

Optimizing Planting Density in Alpine Mountain Strawberry Cultivation in Martell Valley, Italy


Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Management and Experimental Design
2.2. Evaluated Parameters
2.2.1. Morphological and Gas Exchange Parameters
2.2.2. Yield Parameters
2.2.3. Fruit Quality
2.2.4. Economic Analysis
2.3. Statistical Analysis
3. Results
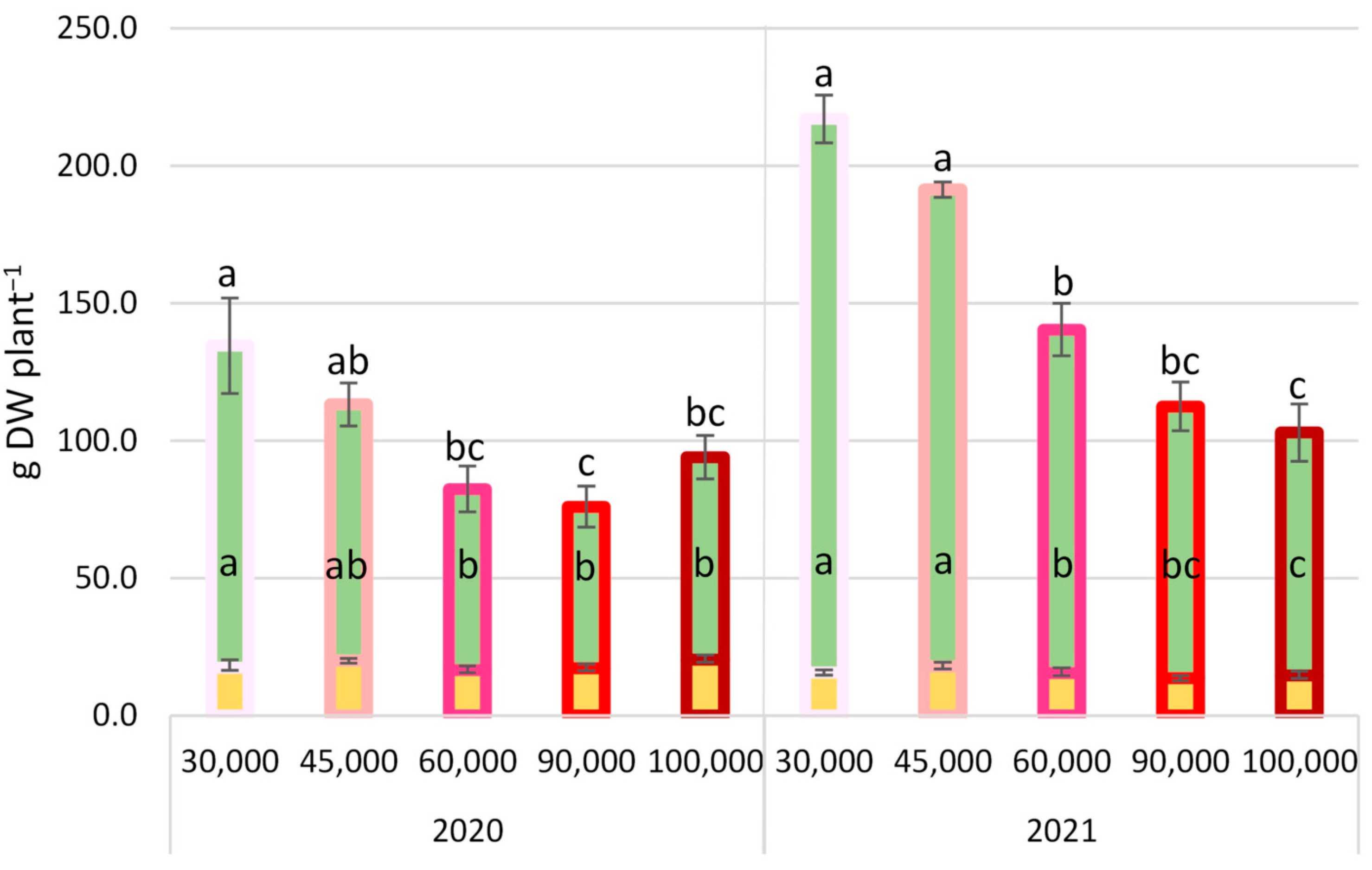
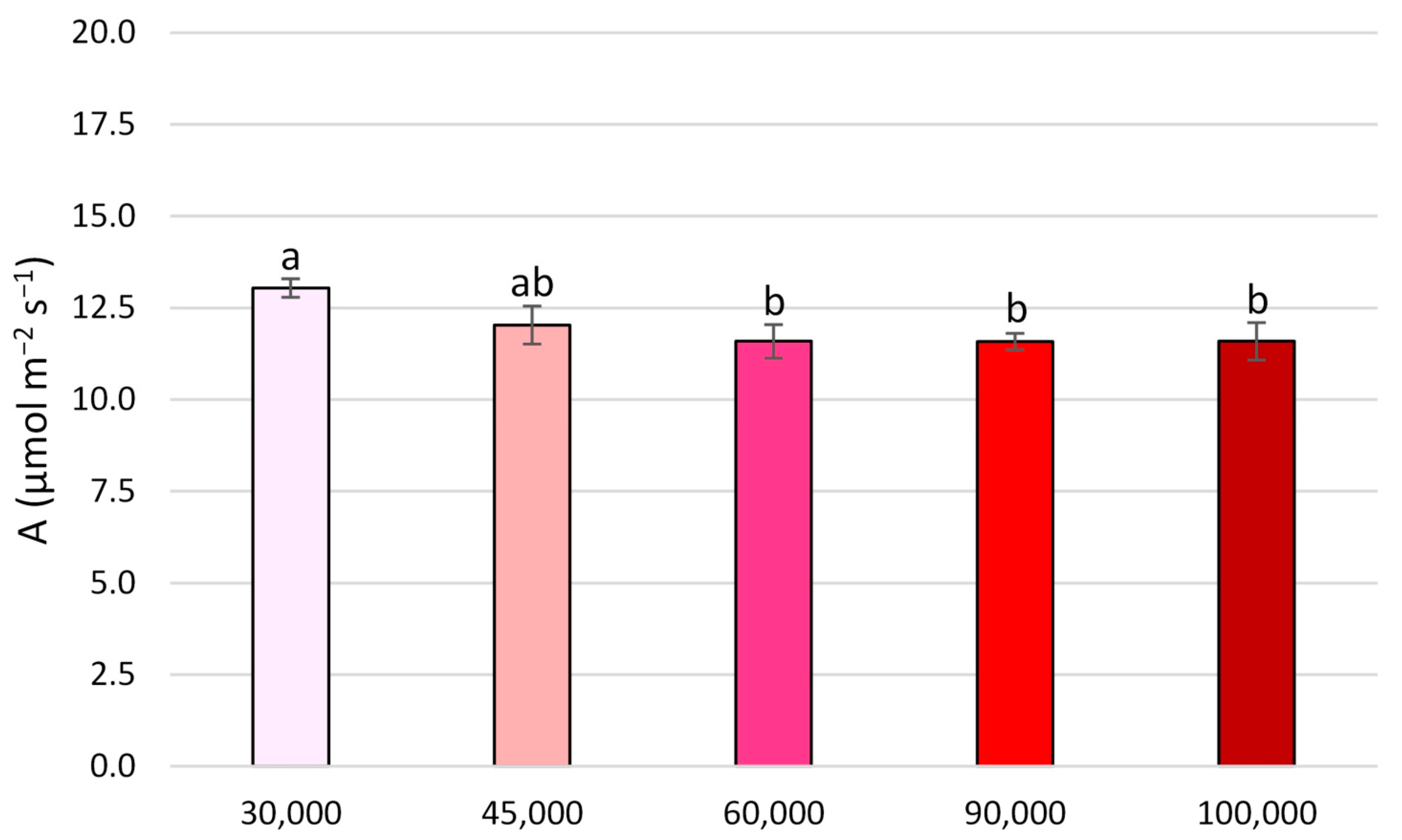
3.1. Morphological and Gas Exchange Parameters
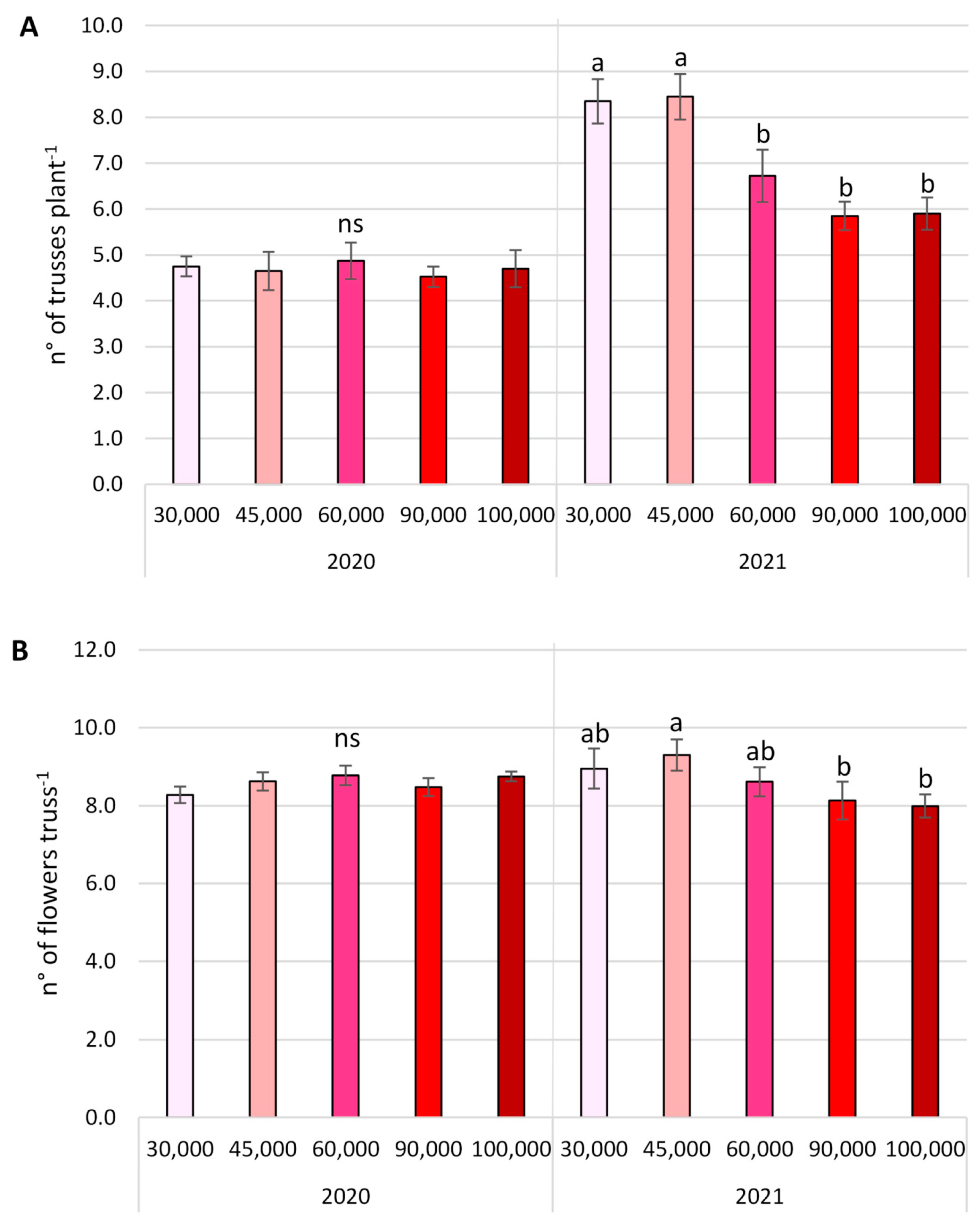
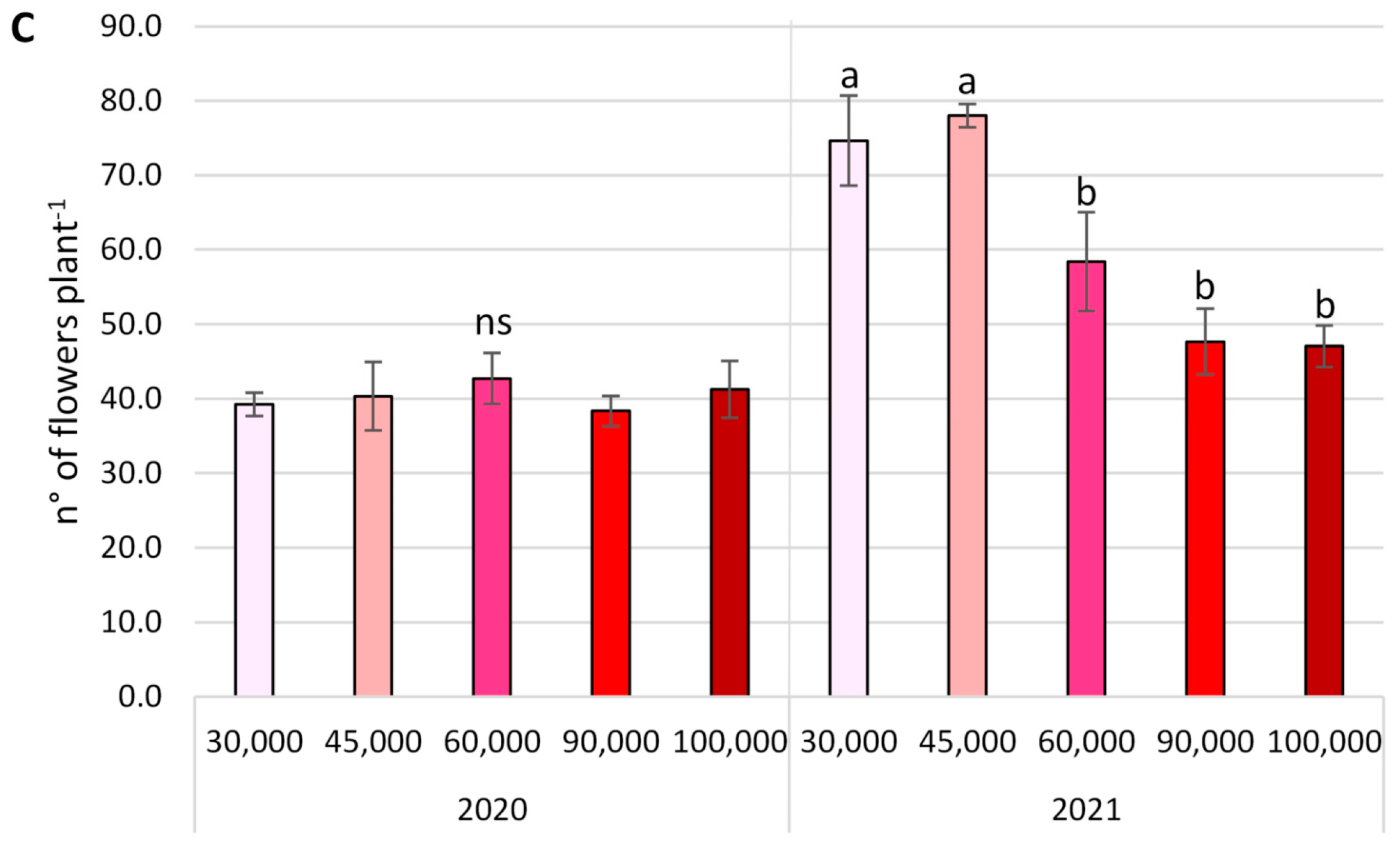
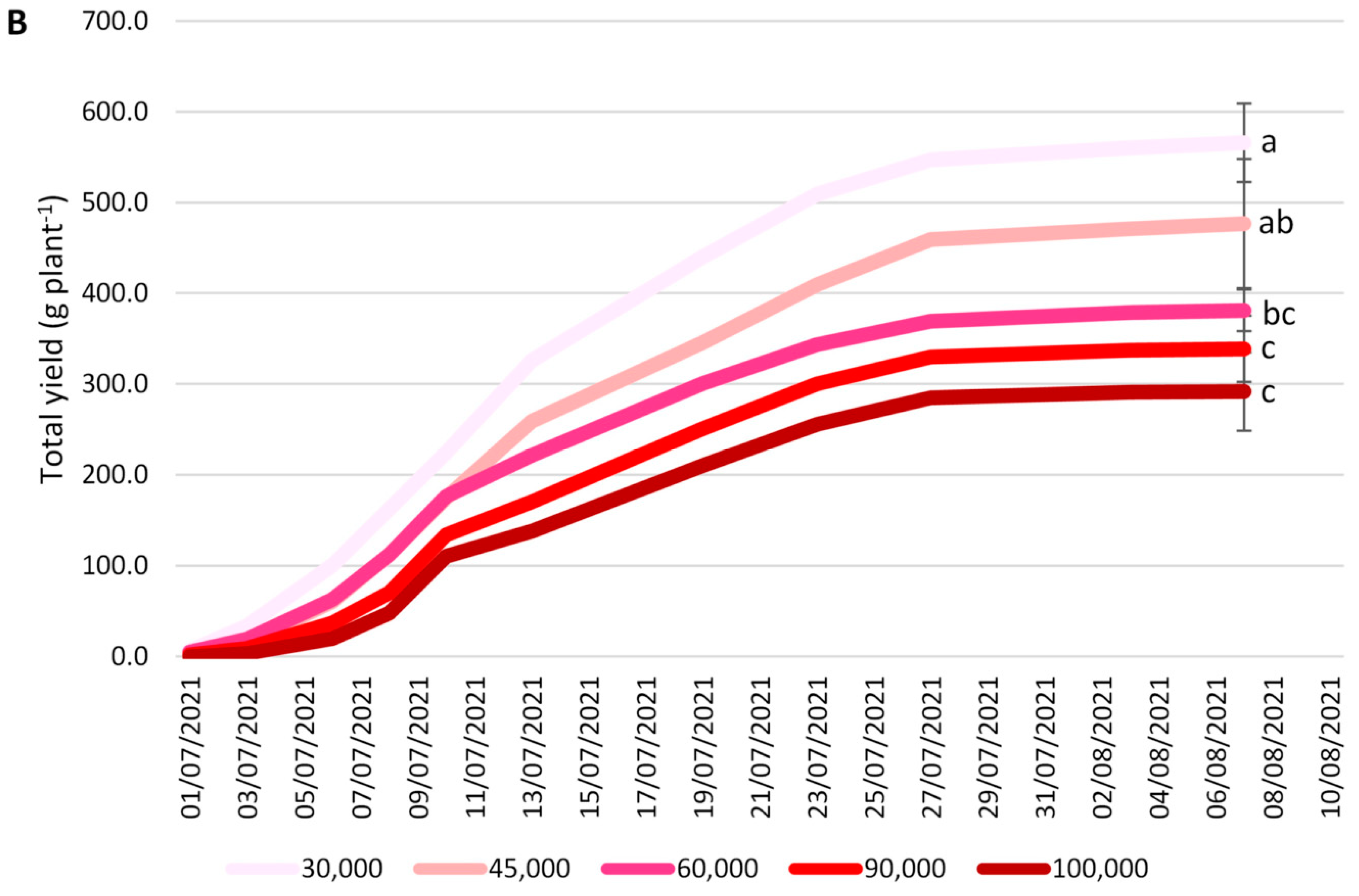
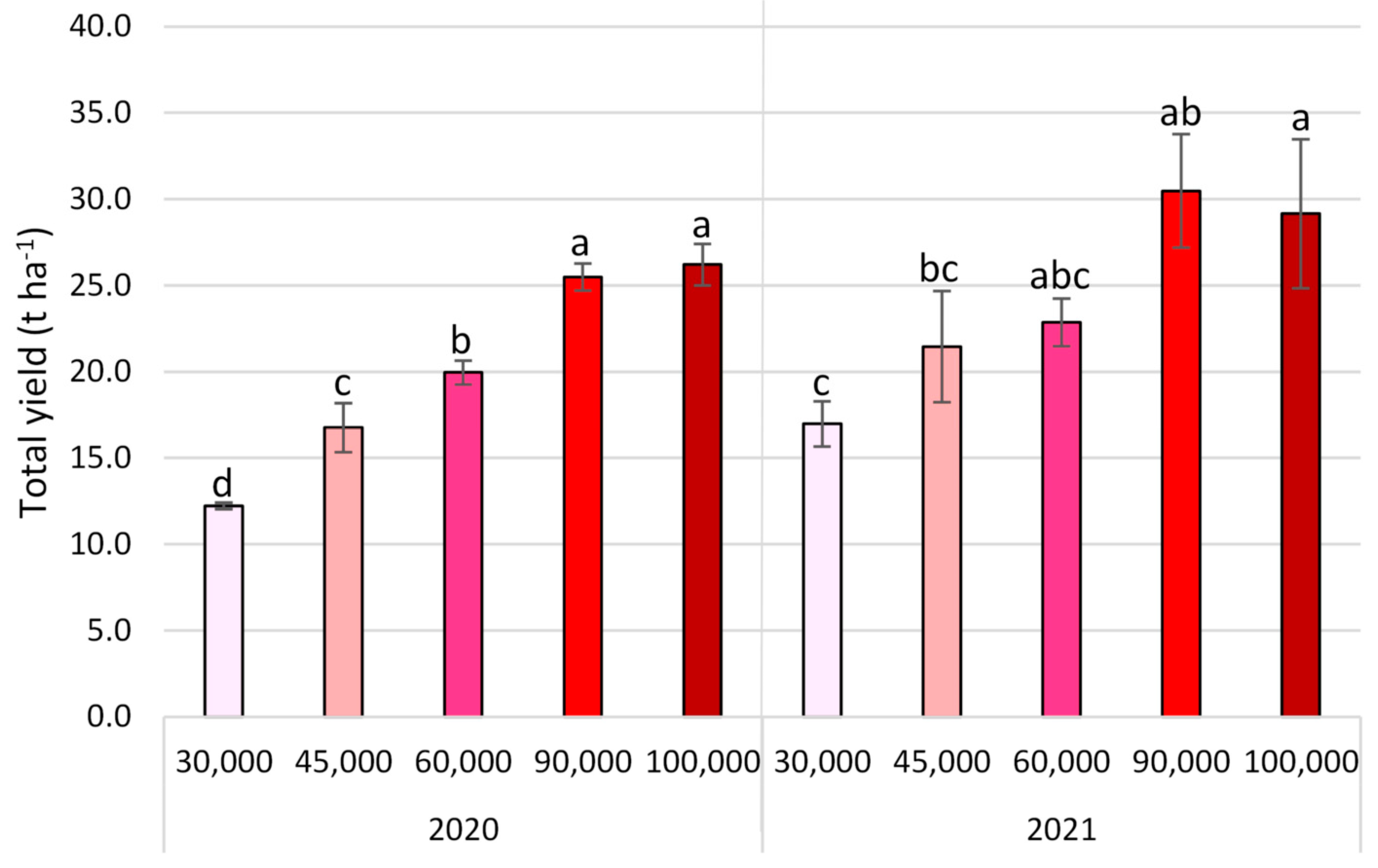
3.2. Yield Parameters
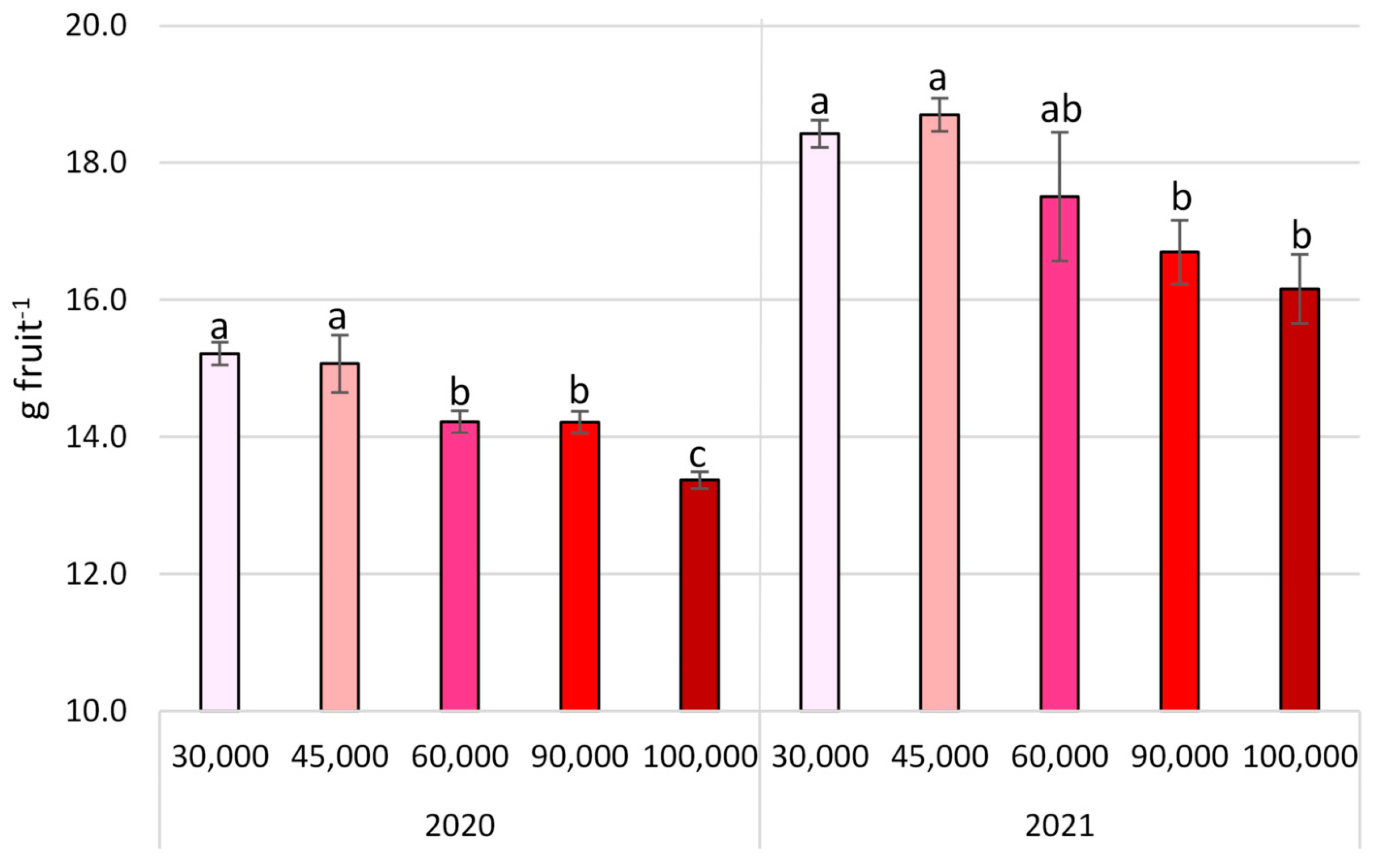
3.3. Fruit Quality
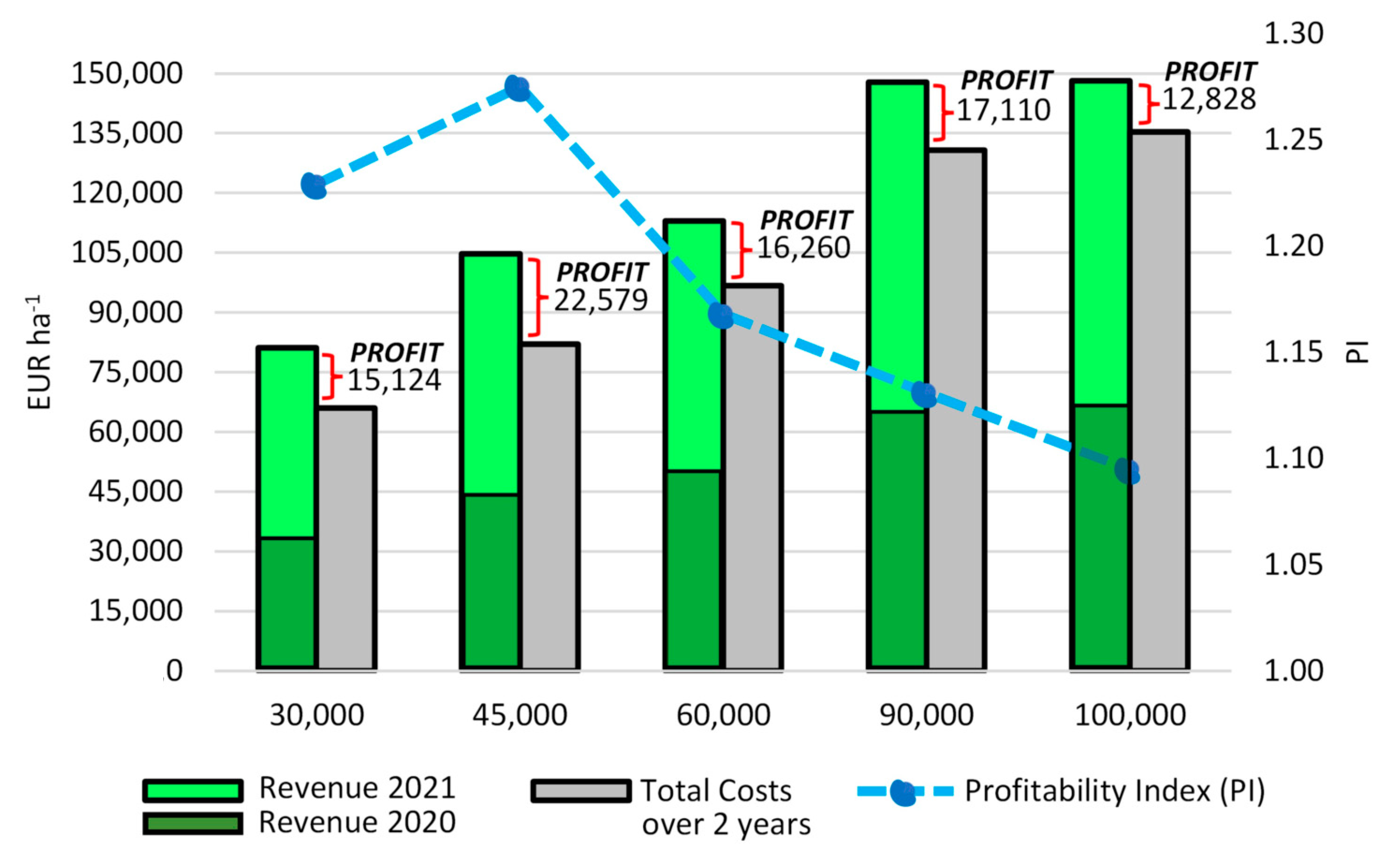
3.4. Economic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liston, A.; Cronn, R.; Ashman, T. Fragaria: A Genus with Deep Historical Roots and Ripe for Evolutionary and Ecological Insights. Am. J. Bot. 2014, 101, 1686–1699. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Su, D.; Wei, S.; Chen, S.; Luo, Z.; Shen, X.; Zhang, Z.; Jamil, A.; Tong, J.; Cui, X. Current and Future Potential Distribution of Wild Strawberry Species in the Biodiversity Hotspot of Yunnan Province, China. Agronomy 2020, 10, 959. [Google Scholar] [CrossRef]
- Darrow, G.M. The Strawberry: History, Breeding, and Physiology; Holt, Rinehart and Winston: New York, NY, USA, 1966. [Google Scholar]
- FAOSTAT. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 20 October 2022).
- ISTAT. Available online: https://www.istat.it (accessed on 20 October 2022).
- ASTAT. Available online: https://astat.provincia.bz.it/it/default.asp (accessed on 20 October 2022).
- Cooperative for Regional Development Martello 3B. Available online: https://www.martell.it/it/la-val-martello/ (accessed on 26 October 2022).
- Bargioni, G. The Italian Strawberry Industry. In Proceedings of the Acta Horticulturae, International Society for Horticultural Science (ISHS), Leuven, Belgium, 1 December 1989; pp. 659–664. [Google Scholar]
- Mezzetti, B.; Giampieri, F.; Zhang, Y.; Zhong, C. Status of Strawberry Breeding Programs and Cultivation Systems in Europe and the Rest of the World. J. Berry Res. 2018, 8, 205–221. [Google Scholar] [CrossRef]
