
Resistance to Anthracnose Rot Disease in Capsicum


Abstract
:1. Introduction
2. Bioassays for Evaluating Anthracnose Resistance

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathogen a | Bioassay b | Inoculation Method c | Disease Evaluation Method d | Phenotypic Responses e | Reference |
|---|---|---|---|---|---|
| C. scovillei | 1 | 1 | DSI | R (1–<3); IR (3–<6); S (6–9) | [52] |
| 2 | 1/2 | DSI | HR (0); R (0.1–1.9); MR (2.0–2.9); S (3.0–4.9); HS (5) | [30] | |
| 4 | 4 | Infection rate | R (infection rate < 10%) | [53] | |
| 3 | 3 | DI | HR (0–<10%); R (10–<20%); MR (20–<40%); S (40–<70%); HS (>70%) | [45] | |
| 3 | 3 | DSI, AUDPC, IP, LP | R (low value for AUDPC, and high values for IP and LP) | [16] | |
| 3 | 3 | DSI | HR (1); R (3); MR (4); MS (6); S (8); HS (10) | [41] | |
| C. brevisporum | 5 | 3 | DSI based on LS | R (<10%); MR (11–20%); S (21–40%); HS (>41%) | [36] |
| 6 | 1 | DSI based on LS (foliar) | 1 (SL); 2 (>1 mm); 3 (1%); 4 (5%); 5 (10%); 6 (25%) | [36] | |
| C. capsici | 7 | 2 | DSI based on LS | R (0–1.0); MR (1.1–2.0); MS (2.1–3.0); S (3.1–4.0); HS (4.1–5.0) | [40] |
| - | 4 | DSI based on DI | I (0%); R (1–5%); MR (6–25%); S (26–50%); HS (51–100%) | [51] | |
| 7 | 1 | DSI based on LS | R (<20%); MR (21–40%); MS (41–60%); S (>60%) | [46] | |
| 6 | 1/2 | DSI based on LS | SL (0); R (0.1–10%); MR (10.1–25%); MS (25.1–50%); S (50.1–75%); HS (75.1–100%) | [31] | |
| C. coccodes | 6 | 1/5 | DSI and AUDPC | Index (0–5) | [54] |
| C. siamense | 3 | 3 | DSI | HR (1); R (3); MR (4); MS (6); S (8); HS (10) | [55] |
| C. siamense | 8 | 6/3 | DSI and AUDPC | HR (1); R (3); MR (4); MS (6); S (8); HS (10) | [48] |
| C. capsid, C. siamense | 7 | 2 | DSI based on DI | I (0); R (1); MR (2); S (3); HS (4) | [39] |
