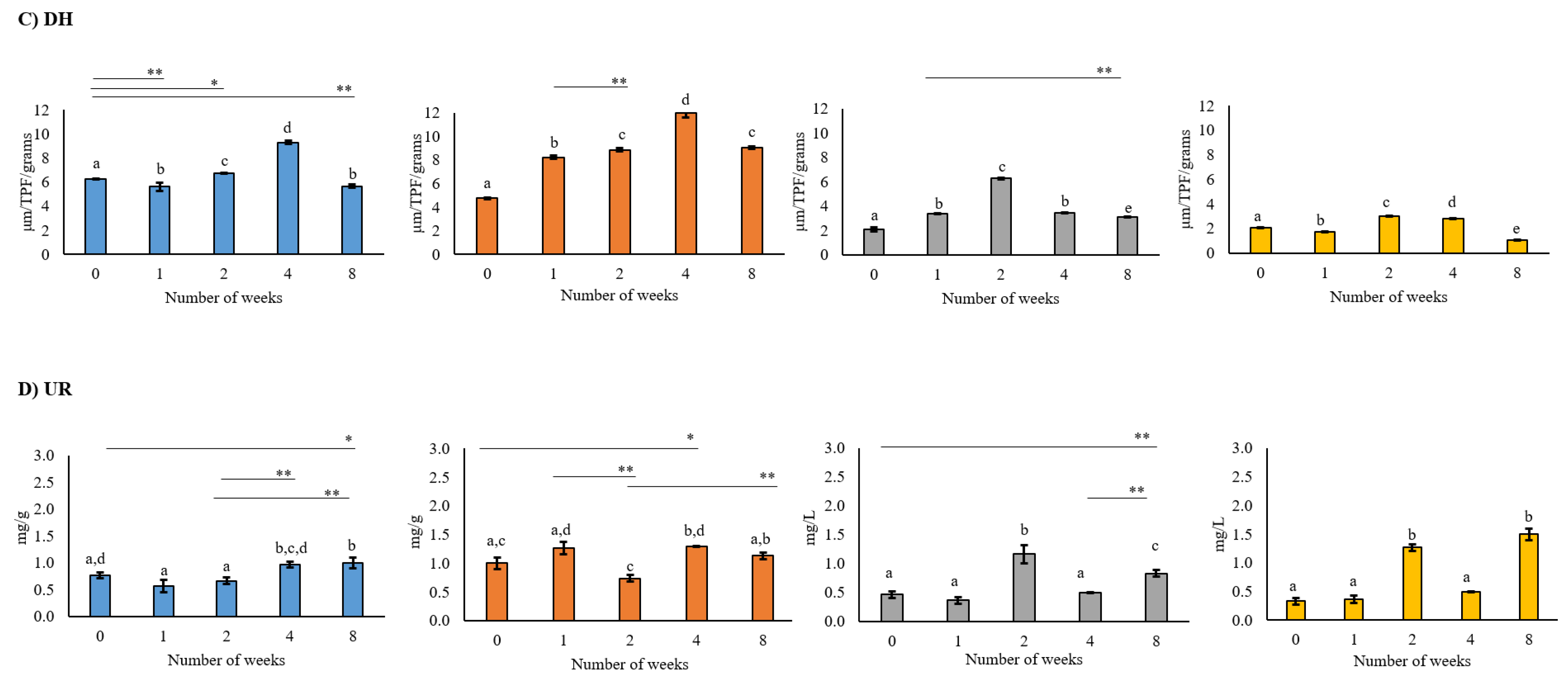
2.3. Soil Enzymatic Activities
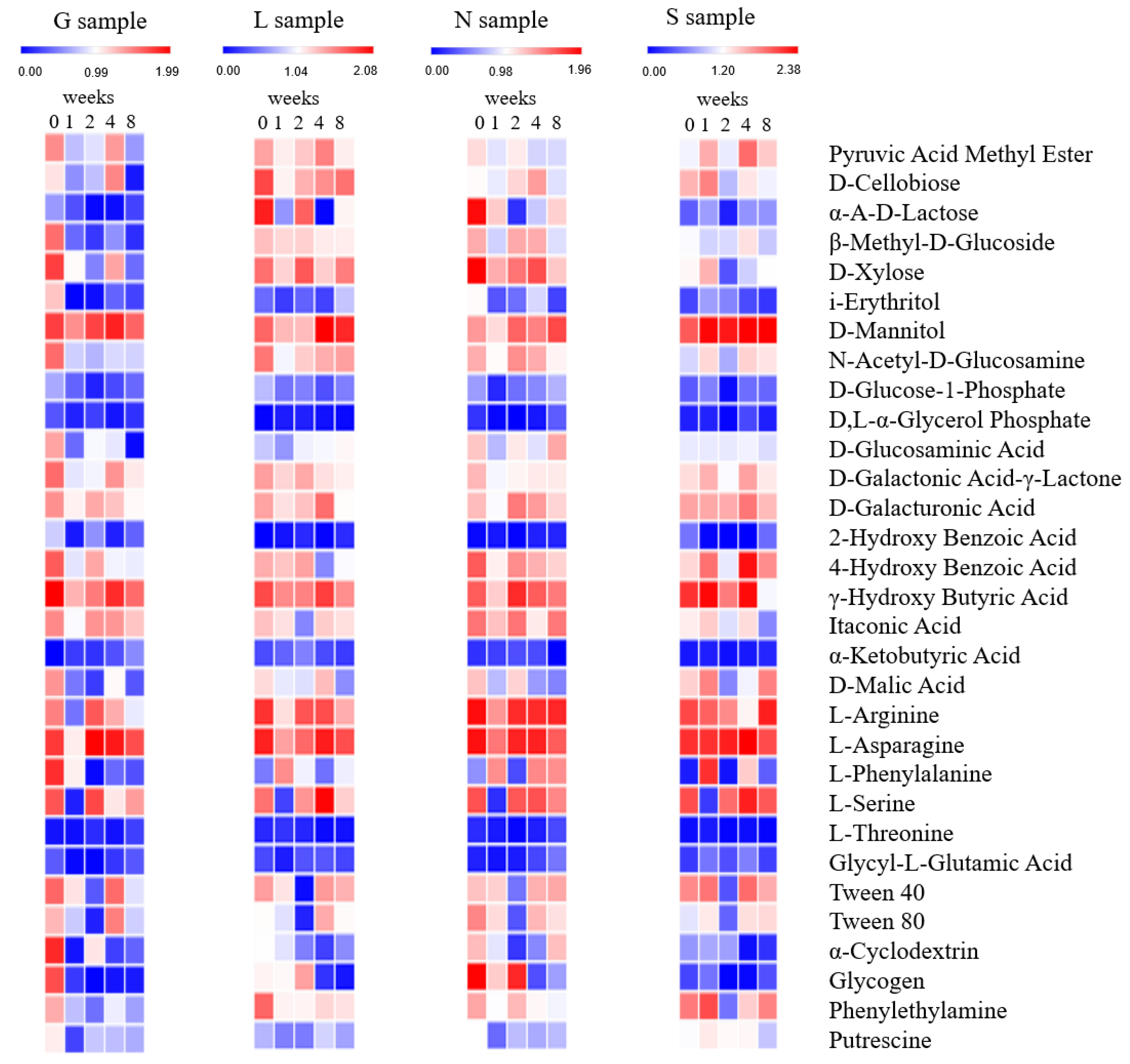
2.4. Metabolic Diversity of Soil Microbes
2.5. Statistical Analysis
2.6. DNA Extraction, Amplicon Sequencing and Bioinformatics Analyses
3. Results
3.1. Chemical Properties of Soil Samples
3.2. Influence of Prolonged Drought Stress on Number of Microorganisms
3.3. Effect of Drought Stress on Enzyme Activity
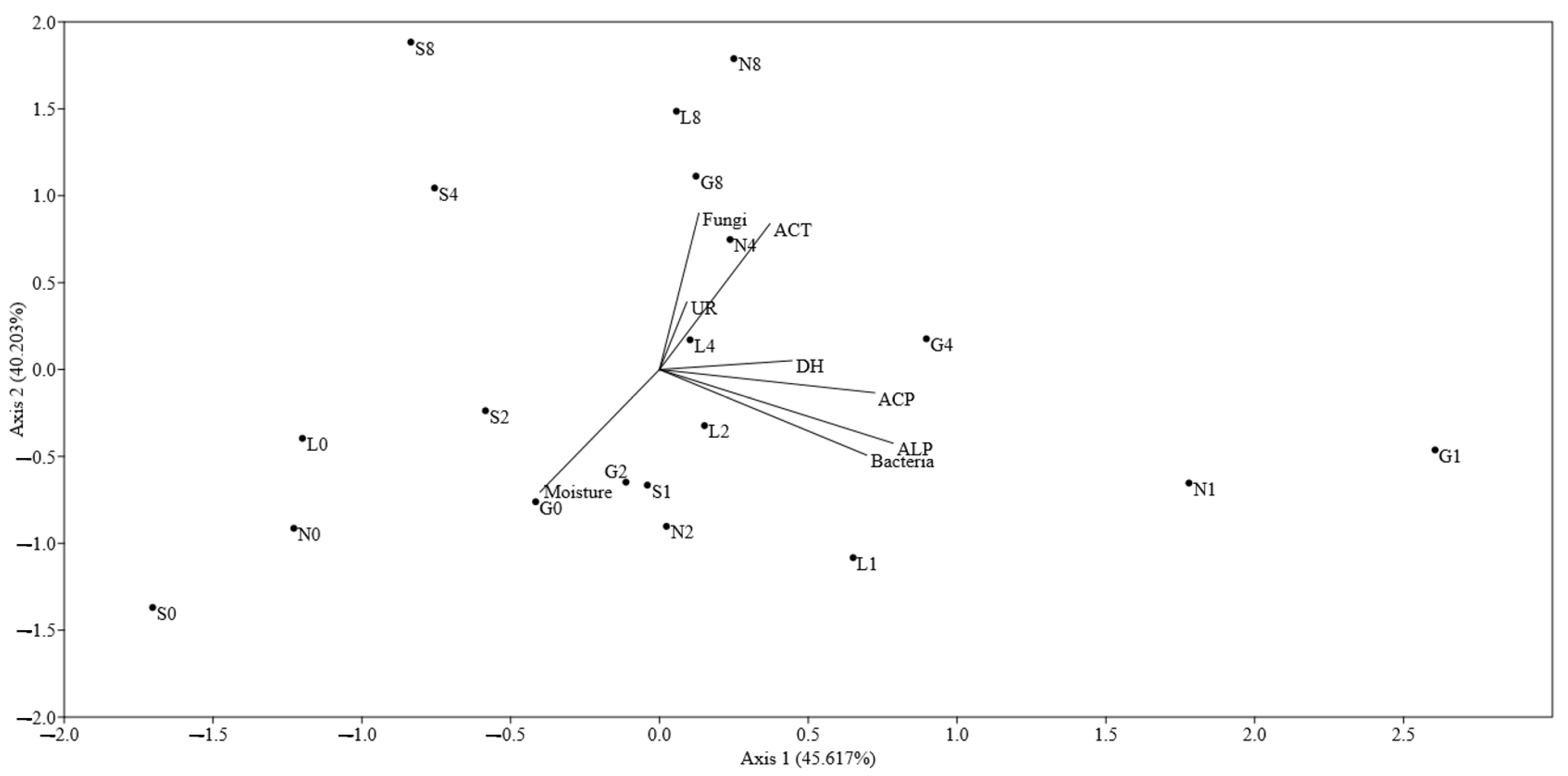
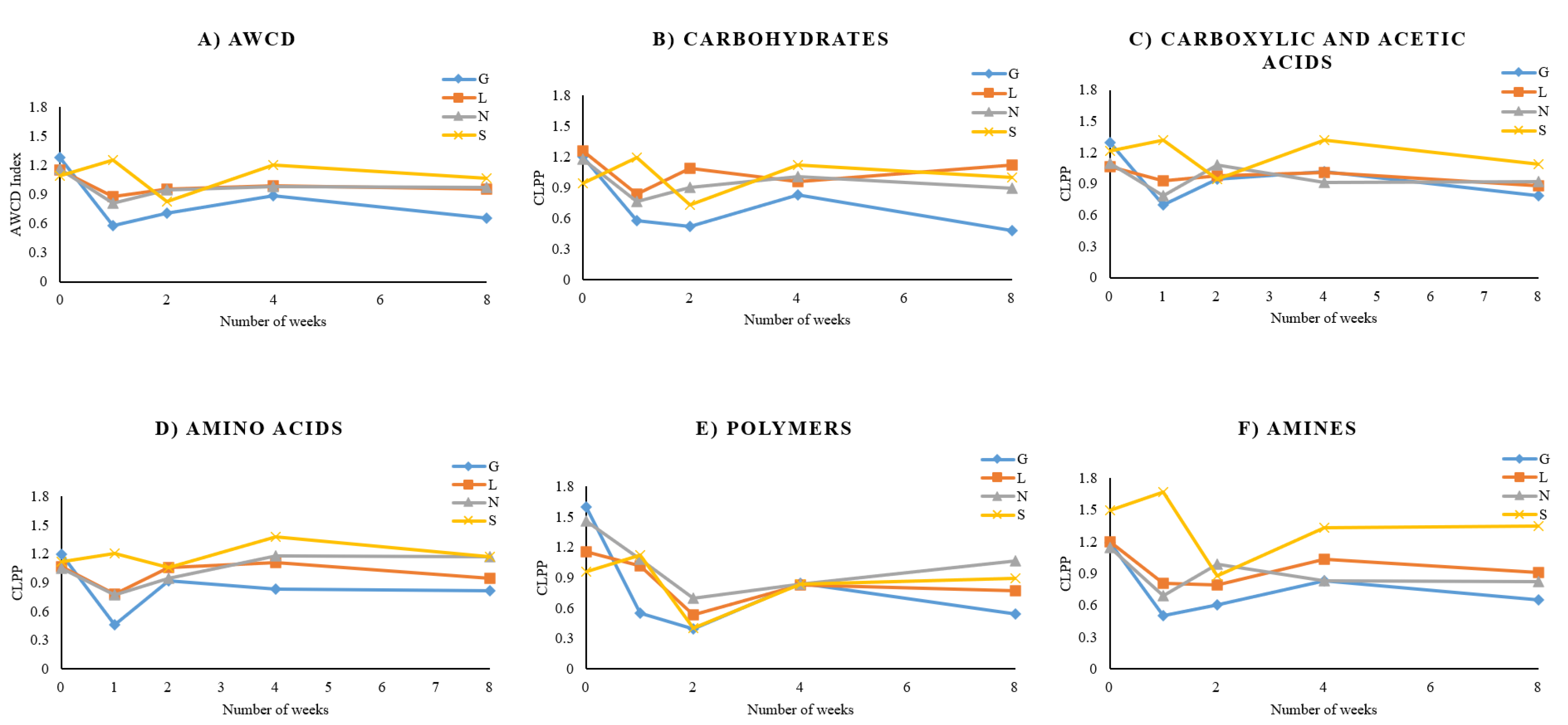
3.4. Estimation of Community Level Physiological Profiling (CLPP) of Soil Samples under Drought Stress
3.5. Impact of Drought on Genetic Diversity of Bacteria and Fungi
4. Discussion
4.1. Soil Chemical Parameters
4.2. Culture-Dependent and Culture-Independent Characterization of Microorganisms
4.3. Impact of Drought on Soil Enzyme Activities
4.4. CLPP Analysis under Drought Conditions
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumar, A.; Rana, K.S.; Choudhary, A.K.; Bana, R.S.; Sharma, V.K.; Prasad, S.; Gupta, G.; Choudhary, M.; Pradhan, A.; Rajpoot, S.K.; et al. Energy Budgeting and Carbon Footprints of Zero-Tilled Pigeonpea–Wheat Cropping System under Sole or Dual Crop Basis Residue Mulching and Zn-Fertilization in a Semi-Arid Agro-Ecology. Energy 2021, 231, 120862. [Google Scholar] [CrossRef]
- Swaminathan, M.S.; Kesavan, P.C. Agricultural Research in an Era of Climate Change. Agric. Res. 2012, 1, 3–11. [Google Scholar] [CrossRef]
- IPCC. Summary for Policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Delmotte, M.V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021; In Press. [Google Scholar]
- Bogati, K.; Walczak, M. The Impact of Drought Stress on Soil Microbial Community, Enzyme Activities and Plants. Agronomy 2022, 12, 189. [Google Scholar] [CrossRef]
- Jansson, J.K.; Hofmockel, K.S. Soil Microbiomes and Climate Change. Nat. Rev. Microbiol. 2020, 18, 35–46. [Google Scholar] [CrossRef]
- Siebielec, S.; Siebielec, G.; Klimkowicz-Pawlas, A.; Gałązka, A.; Grządziel, J.; Stuczyński, T. Impact of Water Stress on Microbial Community and Activity in Sandy and Loamy Soils. Agronomy 2020, 10, 1429. [Google Scholar] [CrossRef]
- Schimel, J.P.; Gulledge, J.M.; Clein-Curley, J.S.; Lindstrom, J.E.; Braddock, J.F. Moisture Effects on Microbial Activity and Community Structure in Decomposing Birch Litter in the Alaskan Taiga. Soil Biol. Biochem. 1999, 31, 831–838. [Google Scholar] [CrossRef]