- Haque, M.A.; Sakimin, S.Z. Planting Arrangement and Effects of Planting Density on Tropical Fruit Crops—A Review. Horticulturae 2022, 8, 485. [Google Scholar] [CrossRef]
- Djaman, K.; Allen, S.; Djaman, D.S.; Koudahe, K.; Irmak, S.; Puppala, N.; Darapuneni, M.K.; Angadi, S.V. Planting Date and Plant Density Effects on Maize Growth, Yield and Water Use Efficiency. Environ. Chall. 2022, 6, 100417. [Google Scholar] [CrossRef]
- Hou, W.; Khan, M.R.; Zhang, J.; Lu, J.; Ren, T.; Cong, R.; Li, X. Nitrogen Rate and Plant Density Interaction Enhances Radiation Interception, Yield and Nitrogen Use Efficiency of Mechanically Transplanted Rice. Agric. Ecosyst. Environ. 2019, 269, 183–192. [Google Scholar] [CrossRef]
- Rodrigo, V.H.L.; Stirling, C.M.; Teklehaimanot, Z.; Nugawela, A. The Effect of Planting Density on Growth and Development of Component Crops in Rubber/Banana Intercropping Systems. Field Crops Res. 1997, 52, 95–108. [Google Scholar] [CrossRef]
- Wright, C.J.; Sandrang, A.K. Density Effects on Vegetative and Reproductive Development in Strawberry Cv. Hapil. J. Hortic. Sci. 1993, 68, 231–236. [Google Scholar] [CrossRef]
- Wu, L.; Deng, Z.; Cao, L.; Meng, L. Effect of Plant Density on Yield and Quality of Perilla Sprouts. Sci. Rep. 2020, 10, 9937. [Google Scholar] [CrossRef] [PubMed]
- Martins de Lima, J.; Welter, P.D.; Soares dos Santos, M.F.; Kavcic, W.; Miranda Costa, B.; Fagherazzi, A.F.; Regianini Nerbass, F.; Anneliese Kretzschmar, A.; Rufato, L.; Baruzzi, G. Planting Density Interferes with Strawberry Production Efficiency in Southern Brazil. Agronomy 2021, 11, 408. [Google Scholar] [CrossRef]
- Postma, J.A.; Hecht, V.L.; Hikosaka, K.; Nord, E.A.; Pons, T.L.; Poorter, H. Dividing the Pie: A Quantitative Review on Plant Density Responses. Plant Cell Environ. 2021, 44, 1072–1094. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Lu, X.; Gu, S.; Guo, X. Improving Nutrient and Water Use Efficiencies Using Water-Drip Irrigation and Fertilization Technology in Northeast China. Agric. Water Manag. 2020, 241, 106352. [Google Scholar] [CrossRef]
- Li, T.; Liu, L.-N.; Jiang, C.-D.; Liu, Y.-J.; Shi, L. Effects of Mutual Shading on the Regulation of Photosynthesis in Field-Grown Sorghum. J. Photochem. Photobiol. B 2014, 137, 31–38. [Google Scholar] [CrossRef]