| C. scovillei | 2 | 3 | Lesion diameter and infection frequency | [26,47] | |
| C. capsici, C. siamense | 3 | 2 | [25] | ||
| C. siamense | 2 | 3 | LI | [56] | |
| C. scovillei, C. capsici, C. siamense, C. truncatum | 8 | 1/3 | DSI based on LS | HR (0); R (1); MR (3); MS (5); S (7); HS (9) | [28,44,49,50,57] |
| Colletotrichum spp. | 3 | 1 | DI, lesion diameter, LGR, AUDPC, IP | R (low values for AUDPC, DI, lesion diameter, and LGR; high values for IP) | [58] |
3. Sources of Anthracnose Resistance in Capsicum Germplasm
4. Breeding for Anthracnose Resistance in Capsicum
| Genetic Source | Resistance Level a | Population | Colletotrichum spp. b | Inoculation Method | Genetic Mechanism c | Reference | |||
|---|---|---|---|---|---|---|---|---|---|
| Capsicum spp. | Accession | Type | Susceptible Parent | Mature Green | Ripe | ||||
| C. annuum | Chungryong | MR | F2, BC1 | C. annuum PI244670 | C. dematium, C. siamense | Detached pinpricking | Partial dominant * | Partial dominant * | [59,68] |
| Perennial | MR | F2, BC1, BC2 | C. annuum ‘Kolascai E-14’ | C. truncatum | - | Polygenic | na | [60,68] | |
| 83–168 | MR | F2, BC1 | C. annuum ‘KKU-Cluster’ | C. capsici | Detached dropping | Single dominant | na | [73] | |
| Punjab Lal | R | F2 | C. annuum ‘PT 12-3’ | C. scovillei, C. capsici | Detached pinpricking | Polygenic | Polygenic | [77] | |
| GBUEL104 | HR | F2, BC1, BC2 | C. annuum GBUEL103 | C. scovillei | Detached microinfection | Two dominant QTLs * | Two dominant QTLs * | [41] | |
| C. chinense | PRI95030 | HR | F2 | C. annuum ‘Jatilaba’ | C. siamense, C. capsici | Detached pinpricking | na | Polygenic | [25] |