- Santos-Medellín, C.; Edwards, J.; Liechty, Z.; Nguyen, B.; Sundaresan, V. Drought Stress Results in a Compartment-Specific Restructuring of the Rice Root-Associated Microbiomes. mBio 2017, 8, e00764-17. [Google Scholar] [CrossRef]
- Xu, L.; Naylor, D.; Dong, Z.; Simmons, T.; Pierroz, G.; Hixson, K.K.; Kim, Y.-M.; Zink, E.M.; Engbrecht, K.M.; Wang, Y.; et al. Drought Delays Development of the Sorghum Root Microbiome and Enriches for Monoderm Bacteria. Proc. Natl. Acad. Sci. USA 2018, 115, E4284–E4293. [Google Scholar] [CrossRef]
- Hayden, H.L.; Mele, P.M.; Bougoure, D.S.; Allan, C.Y.; Norng, S.; Piceno, Y.M.; Brodie, E.L.; DeSantis, T.Z.; Andersen, G.L.; Williams, A.L.; et al. Changes in the Microbial Community Structure of Bacteria, Archaea and Fungi in Response to Elevated CO2 and Warming in an Australian Native Grassland Soil: Climate Change Effects on Microbial Communities. Environ. Microbiol. 2012, 14, 3081–3096. [Google Scholar] [CrossRef]
- Oliveira, T.B.; Lucas, R.C.; de Scarcella, A.S.A.; Contato, A.G.; Pasin, T.M.; Martinez, C.A.; Polizeli, M.d.L.T.d.M. Fungal Communities Differentially Respond to Warming and Drought in Tropical Grassland Soil. Mol. Ecol. 2020, 29, 1550–1559. [Google Scholar] [CrossRef]
- Evans, S.E.; Wallenstein, M.D. Soil Microbial Community Response to Drying and Rewetting Stress: Does Historical Precipitation Regime Matter? Biogeochemistry 2012, 109, 101–116. [Google Scholar] [CrossRef]
- Fry, E.L.; Manning, P.; Macdonald, C.; Hasegawa, S.; De Palma, A.; Power, S.A.; Singh, B.K. Shifts in Microbial Communities Do Not Explain the Response of Grassland Ecosystem Function to Plant Functional Composition and Rainfall Change. Soil Biol. Biochem. 2016, 92, 199–210. [Google Scholar] [CrossRef]
- Wu, Z.; Dijkstra, P.; Koch, G.W.; Peñuelas, J.; Hungate, B.A. Responses of Terrestrial Ecosystems to Temperature and Precipitation Change: A Meta-Analysis of Experimental Manipulation: Meta-Analysis Of Experimental Manipulation. Glob. Chang. Biol. 2011, 17, 927–942. [Google Scholar] [CrossRef]
- Alster, C.J.; German, D.P.; Lu, Y.; Allison, S.D. Microbial Enzymatic Responses to Drought and to Nitrogen Addition in a Southern California Grassland. Soil Biol. Biochem. 2013, 64, 68–79. [Google Scholar] [CrossRef]
- Preece, C.; Farré-Armengol, G.; Peñuelas, J. Drought Is a Stronger Driver of Soil Respiration and Microbial Communities than Nitrogen or Phosphorus Addition in Two Mediterranean Tree Species. Sci. Total Environ. 2020, 735, 139554. [Google Scholar] [CrossRef] [PubMed]