- Pierson, E.A.; Mack, R.N.; Black, R.A. The Effect of Shading on Photosynthesis, Growth, and Regrowth Following Defoliation for Bromus Tectorum. Oecologia 1990, 84, 534–543. [Google Scholar] [CrossRef]
- Samad, S.; Rivero, R.; Kalyandurg, P.B.; Vetukuri, R.R.; Heide, O.M.; Sønsteby, A.; Khalil, S. Characterization of Environmental Effects on Flowering and Plant Architecture in an Everbearing Strawberry F1-Hybrid by Meristem Dissection and Gene Expression Analysis. Horticulturae 2022, 8, 626. [Google Scholar] [CrossRef]
- Sidhu, V.; Bernier-English, V.; Lamontagne-Drolet, M.; Gravel, V. Effect of Light Quality and Extended Photoperiod on Flower Bud Induction during Transplant Production of Day-Neutral Strawberry Cultivars. Can. J. Plant Sci. 2022, 102, 356–367. [Google Scholar] [CrossRef]
- Sønsteby, A.; Heide, O.M. Dynamics of Dormancy Regulation in ‘Sonata’ Strawberry and Its Relation to Flowering and Runnering. CABI Agric. Biosci. 2021, 2, 4. [Google Scholar] [CrossRef]
- Sønsteby, A.; Hytonen, T. Manipulating Flower Induction through Temperature and Photoperiod Fluctuations. Int. J. Fruit Sci. 2005, 5, 17–27. [Google Scholar] [CrossRef]
- Wang, R.; Eguchi, M.; Gui, Y.; Iwasaki, Y. Evaluating the Effect of Light Intensity on Flower Development Uniformity in Strawberry (Fragaria × ananassa) under Early Induction Conditions in Forcing Culture. HortScience 2020, 55, 670–675. [Google Scholar] [CrossRef]
- Dhaliwal, D.S.; Williams, M.M. Understanding Variability in Optimum Plant Density and Recommendation Domains for Crowding Stress Tolerant Processing Sweet Corn. PLoS ONE 2020, 15, e0228809. [Google Scholar] [CrossRef]
- Wamser, A.F.; Valmorbida, J.; Suzuki, A.; Hahn, L.; Mueller, S.; Becker, W.F.; Feltrim, A.L.; Ender, M.M. Planting Density and Arrangement for the Mechanized Spraying of Vertically Staked Tomatoes. Hortic. Bras. 2017, 35, 519–526. [Google Scholar] [CrossRef]
- Carvalho, F.J.; Carneiro, L.B.; Benett, C.G.S.; Benett, K.S.S.; Martins, A.S.; Silva, A.T.D.; Seleguini, A. Plant Density and Growth Regulator Applications in a Tomato Crop for Industrial Processing. Rev. Colomb. Cienc. Hortíc. 2019, 13, 397–405. [Google Scholar] [CrossRef]