| PBC932 | R | F2, BC1, BC3 | C. annuum ‘9955-15’ | C. scovillei | Detached microinfection | Two dominant genes * | Polygenic recessive * | [72] | |
| PBC932 | HR | F2, BC1 | C. annuum ‘Yeoju’, ‘Bangchang’ | C. capsici | Single recessive * | Single recessive * | [22,26] | ||
| PBC932 | R | BC1 | C. annuum ‘77013’ | C. scovillei | Polygenic dominant * | Polygenic dominant * | [15] | ||
| C. baccatum | PI594137 | R | F2, BC1 | C. baccatum Golden-aji | C. scovillei | Detached microinfection | Single dominant | na | [47] |
| PBC80 | HR | F2, BC1 | C. baccatum CA1316 | C. scovillei | Single recessive * | Single dominant * | [28,34,43] | ||
| PBC81 | HR | F2, BC1 | C. annuum SP26, Matikas | C. scovillei, C. capsici | Polygenic * | Polygenic * | [29,78] | ||
| 881045 (Cbp) | R | F2 | C. baccatum Golden-aji | C. scovillei | na | Polygenic | [33] | ||
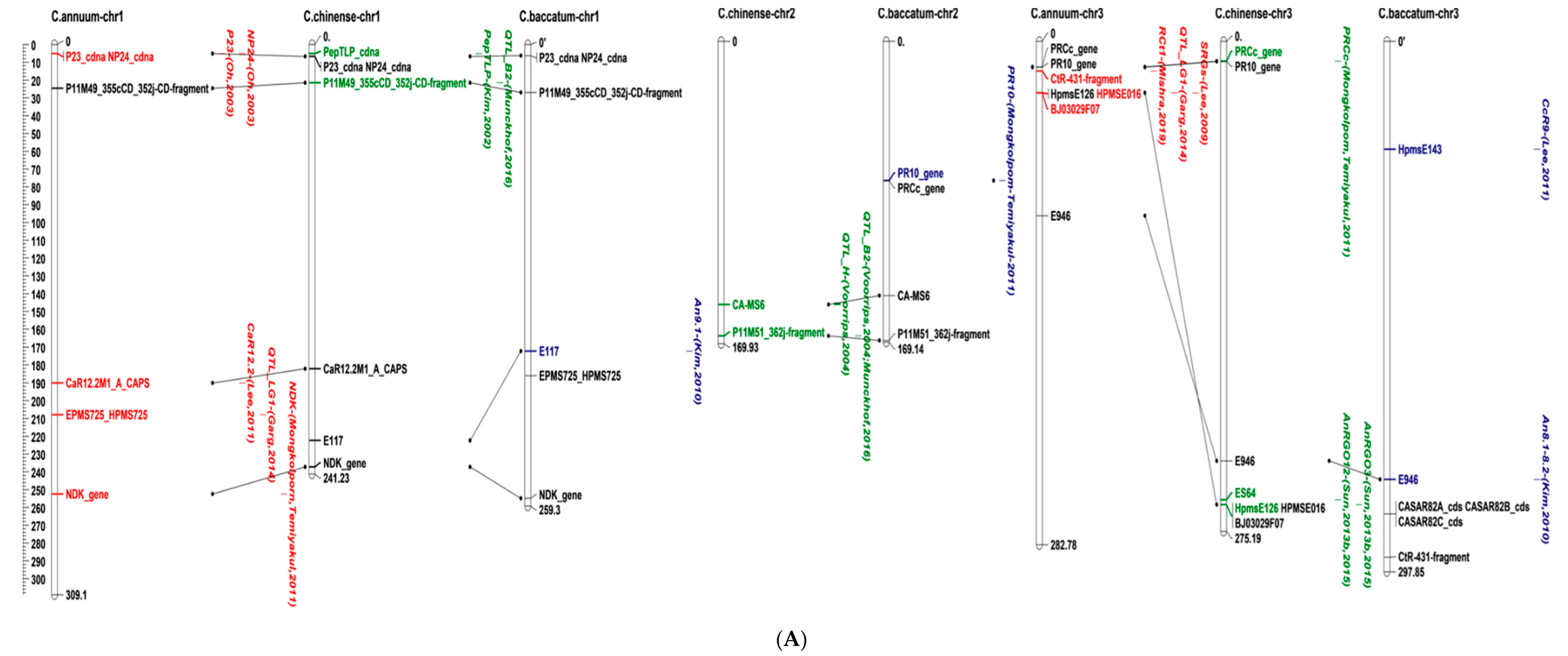
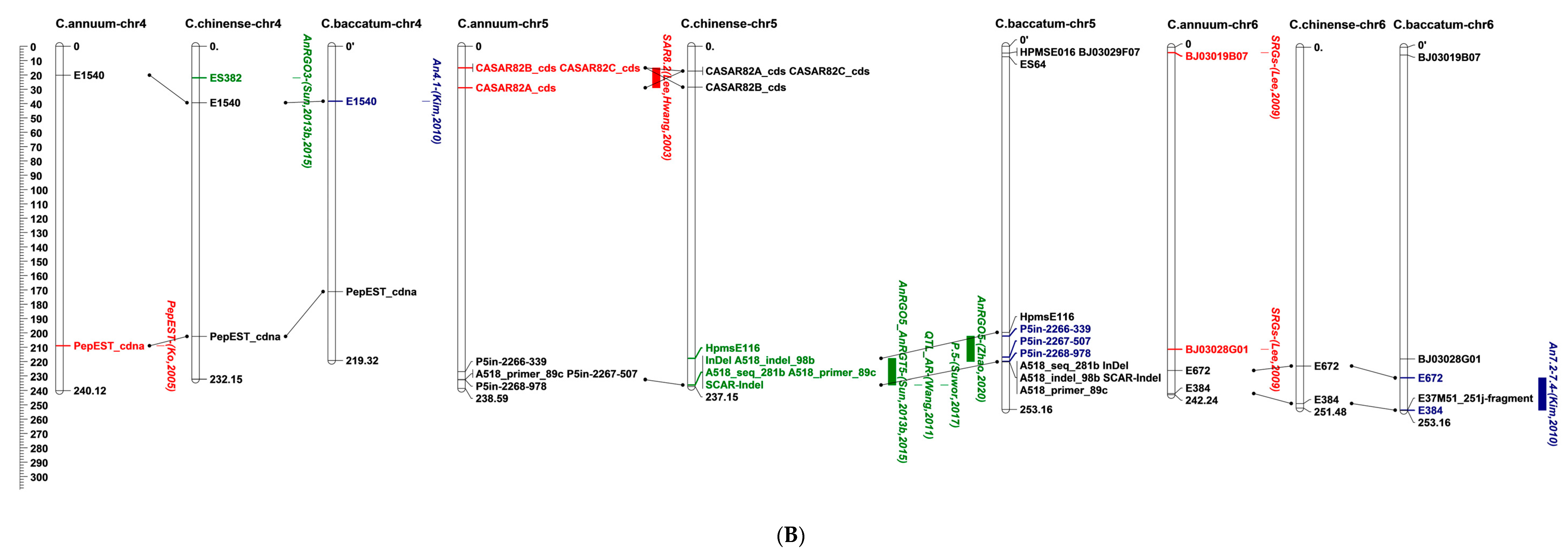
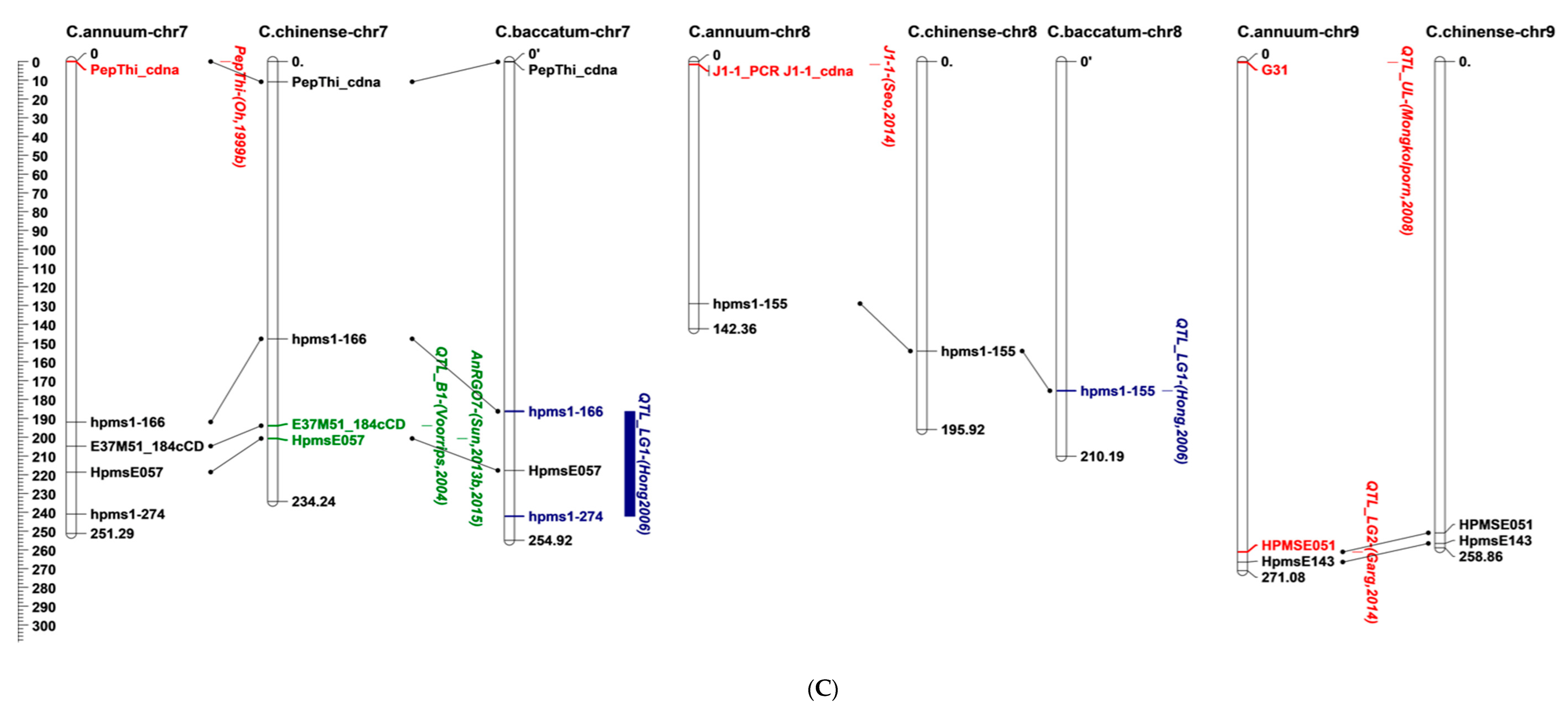
5. Location of Anthracnose Resistance on the Pepper Genome
| Study | QTL | Flanking Marker(s) | LOD a | R2 b | In Silico Mapping Chromosome | Colletotrichum Species | Resistant Source | Fruit Stage c | Trait d | |
|---|---|---|---|---|---|---|---|---|---|---|
| Capsicum Species | Accession | |||||||||
| [25] | B1 | E37M51_184cCD | 3.9–9.3 | 16.3–77.1 | 7 | C. capsici, C. scovillei | C. chinense | PRI95030 | F | LA, IF |
| B2 | P11M49_355cCD_352jCD | 2.7–4.7 | 30.2–51.8 | 1 | C. scovillei | |||||
| P11M51_362j | 2 | |||||||||
| E37M51_184cCD | 7 | |||||||||
| E37M51_251j | 10 | |||||||||
| P11M51_280 | 12 | |||||||||
| H1 | P11M51_362j | 2.9–3.7 | 13.4–20.5 | 2 | ||||||
| G1 | P11M50_137j, P14M58_199jCD | 2.8 | 7.6 | - | C. capsici | LD | ||||
| [85] | QTL_LG1 | hpms1-166, hpms1-274 | 7 | C. scovillei | C. baccatum | PBC81 | GF, RF | DI, TLD | ||