- Steinweg, J.M.; Dukes, J.S.; Paul, E.A.; Wallenstein, M.D. Microbial Responses to Multi-Factor Climate Change: Effects on Soil Enzymes. Front. Microbiol. 2013, 4, 146. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Li, Y.; Wu, H.; Zhang, K.; Hao, Y.; Wang, J.; Zhang, X.; Yan, L.; Kang, X. Different Responses of Soil Hydrolases and Oxidases to Extreme Drought in an Alpine Peatland on the Qinghai-Tibet Plateau, China. Eur. J. Soil Biol. 2020, 99, 103195. [Google Scholar] [CrossRef]
- Deng, L.; Peng, C.; Kim, D.-G.; Li, J.; Liu, Y.; Hai, X.; Liu, Q.; Huang, C.; Shangguan, Z.; Kuzyakov, Y. Drought Effects on Soil Carbon and Nitrogen Dynamics in Global Natural Ecosystems. Earth-Sci. Rev. 2021, 214, 103501. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Drought Decreases Soil Enzyme Activity in a Mediterranean Quercus Ilex L. Forest. Soil Biol. Biochem. 2005, 37, 455–461. [Google Scholar] [CrossRef]
- Wolińska, A.; Stepniewsk, Z. Dehydrogenase Activity in the Soil Environment. In Dehydrogenases; Canuto, R.A., Ed.; InTech: Houston, TX, USA, 2012; ISBN 978-953-307-019-3. [Google Scholar]
- Henry, H.A.L. Reprint of “Soil Extracellular Enzyme Dynamics in a Changing Climate”. Soil Biol. Biochem. 2013, 56, 53–59. [Google Scholar] [CrossRef]
- Warzyński, H.; Sosnowska, A.; Harasimiuk, A. Effect of Variable Content of Organic Matter and Carbonates on Results of Determination of Granulometric Composition by Means of Casagrande’s Areometric Method in Modification by Prószyński. Soil Sci. Annu. 2018, 69, 39–48. [Google Scholar] [CrossRef]
- Bednarek, R.; Dziadowiec, H.; Pokojska, U.; Prusinkiewicz, Z. Eco-Pedological Studies; PWN: Warsaw, Poland, 2004; p. 344. [Google Scholar]
- Furtak, K.; Grządziel, J.; Gałązka, A.; Niedźwiecki, J. Analysis of Soil Properties, Bacterial Community Composition, and Metabolic Diversity in Fluvisols of a Floodplain Area. Sustainability 2019, 11, 3929. [Google Scholar] [CrossRef]
- Nakano, H.; Takenishi, S.; Watanabe, Y. Purification and Properties of Urease from Brevibacterium Ammoniagenes. Agric. Biol. Chem. 1984, 48, 1495–1502. [Google Scholar] [CrossRef]