- De Camacaro, M.E.P.; Camacaro, G.J.; Hadley, P.; Dennett, M.D.; Battey, N.H.; Carew, J.G. Effect of Plant Density and Initial Crown Size on Growth, Development and Yield in Strawberry Cultivars Elsanta and Bolero. J. Hortic. Sci. Biotechnol. 2004, 79, 739–746. [Google Scholar] [CrossRef]
- Heuvelink, E. Effect of Plant Density on Biomass Allocation to the Fruits in Tomato (Lycopersicon esculentum Mill.). Sci. Hortic. 1995, 64, 193–201. [Google Scholar] [CrossRef]
- Khasmakhi-Sabet, A.; Sedaghathoor, S.; Mohammady, J.; Olfati, J.A. Effect of Plant Density on Bell Pepper Yield and Quality. Int. J. Veg. Sci. 2009, 15, 264–271. [Google Scholar] [CrossRef]
- Nerson, H. Relationship between Plant Density and Fruit and Seed Production in Muskmelon. J. Am. Soc. Hortic. Sci. 2002, 127, 855–859. [Google Scholar] [CrossRef]
- Ramos-López, B.I.; Ortiz-Hernández, Y.D.; Morales, I.; Aquino-Bolaños, T. Plant Density on Yield of Husk Tomato (Physalis ixocarpa Brot.) in Field and Greenhouse. Ciênc. Rural 2021, 51, e20200992. [Google Scholar] [CrossRef]
- Zenobi, S.; Fiorentini, M.; Aquilanti, L.; Foligni, R.; Mannozzi, C.; Mozzon, M.; Zitti, S.; Casavecchia, S.; Al Mohandes Dridi, B.; Orsini, R. Effect of Planting Density in Two Thistle Species Used for Vegetable Rennet Production in Marginal Mediterranean Areas. Agronomy 2021, 11, 135. [Google Scholar] [CrossRef]
- Gale, J. Plants and Altitude—Revisited. Ann. Bot. 2004, 94, 199. [Google Scholar] [CrossRef]
- Guerrero-Chavez, G.; Scampicchio, M.; Andreotti, C. Influence of the Site Altitude on Strawberry Phenolic Composition and Quality. Sci. Hortic. 2015, 192, 21–28. [Google Scholar] [CrossRef]
- Kiełtyk, P. Patterns of Floral Allocation along an Elevation Gradient: Variation in Senecio Subalpinus Growing in the Tatra Mountains. Alp. Bot. 2021, 131, 117–124. [Google Scholar] [CrossRef]
- Wang, H.; Prentice, I.C.; Davis, T.W.; Keenan, T.F.; Wright, I.J.; Peng, C. Photosynthetic Responses to Altitude: An Explanation Based on Optimality Principles. N. Phytol. 2017, 213, 976–982. [Google Scholar] [CrossRef] [PubMed]
- Tessmer, M.A.; Appezzato-da-Glória, B.; Antoniolli, L.R. Influence of Growing Sites and Physicochemical Features on the Incidence of Lenticel Breakdown in ‘Gala’ and ‘Galaxy’ Apples. Sci. Hortic. 2016, 205, 119–126. [Google Scholar] [CrossRef]