| hpms1-155 | 8 | |||||||||
| QTL_LG7 | hpms2-24 | - | ||||||||
| QTL_LG9 | hpms1-216, hpms1-274 | - | ||||||||
| QTL_LG10 | AF039662 | - | ||||||||
| [84] | QTL-UL | G31 | 9 | C. capsici | C. chinense | PBC932 | GF | LA | ||
| QTL_L4 | G28 | 11 | ||||||||
| QTL_L3 | G41 | - | ||||||||
| [33] | An9.1 | E117 | 4.6 | 14.4 | 1 | C. scovillei | C. baccatum var. pendulum | 881045 | F | DA |
| An8.1-8.2 | E946 | 4–4.1 | 2–8.4 | 3 | ||||||
| An8.1-8.2 | E946 | 4–4.1 | 2–8.4 | 3 | DR | |||||
| An4.1 | E1540 | 3.1 | 0.6 | 4 | - | |||||
| An7.2-7.4 | E672, E384 | 2.5–3.8 | 4.9–37.5 | 6 | DR | |||||
| An13.1 | E246 | 2.9 | 15.4 | 10 | DA | |||||
| An3.1-3.3 | E63M82_270, E65M81_450 | 2.7–3.6 | 12.8–30.1 | - | DR | |||||
| An5.1 | E74M79_370, E77M81_320 | 2.9 | 3.9 | - | ||||||
| [33] | An6.1 | me02em03_04, me02em02_05 | 2.8 | 0.9 | - | C. scovillei | C. baccatum var. pendulum | F | DR | |
| An7.1 | E76M83_250, E65M80_420 | 3.4 | 11.3 | - | DA | |||||
| An7.5 | E77M83_320, E75M83_450 | 3.9 | 6.9 | - | DR | |||||
| An8.3 | E70M84_380, E71M79_295 | 3.3 | 15.2 | - | ||||||
| An8.4 | E71M79_330, E67M79_250 | 3.8 | 20.5 | - | ||||||
| An9.2 | E76M83_350, E75M81_480 | 3 | 10.8 | - | DA | |||||
| [29] | CaR12.2 | CaR12.2M1_A_CAPS | 7.8–9.6 | 11.9–20.5 | 1 | C. scovillei | C. baccatum | PBC81 | GF, RF | TLD |
| CaR12.2M1_B_CAPS | 12 | |||||||||
| CcR9 | HpmsE143 | 13.4–15.9 | 57.5–78.9 | 3 | C. capsici | DI, TLD, OLD | ||||
| CaR12.1 | EtagMcag11, EtccMcga05 | 4.7 | 17.9 | - | C. scovillei | RF | TLD | |||
| CcRC | EaacMcgc02, EaatMccg07 | 6.7 | 10.6 | - | C. capsici | DI | ||||
| [77] | QTL_LG1 | EPMS725_HPMS725 | 3.4–5.8 | 16.7–71 | 1 | C. scovillei | C. annuum | Punjab Lal | GF, RF | IP |