- Kandeler, E.; Gerber, H. Short-Term Assay of Soil Urease Activity Using Colorimetric Determination of Ammonium. Biol. Fert. Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Tabatabai, M.A. Soil enzymes. In Methods of Soil Analysis, Part 2; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy and Soil Science Society of America: Madison, WI, USA, 1982. [Google Scholar]
- Weber, K.P.; Legge, R.L. One-Dimensional Metric for Tracking Bacterial Community Divergence Using Sole Carbon Source Utilization Patterns. J. Microbiol. Methods 2009, 79, 55–61. [Google Scholar] [CrossRef]
- Garland, J.L.; Mills, A.L. Classification and Characterization of Heterotrophic Microbial Communities on the Basis of Patterns of Community-Level Sole-Carbon-Source Utilization. Appl. Environ. Microbiol. 1991, 57, 2351–2359. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S Ribosomal DNA Amplification for Phylogenetic Study. J Bacteriol 1991, 173, 697–703. [Google Scholar] [CrossRef]
- Leser, T.D.; Amenuvor, J.Z.; Jensen, T.K.; Lindecrona, R.H.; Boye, M.; Møller, K. Culture-Independent Analysis of Gut Bacteria: The Pig Gastrointestinal Tract Microbiota Revisited. Appl. Environ. Microbiol. 2002, 68, 673–690. [Google Scholar] [CrossRef]
- Turner, S.; Pryer, K.M.; Miao, V.P.W.; Palmer, J.D. Investigating Deep Phylogenetic Relationships among Cyanobacteria and Plastids by Small Subunit RRNA Sequence Analysis. J. Eukaryot. Microbiol. 1999, 46, 327–338. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal Rna Genes for Phylogenetics. In PCR Protocols; Elsevier: Amsterdam, The Netherlands, 1990; pp. 315–322. ISBN 978-0-12-372180-8. [Google Scholar]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Shen, W.; Le, S.; Li, Y.; Hu, F. SeqKit: A Cross-Platform and Ultrafast Toolkit for FASTA/Q File Manipulation. PLoS ONE 2016, 11, e0163962. [Google Scholar] [CrossRef] [PubMed]
- Truong, D.T.; Franzosa, E.A.; Tickle, T.L.; Scholz, M.; Weingart, G.; Pasolli, E.; Tett, A.; Huttenhower, C.; Segata, N. MetaPhlAn2 for Enhanced Metagenomic Taxonomic Profiling. Nat. Methods 2015, 12, 902–903. [Google Scholar] [CrossRef] [PubMed]
- Breitwieser, F.P.; Baker, D.N.; Salzberg, S.L. KrakenUniq: Confident and Fast Metagenomics Classification Using Unique k-Mer Counts. Genome Biol. 2018, 19, 198. [Google Scholar] [CrossRef] [PubMed]
- Wood, D.E.; Salzberg, S.L. Kraken: Ultrafast Metagenomic Sequence Classification Using Exact Alignments. Genome Biol. 2014, 15, R46. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.2-0. 2014. Available online: http://CRAN.Rproject.org/package=vegan (accessed on 10 April 2023).