- Heitholt, J.J.; Sassenrath-Cole, G.F. Inter-Plant Competition: Growth Responses to Plant Density and Row Spacing. In Physiology of Cotton; Stewart, J.D., Oosterhuis, D.M., Heitholt, J.J., Mauney, J.R., Eds.; Springer: Dordrecht, The Netherland, 2010; pp. 179–186. ISBN 978-90-481-3194-5. [Google Scholar]
- Yang, X.; Zhang, W.; He, Q. Effects of Intraspecific Competition on Growth, Architecture and Biomass Allocation of Quercus Liaotungensis. J. Plant Interact. 2019, 14, 284–294. [Google Scholar] [CrossRef]
- Craine, J.M.; Dybzinski, R. Mechanisms of Plant Competition for Nutrients, Water and Light. Funct. Ecol. 2013, 27, 833–840. [Google Scholar] [CrossRef]
- Belcher, J.W.; Keddy, P.A.; Twolan-Strutt, L. Root and Shoot Competition Intensity along a Soil Depth Gradient. J. Ecol. 1995, 83, 673–682. [Google Scholar] [CrossRef]
- Schenk, H.J. Root Competition: Beyond Resource Depletion. J. Ecol. 2006, 94, 725–739. [Google Scholar] [CrossRef]
- Wang, L.-W.; Showalter, A.M.; Ungar, I.A. Effects of Intraspecific Competition on Growth and Photosynthesis of Atriplex Prostrata. Aquat. Bot. 2005, 83, 187–192. [Google Scholar] [CrossRef]
- Wilson, S.D.; Tilman, D. Plant Competition and Resource Availability in Response to Disturbance and Fertilization. Ecology 1993, 74, 599–611. [Google Scholar] [CrossRef]
- Yang, J.; Jeong, B.R. Side Lighting Enhances Morphophysiology by Inducing More Branching and Flowering in Chrysanthemum Grown in Controlled Environment. Int. J. Mol. Sci. 2021, 22, 12019. [Google Scholar] [CrossRef]
- Hytönen, T.; Palonen, P.; Mouhu, K.; Junttila, O. Crown Branching and Cropping Potential in Strawberry (Fragaria ananassa Duch.) Can Be Enhanced by Daylength Treatments. J. Hortic. Sci. Biotechnol. 2004, 79, 466–471. [Google Scholar] [CrossRef]
- Koskela, E.A.; Hytönen, T. Control of Flowering in Strawberries. In The Genomes of Rosaceous Berries and Their Wild Relatives; Hytönen, T., Graham, J., Harrison, R., Eds.; Compendium of Plant Genomes; Springer: Cham, Switzerland, 2018; pp. 35–48. ISBN 978-3-319-76019-3. [Google Scholar]
- Tuller, J.; Marquis, R.J.; Andrade, S.M.M.; Monteiro, A.B.; Faria, L.D.B. Trade-Offs between Growth, Reproduction and Defense in Response to Resource Availability Manipulations. PLoS ONE 2018, 13, e0201873. [Google Scholar] [CrossRef] [PubMed]