| HPMSE016 | 3 | |||||||||
| QTL_LG2 | HPMSE051 | 2.1–4.9 | 7.2–18 | 9 | ||||||
| QCcR-ifp-iivr-1.1 QCcG-la.iivr1.1 | CAMS020, HPMSE016 HPMS725, CAMS644 | 3.5 | 14.2 | - | C. capsici | IP, LA | ||||
| [80] | QTL_AR | A518_InDel_98b, A518_seq_281b, A518_primer_89c | 5 | C. scovillei | C. chinense | PBC932 | GF | OLD | ||
| [15] | AnRGO3 | HpmsE126 | 2.3 | 2.9 | 3 | C. scovillei | C. chinense | PBC932 | GF | OLD |
| ES382 | 4 | |||||||||
| AnRGO12 | ES64, Epms745 | 2.7 | 3.1 | 3 | ||||||
| AnRGO5_AnRGT5 | InDel-HpmsE116, InDel | 31.9–32.3 | 60.5–62.4 | 5 | OLD, TLD | |||||
| AnRGO7 | HpmsE057 | 2.2 | 2.5 | 7 | OLD | |||||
| AnRGO10, AnRGD10 | C2_At4g03400, Gp20068 | 2.2–2.3 | 2.9–4.7 | 10 | OLD, DI | |||||
| AnRGD5, AnRRO5, AnRRT5, AnRRD5 | HpmsE116 | 2.7–12.3 | 9.3–33.2 | - | GF, RF | DI, OLD, TLD | ||||
| AnRGD12 | ES118, ES181 | 2.8 | 5.4 | - | GF | DI | ||||
| [81] | P.5 | SCAR-InDel | 5 | C. scovillei | C. chinense | PBC932 | GF | LD | ||
| LG12 | SSR-HpmsE032 | 12 | C. baccatum | PBC80 | ||||||
| [82] | AnRGO5 | P5in-2266-404, P5in-2268-978 | 24.4 | 69.3 | 5 | C. scovillei | C. chinense | PBC932 | GF | TLD |
| [72] | RA80f6_r1 | SNP_305/331 | 4 | 17.7 | 4 | C. scovillei | C. baccatum | PBC81 | GF, RF | IP |
| RA80f6_g1 | SNP_541/571 | 5.2 | 20.2 | 8 | ||||||
| RA80f6_g2 | SNP_228/218 | 3.5 | 12.8 | 3 | ||||||




6. Defense Mechanisms of Anthracnose Resistance Caused by Colletotrichum spp.
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- FAO. FAOSTAT. 2017. Available online: http://faostat.fao.org (accessed on 13 November 2021).
- Bosland, P.W.; Votava, E.J.; Votava, E.M. Peppers: Vegetable and spice capsicums. In Crop Production Science in Horticulture; CABI: Cambridge, UK, 2012; Volume 22. [Google Scholar]