- Nguyen, L.T.T.; Osanai, Y.; Anderson, I.C.; Bange, M.P.; Tissue, D.T.; Singh, B.K. Flooding and Prolonged Drought Have Differential Legacy Impacts on Soil Nitrogen Cycling, Microbial Communities and Plant Productivity. Plant Soil 2018, 431, 371–387. [Google Scholar] [CrossRef]
- Schimel, J.P. Life in Dry Soils: Effects of Drought on Soil Microbial Communities and Processes. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 409–432. [Google Scholar] [CrossRef]
- Zhang, Q.; Shao, M.; Jia, X.; Wei, X. Changes in Soil Physical and Chemical Properties after Short Drought Stress in Semi-Humid Forests. Geoderma 2019, 338, 170–177. [Google Scholar] [CrossRef]
- Luis Moreno, J.; Bastida, F.; Díaz-López, M.; Li, Y.; Zhou, Y.; López-Mondéjar, R.; Benavente-Ferraces, I.; Rojas, R.; Rey, A.; Carlos García-Gil, J.; et al. Response of Soil Chemical Properties, Enzyme Activities and Microbial Communities to Biochar Application and Climate Change in a Mediterranean Agroecosystem. Geoderma 2022, 407, 115536. [Google Scholar] [CrossRef]
- Chodak, M.; Gołębiewski, M.; Morawska-Płoskonka, J.; Kuduk, K.; Niklińska, M. Soil Chemical Properties Affect the Reaction of Forest Soil Bacteria to Drought and Rewetting Stress. Ann. Microbiol. 2015, 65, 1627–1637. [Google Scholar] [CrossRef]
- Hartmann, M.; Brunner, I.; Hagedorn, F.; Bardgett, R.D.; Stierli, B.; Herzog, C.; Chen, X.; Zingg, A.; Graf-Pannatier, E.; Rigling, A.; et al. A Decade of Irrigation Transforms the Soil Microbiome of a Semi-Arid Pine Forest. Mol. Ecol. 2017, 26, 1190–1206. [Google Scholar] [CrossRef]
- Curiel Yuste, J.; Fernandez-Gonzalez, A.J.; Fernandez-Lopez, M.; Ogaya, R.; Penuelas, J.; Sardans, J.; Lloret, F. Strong Functional Stability of Soil Microbial Communities under Semiarid Mediterranean Conditions and Subjected to Long-Term Shifts in Baseline Precipitation. Soil Biol. Biochem. 2014, 69, 223–233. [Google Scholar] [CrossRef]
- Mickan, B.S.; Abbott, L.K.; Solaiman, Z.M.; Mathes, F.; Siddique, K.H.M.; Jenkins, S.N. Soil Disturbance and Water Stress Interact to Influence Arbuscular Mycorrhizal Fungi, Rhizosphere Bacteria and Potential for N and C Cycling in an Agricultural Soil. Biol. Fertil. Soils 2019, 55, 53–66. [Google Scholar] [CrossRef]
- Naylor, D.; Coleman-Derr, D. Drought Stress and Root-Associated Bacterial Communities. Front. Plant Sci. 2018, 8, 2223. [Google Scholar] [CrossRef]
- Veach, A.M.; Zeglin, L.H. Historical Drought Affects Microbial Population Dynamics and Activity During Soil Drying and Re-Wet. Microb. Ecol. 2020, 79, 662–674. [Google Scholar] [CrossRef]
- Mohammadipanah, F.; Wink, J. Actinobacteria from Arid and Desert Habitats: Diversity and Biological Activity. Front. Microbiol. 2016, 6, 1541. [Google Scholar] [CrossRef]
- Bouskill, N.J.; Wood, T.E.; Baran, R.; Ye, Z.; Bowen, B.P.; Lim, H.; Zhou, J.; Nostrand, J.D.V.; Nico, P.; Northen, T.R.; et al. Belowground Response to Drought in a Tropical Forest Soil. I. Changes in Microbial Functional Potential and Metabolism. Front. Microbiol. 2016, 7, 525. [Google Scholar] [CrossRef]
- Hawkes, C.V.; Kivlin, S.N.; Rocca, J.D.; Huguet, V.; Thomsen, M.A.; Suttle, K.B. Fungal Community Responses to Precipitation: Fungal Climate Response. Glob. Chang. Biol. 2011, 17, 1637–1645. [Google Scholar] [CrossRef]
- Fahey, C.; Koyama, A.; Antunes, P.M.; Dunfield, K.; Flory, S.L. Plant Communities Mediate the Interactive Effects of Invasion and Drought on Soil Microbial Communities. ISME J. 2020, 14, 1396–1409. [Google Scholar] [CrossRef]
- Balasooriya, W.K.; Denef, K.; Huygens, D.; Boeckx, P. Translocation and Turnover of Rhizodeposit Carbon within Soil Microbial Communities of an Extensive Grassland Ecosystem. Plant Soil 2014, 376, 61–73. [Google Scholar] [CrossRef]
- Thaysen, E.M.; Reinsch, S.; Larsen, K.S.; Ambus, P. Decrease in Heathland Soil Labile Organic Carbon under Future Atmospheric and Climatic Conditions. Biogeochemistry 2017, 133, 17–36. [Google Scholar] [CrossRef]
- Zornoza, R.; Guerrero, C.; Mataix-Solera, J.; Arcenegui, V.; García-Orenes, F.; Mataix-Beneyto, J. Assessing the Effects of Air-Drying and Rewetting Pre-Treatment on Soil Microbial Biomass, Basal Respiration, Metabolic Quotient and Soluble Carbon under Mediterranean Conditions. Eur. J. Soil Biol. 2007, 43, 120–129. [Google Scholar] [CrossRef]