- Soppelsa, S.; Kelderer, M.; Casera, C.; Bassi, M.; Robatscher, P.; Matteazzi, A.; Andreotti, C. Foliar Applications of Biostimulants Promote Growth, Yield and Fruit Quality of Strawberry Plants Grown under Nutrient Limitation. Agronomy 2019, 9, 483. [Google Scholar] [CrossRef]
- Commission of the European Communities. Annex to the Report from the Commission to the Council and the European Parliament on the Situation of the Sector of Soft Fruits and Cherries Intended for Processing; European Commission: Brussels, Belgium, 2006. [Google Scholar]
- Naryal, A.; Dolkar, D.; Bhardwaj, A.K.; Kant, A.; Chaurasia, O.P.; Stobdan, T. Effect of Altitude on the Phenology and Fruit Quality Attributes of Apricot (Prunus armeniaca L.) Fruits. Def. Life Sci. J. 2020, 5, 18–24. [Google Scholar] [CrossRef]
- Matsumoto, S.; Tsuboi, T.; Asea, G.; Miyamoto, K.; Maruyama, A.; Takagaki, M.; Kikuchi, M. Effects of Plant Density on the Performance of Selected African Upland Rice Varieties. Afr. J. Agric. Res. 2017, 12, 2262–2272. [Google Scholar] [CrossRef]
- Castellanos, J.Z.; Vargas-Tapia, P.; Ojodeagua, J.L.; Hoyos, G.; Alcantar-Gonzalez, G.; Mendez, F.S.; Alvarez-Sanchez, E.; Gardea, A.A. Garlic Productivity and Profitability as Affected by Seed Clove Size, Planting Density and Planting Method. HortScience 2004, 39, 1272–1277. [Google Scholar] [CrossRef]
- Jettner, R.; Loss, S.P.; Martin, L.D.; Siddique, K.H.M. Responses of Faba Bean (Vicia faba L.) to Sowing Rate in South-Western Australia. I. Seed Yield and Economic Optimum Plant Density. Aust. J. Agric. Res. 1998, 49, 989. [Google Scholar] [CrossRef]
- Di Stefano, V.; Scandurra, S.; Pagliaro, A.; Di Martino, V.; Melilli, M.G. Effect of Sunlight Exposure on Anthocyanin and Non-Anthocyanin Phenolic Levels in Pomegranate Juices by High Resolution Mass Spectrometry Approach. Foods 2020, 9, 1161. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, Y.; Koyama, N.; Tamura, H. Color and Anthocyanin Composition of Strawberry Fruit. Changes during Fruit Development and Differences among Cultivars, with Special Reference to the Occurrence of Pelargonidin 3-Malonylglucoside. J. Jpn. Soc. Hortic. 2002, 71, 355–361. [Google Scholar] [CrossRef]
- Bhat, R.; Geppert, J.; Funken, E.; Stamminger, R. Consumers Perceptions and Preference for Strawberries—A Case Study from Germany. Int. J. Fruit Sci. 2015, 15, 405–424. [Google Scholar] [CrossRef]
- Lee, M.A.; Burger, G.; Green, E.R.; Kooij, P.W. Relationships between Resource Availability and Elevation Vary between Metrics Creating Gradients of Nutritional Complexity. Oecologia 2021, 195, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Maroto, J.V.; López-Galarza, S.; San Bautista, A.; Pascual, B. Cold Stored and Fresh Multicrown Strawberry Plants for Autumn-Winter Production in Eastern Spain. In Proceedings of the Acta Horticulturae, International Society for Horticultural Science (ISHS), Leuven, Belgium, 1 September 1997; pp. 545–548. [Google Scholar]
- Pirlak, L.; Güleryüz, M.; Bolat, I. The Altitude Affects the Runner Plant Production and Quality in Strawberry Cultivars. In Proceedings of the Acta Horticulturae, International Society for Horticultural Science (ISHS), Leuven, Belgium, 31 January 2002; pp. 305–308. [Google Scholar]
- González, J.A.; Mercado, M.I.; Martinez-Calsina, L.; Erazzú, L.E.; Buedo, S.E.; González, D.A.; Ponessa, G.I. Plant Density Effects on Quinoa Yield, Leaf Anatomy, Ultrastructure and Gas Exchange. J. Agric. Sci. 2022, 160, 349–359. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
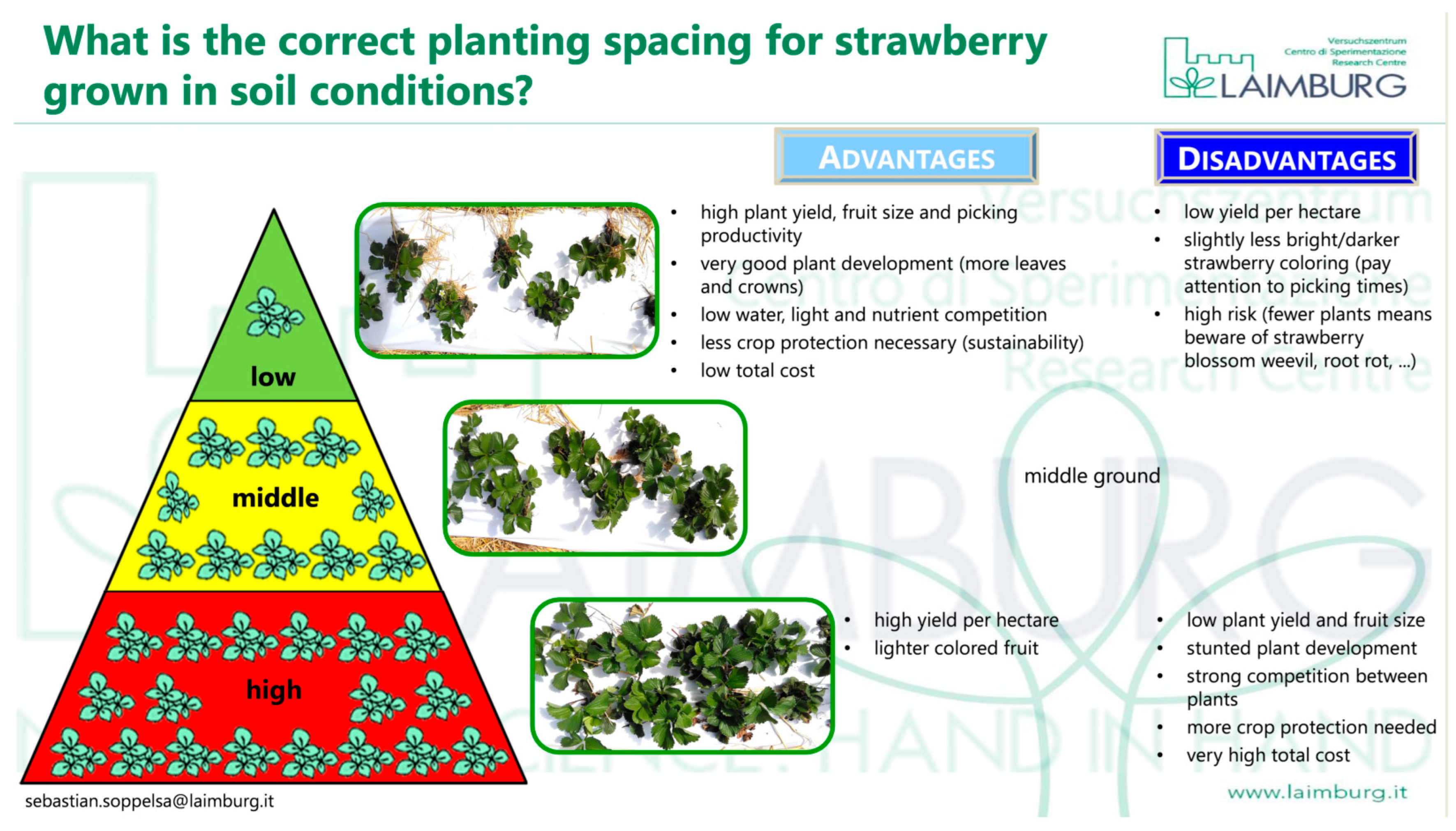{kind=link}
| Air Temperature (°C) | Relative Humidity (%) | Rainfall (mm) | |||
|---|---|---|---|---|---|
| Minimum Temperature | Maximum Temperature | Mean Temperature | |||
| 2020 | |||||
| May | 0.1 | 22.2 | 10.9 | 67.9 | 56.8 |
| June | 3.4 | 27.6 | 13.8 | 76.0 | 97.6 |
| July | 5.8 | 31.0 | 16.8 | 74.6 | 70.5 |
| August | 6.0 | 31.7 | 16.3 | 80.1 | 120.4 |
| 2021 | |||||
| May | 0.0 | 21.4 | 9.3 | 64.9 | 130.8 |
| June | 6.6 | 30.0 | 17.5 | 64.0 | 39.8 |
| July | 7.6 | 27.8 | 17.0 | 71.6 | 135.0 |
| August | 6.2 | 29.2 | 15.9 | 71.8 | 137.4 |
| First-Class Yield (g Plant−1) | Small Fruits (g Plant−1) | Misshapen Fruits (g Plant−1) | Rotten Fruits (g Plant−1) | |||||
|---|---|---|---|---|---|---|---|---|
| Plant density level (D) | ||||||||
| 30,000 | 394.60 ± 61.18 1 | A | 39.93 ± 4.64 | A | 50.18 ± 0.02 | A | 1.94 ± 0.56 | A |
| 45,000 | 336.95 ± 45.23 | AB | 39.61 ± 7.55 | A | 45.36 ± 0.73 | A | 2.71 ± 0.14 | A |