- Bowles, T.M.; Acosta-Martínez, V.; Calderón, F.; Jackson, L.E. Soil Enzyme Activities, Microbial Communities, and Carbon and Nitrogen Availability in Organic Agroecosystems across an Intensively-Managed Agricultural Landscape. Soil Biol. Biochem. 2014, 68, 252–262. [Google Scholar] [CrossRef]
- Carbone, M.J.; Alaniz, S.; Mondino, P.; Gelabert, M.; Eichmeier, A.; Tekielska, D.; Bujanda, R.; Gramaje, D. Drought Influences Fungal Community Dynamics in the Grapevine Rhizosphere and Root Microbiome. J. Fungi 2021, 7, 686. [Google Scholar] [CrossRef] [PubMed]
- Hanaka, A.; Ozimek, E.; Reszczyńska, E.; Jaroszuk-Ściseł, J.; Stolarz, M. Plant Tolerance to Drought Stress in the Presence of Supporting Bacteria and Fungi: An Efficient Strategy in Horticulture. Horticulturae 2021, 7, 390. [Google Scholar] [CrossRef]
- Yan, L.; Zhu, J.; Zhao, X.; Shi, J.; Jiang, C.; Shao, D. Beneficial Effects of Endophytic Fungi Colonization on Plants. Appl. Microbiol. Biotechnol. 2019, 103, 3327–3340. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Hueso, R.; Collins, S.L.; Delgado-Baquerizo, M.; Hamonts, K.; Pockman, W.T.; Sinsabaugh, R.L.; Smith, M.D.; Knapp, A.K.; Power, S.A. Drought Consistently Alters the Composition of Soil Fungal and Bacterial Communities in Grasslands from Two Continents. Glob. Chang. Biol. 2018, 24, 2818–2827. [Google Scholar] [CrossRef]
- Manzoni, S.; Schimel, J.P.; Porporato, A. Responses of Soil Microbial Communities to Water Stress: Results from a Meta-analysis. Ecology 2012, 93, 930–938. [Google Scholar] [CrossRef]
- Chenu, C.; Hassink, J.; Bloem, J. Short-term changes in the spatial distribution of microorganisms in soil aggregates as affected by glucose addition. Biol. Fertil. Soils 2001, 34, 349–356. [Google Scholar] [CrossRef]
- Lennon, J.T.; Jones, S.E. Microbial Seed Banks: The Ecological and Evolutionary Implications of Dormancy. Nat. Rev. Microbiol. 2011, 9, 119–130. [Google Scholar] [CrossRef]
- Maestre, F.T.; Delgado-Baquerizo, M.; Jeffries, T.C.; Eldridge, D.J.; Ochoa, V.; Gozalo, B.; Quero, J.L.; García-Gómez, M.; Gallardo, A.; Ulrich, W.; et al. Increasing Aridity Reduces Soil Microbial Diversity and Abundance in Global Drylands. Proc. Natl. Acad. Sci. USA 2015, 112, 15684–15689. [Google Scholar] [CrossRef]
- Vargas-Gastélum, L.; Romero-Olivares, A.L.; Escalante, A.E.; Rocha-Olivares, A.; Brizuela, C.; Riquelme, M. Impact of Seasonal Changes on Fungal Diversity of a Semi-Arid Ecosystem Revealed by 454 Pyrosequencing. FEMS Microbiol. Ecol. 2015, 91, fiv044. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, L.; Chen, Q.; Wen, X.; Liao, Y. Conservation Tillage Increases Soil Bacterial Diversity in the Dryland of Northern China. Agron. Sustain. Dev. 2016, 36, 28. [Google Scholar] [CrossRef]
- Joergensen, R.; Wichern, F. Quantitative Assessment of the Fungal Contribution to Microbial Tissue in Soil. Soil Biol. Biochem. 2008, 40, 2977–2991. [Google Scholar] [CrossRef]
- Kaisermann, A.; Maron, P.A.; Beaumelle, L.; Lata, J.C. Fungal Communities Are More Sensitive Indicators to Non-Extreme Soil Moisture Variations than Bacterial Communities. Appl. Soil Ecol. 2015, 86, 158–164. [Google Scholar] [CrossRef]
- Allison, S.D.; Treseder, K.K. Warming and Drying Suppress Microbial Activity and Carbon Cycling in Boreal Forest Soils: Warming Suppresses Microbial Activity. Glob. Chang. Biol. 2008, 14, 2898–2909. [Google Scholar] [CrossRef]
- Li, X.; Sarah, P. Enzyme Activities along a Climatic Transect in the Judean Desert. CATENA 2003, 53, 349–363. [Google Scholar] [CrossRef]
- Williams, M.A.; Rice, C.W. Seven Years of Enhanced Water Availability Influences the Physiological, Structural, and Functional Attributes of a Soil Microbial Community. Appl. Soil Ecol. 2007, 35, 535–545. [Google Scholar] [CrossRef]
- Nannipieri, P.; Giagnoni, L.; Landi, L.; Renella, G. Role of Phosphatase Enzymes in Soil. In Phosphorus in Action; Bünemann, E., Oberson, A., Frossard, E., Eds.; Soil Biology; Springer: Berlin/Heidelberg, Germany, 2011; Volume 26, pp. 215–243. ISBN 978-3-642-15270-2. [Google Scholar]
- Dodor, D.E.; Tabatabai, M.A. Effect of Cropping Systems on Phosphatases in Soils. Z. Pflanzenernähr. Bodenk. 2003, 166, 7–13. [Google Scholar] [CrossRef]