| 60,000 | 266.74 ± 23.06 | BC | 42.20 ± 2.33 | A | 45.99 ± 2.97 | A | 1.89 ± 0.64 | A |
| 90,000 | 232.65 ± 21.67 | C | 42.83 ± 1.31 | A | 34.14 ± 0.17 | B | 1.26 ± 0.55 | A |
| 100,000 | 211.74 ± 17.33 | C | 32.03 ± 1.18 | A | 31.09 ± 5.23 | B | 1.96 ± 0.50 | A |
| Significance | *** | ns | *** | ns | ||||
| Year (Y) | ||||||||
| 2020 | 240.89 ± 24.98 | 44.13 ± 3.11 | 43.92 ± 3.58 | 2.63 ± 0.17 | ||||
| 2021 | 336.18 ± 51.15 | 34.51 ± 2.63 | 38.79 ± 5.14 | 1.28 ± 0.38 | ||||
| Significance | *** | ** | ns | ** | ||||
| D × Y | ns | ns | ns | ns | ||||
| FF (Durofel Index) | TSS (°Brix) | TA (g Acid Citric L−1) | CI | |||||
|---|---|---|---|---|---|---|---|---|
| Plant density level (D) | ||||||||
| 30,000 | 36.66 ± 4.87 1 | A | 7.39 ± 0.09 | A | 6.67 ± 0.03 | A | 39.64 ± 1.78 | A |
| 45,000 | 36.19 ± 2.48 | A | 7.23 ± 0.12 | A | 6.50 ± 0.15 | A | 38.35 ± 1.10 | AB |
| 60,000 | 35.73 ± 2.63 | A | 7.04 ± 0.04 | A | 6.36 ± 0.34 | A | 36.12 ± 2.20 | ABC |
| 90,000 | 36.23 ± 2.81 | A | 7.09 ± 0.01 | A | 6.64 ± 0.04 | A | 35.86 ± 1.33 | BC |
| 100,000 | 35.87 ± 3.47 | A | 7.15 ± 0.09 | A | 6.85 ± 0.07 | A | 33.85 ± 1.82 | C |
| Significance | ns | ns | ns | *** | ||||
| Year (Y) | ||||||||
| 2020 | 40.74 ± 0.83 | 7.15 ± 0.05 | 6.77 ± 0.06 | 34.44 ± 1.25 | ||||
| 2021 | 31.54 ± 0.59 | 7.21 ± 0.12 | 6.44 ± 0.18 | 39.09 ± 1.09 | ||||
| Significance | *** | ns | ns | *** | ||||
| D × Y | ns | ns | ns | ns | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soppelsa, S.; Gasser, M.; Zago, M. Optimizing Planting Density in Alpine Mountain Strawberry Cultivation in Martell Valley, Italy. Agronomy 2023, 13, 1422. https://doi.org/10.3390/agronomy13051422
Soppelsa S, Gasser M, Zago M. Optimizing Planting Density in Alpine Mountain Strawberry Cultivation in Martell Valley, Italy. Agronomy. 2023; 13(5):1422. https://doi.org/10.3390/agronomy13051422
Chicago/Turabian StyleSoppelsa, Sebastian, Michael Gasser, and Massimo Zago. 2023. "Optimizing Planting Density in Alpine Mountain Strawberry Cultivation in Martell Valley, Italy" Agronomy 13, no. 5: 1422. https://doi.org/10.3390/agronomy13051422
APA StyleSoppelsa, S., Gasser, M., & Zago, M. (2023). Optimizing Planting Density in Alpine Mountain Strawberry Cultivation in Martell Valley, Italy. Agronomy, 13(5), 1422. https://doi.org/10.3390/agronomy13051422