- Pukalchik, M.; Mercl, F.; Terekhova, V.; Tlustoš, P. Biochar, Wood Ash and Humic Substances Mitigating Trace Elements Stress in Contaminated Sandy Loam Soil: Evidence from an Integrative Approach. Chemosphere 2018, 203, 228–238. [Google Scholar] [CrossRef]
- Song, F.; Han, X.; Zhu, X.; Herbert, S.J. Response to Water Stress of Soil Enzymes and Root Exudates from Drought and Non-Drought Tolerant Corn Hybrids at Different Growth Stages. Can. J. Soil. Sci. 2012, 92, 501–507. [Google Scholar] [CrossRef]
- Huang, W.; Liu, J.; Zhou, G.; Zhang, D.; Deng, Q. Effects of Precipitation on Soil Acid Phosphatase Activity in Three Successional Forests in Southern China. Biogeosciences 2011, 8, 1901–1910. [Google Scholar] [CrossRef]
- Suriyagoda, L.D.B.; Ryan, M.H.; Renton, M.; Lambers, H. Plant Responses to Limited Moisture and Phosphorus Availability. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2014; Volume 124, pp. 143–200. ISBN 978-0-12-800138-7. [Google Scholar]
- Menge, D.N.L.; Field, C.B. Simulated Global Changes Alter Phosphorus Demand in Annual Grassland. Glob. Chang. Biol. 2007, 13, 2582–2591. [Google Scholar] [CrossRef]
- Piotrowska-Długosz, A.; Charzyński, P. The Impact of the Soil Sealing Degree on Microbial Biomass, Enzymatic Activity, and Physicochemical Properties in the Ekranic Technosols of Toruń (Poland). J. Soils Sediments 2015, 15, 47–59. [Google Scholar] [CrossRef]
- Ng, E.-L.; Patti, A.F.; Rose, M.T.; Schefe, C.R.; Smernik, R.J.; Cavagnaro, T.R. Do Organic Inputs Alter Resistance and Resilience of Soil Microbial Community to Drying? Soil Biol. Biochem. 2015, 81, 58–66. [Google Scholar] [CrossRef]
- Burns, R.G.; DeForest, J.L.; Marxsen, J.; Sinsabaugh, R.L.; Stromberger, M.E.; Wallenstein, M.D.; Weintraub, M.N.; Zoppini, A. Soil Enzymes in a Changing Environment: Current Knowledge and Future Directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar] [CrossRef]
- Ros, M.; Goberna, M.; Moreno, J.L.; Hernandez, T.; García, C.; Insam, H.; Pascual, J.A. Molecular and Physiological Bacterial Diversity of a Semi-Arid Soil Contaminated with Different Levels of Formulated Atrazine. Appl. Soil Ecol. 2006, 34, 93–102. [Google Scholar] [CrossRef]
- Chakraborty, A.; Chakrabarti, K.; Chakraborty, A.; Ghosh, S. Effect of Long-Term Fertilizers and Manure Application on Microbial Biomass and Microbial Activity of a Tropical Agricultural Soil. Biol. Fertil. Soils 2011, 47, 227–233. [Google Scholar] [CrossRef]
- Braun, B.; Böckelmann, U.; Grohmann, E.; Szewzyk, U. Bacterial Soil Communities Affected by Water-Repellency. Geoderma 2010, 158, 343–351. [Google Scholar] [CrossRef]
- Misson, L.; Rocheteau, A.; Rambal, S.; Ourcival, J.-M.; Limousin, J.-M.; Rodriguez, R. Functional Changes in the Control of Carbon Fluxes after 3 Years of Increased Drought in a Mediterranean Evergreen Forest? Glob. Chang. Biol. 2009, 16, 2461–2475. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Abbreviation | Total Organic Carbon Content (%) (Mean ± SD) | Total Nitrogen Content (%) (Mean ± SD) | pH (Mean ± SD) | |||
|---|---|---|---|---|---|---|---|
| T0 | T8 | T0 | T8 | T0 | T8 | ||
| 1. Gniewkowo | G | 0.97 ± 0.010 | 0.89 ± 0.010 b | 0.112 ± 0.002 | 0.110 ± 0.002 | 6.96 ± 0.006 | 7.49 ± 0.01 c |
| 2. Lulkowo | L | 0.77 ± 0.006 | 0.80 ± 0.012 | 0.092 ± 0.001 | 0.087 ± 0.002 a | 6.52 ± 0.012 | 6.70 ± 0.006 c |
| 3. Wielka Nieszawka | N | 1.10 ± 0.015 | 0.99 ± 0.511 | 0.140 ± 0.001 | 0.135 ± 0.001 a | 6.69 ± 0.01 | 6.66 ± 0.01 |
| 4. Suchatówka | S | 0.62 ± 0.010 | 0.60 ± 0.015 | 0.057 ± 0.001 | 0.052 ± 0.001 a | 6.17 ± 0.015 | 5.77 ± 0.012 b |
| Taxon | Relative Abundance (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| G0 | G8 | L0 | L8 | N0 | N8 | S0 | S8 | ||
| A. Bacterial Phyla | Pseudomonadota | 57.65 | 44.91 | 63.84 | 46.70 | 59.08 | 52.38 | 60.83 | 48.37 |
| Actinomycetota | 38.38 | 53.36 | 32.83 | 51.09 | 37.57 | 45.65 | 36.96 | 49.40 | |
| Bacteroidota | 3.14 | 0.61 | 2.47 | 0.97 | 2.15 | 0.93 | 1.14 | 1.10 | |
| Bacillota | 0.83 | 1.13 | 0.85 | 1.24 | 1.21 | 1.05 | 1.06 | 1.12 | |
| Genera | Bradyrhizobium | 26.70 | 28.16 | 29.93 | 33.61 | 48.51 | 46.87 | 36.97 | 24.63 |
| Streptomyces | 17.33 | 19.38 | 12.18 | 20.66 | 12.25 | 16.56 | 12.14 | 17.42 | |
| Sphingomonas | 16.06 | 3.82 | 19.81 | 3.68 | 9.73 | 4.11 | 16.62 | 5.45 | |
| Nocardioides | 8.53 | 13.66 | 8.59 | 11.24 | 5.99 | 7.42 | 9.26 | 12.17 | |
| Mycobacterium | 6.22 | 8.22 | 7.16 | 8.91 | 6.73 | 8.01 | 4.42 | 15.46 | |
| Micromonospora | 4.44 | 7.07 | 2.72 | 4.28 | 3.03 | 3.75 | 3.58 | 4.56 | |
| Lysobacter | 6.67 | 2.18 | 7.48 | 1.74 | 5.01 | 1.74 | 5.38 | 2.58 | |
| Solirubrobacter | 3.11 | 4.96 | 2.82 | 4.38 | 3.18 | 4.07 | 2.69 | 3.44 | |
| Actinoplanes | 4.14 | 4.16 | 2.84 | 4.52 | 2.12 | 2.65 | 3.30 | 4.73 | |
| Geodermatophilus | 3.99 | 5.94 | 2.98 | 2.98 | 1.78 | 2.06 | 3.16 | 3.57 | |
| Sorangium | 2.80 | 2.44 | 3.49 | 4.01 | 1.67 | 2.77 | 2.47 | 6.00 | |
| B. Fungal Phyla | Ascomycota | 83.48 | 82.58 | 83.46 | 83.22 | 83.27 | 85.23 | 81.54 | 83.76 |
| Basidiomycota | 10.25 | 12.52 | 11.34 | 10.91 | 10.86 | 10.11 | 13.51 | 11.64 | |
| Mucoromycota | 3.57 | 2.82 | 2.79 | 3.95 | 3.31 | 2.18 | 2.11 | 2.86 | |
| Apicomplexa | 2.70 | 2.08 | 2.41 | 1.92 | 2.56 | 2.49 | 2.84 | 1.73 | |
| Genera | Fusarium | 22.88 | 14.49 | 17.28 | 29.15 | 21.39 | 26.07 | 19.79 | 40.10 |
| Aspergillus | 15.35 | 16.03 | 19.57 | 13.75 | 16.43 | 15.12 | 16.26 | 12.66 | |
| Colletotrichum | 5.99 | 8.61 | 5.76 | 6.16 | 7.36 | 5.76 | 8.39 | 5.02 | |
| Trichoderma | 3.88 | 4.85 | 5.19 | 5.02 | 14.09 | 12.49 | 3.37 | 2.40 | |
| Penicillium | 2.80 | 8.27 | 2.28 | 3.82 | 4.68 | 4.91 | 4.56 | 2.34 | |
| Exophiala | 4.73 | 4.85 | 2.68 | 6.67 | 2.74 | 3.99 | 3.65 | 3.14 | |
| Ustilago | 1.54 | 1.94 | 5.31 | 4.56 | 3.14 | 1.08 | 8.16 | 5.36 | |
| Pseudogymnoascus | 4.28 | 6.85 | 3.31 | 1.60 | 4.51 | 5.08 | 3.02 | 1.03 | |
| Verticillium | 3.42 | 2.62 | 11.87 | 3.02 | 2.05 | 1.43 | 2.85 | 1.94 | |
| Chaetomium | 7.24 | 2.68 | 1.60 | 1.31 | 1.65 | 2.00 | 2.80 | 1.43 | |
| Lobosporangium | 3.31 | 2.97 | 2.17 | 1.83 | 2.85 | 2.40 | 1.77 | 1.43 | |
| Marssonina | 2.17 | 3.54 | 2.34 | 1.88 | 2.28 | 2.00 | 2.91 | 0.80 | |
| Metarhizium | 1.25 | 3.02 | 1.48 | 1.77 | 1.54 | 1.71 | 5.19 | 1.48 | |
| Anthracocystis | 2.62 | 3.31 | 2.17 | 1.77 | 2.11 | 1.43 | 1.48 | 1.71 | |
| Thermothelomyces | 2.34 | 2.62 | 1.88 | 1.43 | 1.14 | 1.25 | 2.51 | 1.71 | |
| Gaeumannomyces | 1.71 | 1.83 | 1.77 | 1.65 | 1.08 | 1.48 | 1.83 | 3.08 | |
| Rhizophagus | 1.94 | 0.97 | 1.83 | 4.28 | 1.31 | 1.03 | 1.54 | 1.43 | |
| Phycomyces | 1.43 | 2.34 | 1.83 | 2.11 | 2.34 | 1.31 | 1.43 | 1.43 | |
| Tilletiopsis | 1.77 | 2.34 | 1.60 | 1.48 | 1.88 | 2.00 | 2.05 | 0.97 | |
| Bipolaris | 1.83 | 1.14 | 1.48 | 2.40 | 0.91 | 1.31 | 2.05 | 2.80 | |
| Purpureocillium | 4.45 | 2.00 | 0.63 | 0.97 | 1.77 | 1.88 | 1.48 | 0.74 | |
| Pyricularia | 2.05 | 1.94 | 1.54 | 1.48 | 1.60 | 1.65 | 2.34 | 1.14 | |
| Alternaria | 1.03 | 0.80 | 4.45 | 1.88 | 1.14 | 2.62 | 0.57 | 5.88 | |
| Sites | ||||||||
|---|---|---|---|---|---|---|---|---|
| G0 | G8 | L0 | L8 | N0 | N8 | S0 | S8 | |
| Bacteria | 2.12 | 2.09 | 2.05 | 1.99 b | 1.77 | 1.78 | 1.97 | 2.14 c |
| Fungi | 2.72 | 2.81 c | 2.68 | 2.60 c | 2.62 | 2.57 a | 2.73 | 2.33 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bogati, K.A.; Golińska, P.; Sewerniak, P.; Burkowska-But, A.; Walczak, M. Deciphering the Impact of Induced Drought in Agriculture Soils: Changes in Microbial Community Structure, Enzymatic and Metabolic Diversity. Agronomy 2023, 13, 1417. https://doi.org/10.3390/agronomy13051417
Bogati KA, Golińska P, Sewerniak P, Burkowska-But A, Walczak M. Deciphering the Impact of Induced Drought in Agriculture Soils: Changes in Microbial Community Structure, Enzymatic and Metabolic Diversity. Agronomy. 2023; 13(5):1417. https://doi.org/10.3390/agronomy13051417
Chicago/Turabian StyleBogati, Kalisa Amarsingh, Patrycja Golińska, Piotr Sewerniak, Aleksandra Burkowska-But, and Maciej Walczak. 2023. "Deciphering the Impact of Induced Drought in Agriculture Soils: Changes in Microbial Community Structure, Enzymatic and Metabolic Diversity" Agronomy 13, no. 5: 1417. https://doi.org/10.3390/agronomy13051417
APA StyleBogati, K. A., Golińska, P., Sewerniak, P., Burkowska-But, A., & Walczak, M. (2023). Deciphering the Impact of Induced Drought in Agriculture Soils: Changes in Microbial Community Structure, Enzymatic and Metabolic Diversity. Agronomy, 13(5), 1417. https://doi.org/10.3390/agronomy13051417