- Ramchiary, N.; Kehie, M.; Brahma, V.; Kumaria, S.; Tandon, P. Application of genetics and genomics towards Capsicum translational research. Plant Biotechnol. Rep. 2013, 8, 101–123. [Google Scholar] [CrossRef]
- Kim, S.; Park, M.; Yeom, S.-I.; Kim, Y.-M.; Lee, J.M.; Lee, H.-A.; Seo, E.; Choi, J.; Cheong, K.; Kim, K.-T.; et al. Genome sequence of the hot pepper provides insights into the evolution of pungency in Capsicum species. Nat. Genet. 2014, 46, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Park, J.; Yeom, S.-I.; Kim, Y.-M.; Seo, E.; Kim, K.-T.; Kim, M.-S.; Lee, J.M.; Cheong, K.; Shin, H.-S.; et al. New reference genome sequences of hot pepper reveal the massive evolution of plant disease-resistance genes by retroduplication. Genome Biol. 2017, 18, 210. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Tabata, S.; Hirakawa, H.; Asamizu, E.; Shirasawa, K.; Isobe, S.; Kaneko, T.; Nakamura, Y.; Shibata, D.; Aoki, K.; et al. The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635–641. [Google Scholar] [CrossRef]
- Potato Genome Sequencing Consortium. Genome sequence and analysis of the tuber crop potato. Nature 2011, 475, 189. [Google Scholar] [CrossRef]
- Qin, C.; Yu, C.; Shen, Y.; Fang, X.; Chen, L.; Min, J.; Cheng, J.; Zhao, S.; Xu, M.; Luo, Y.; et al. Whole-genome sequencing of cultivated and wild peppers provides insights into Capsicum domestication and specialization. Proc. Natl. Acad. Sci. USA 2014, 111, 5135–5140. [Google Scholar] [CrossRef]
- Amusa, N.; Kehinde, I.; Adegbite, A. Pepper (Capsicum frutescens) fruit anthracnose in the humid forest region of south-western Nigeria. Nutr. Food Sci. 2004, 34, 130–134. [Google Scholar] [CrossRef]
- Sang, M.K.; Shrestha, A.; Kim, D.-Y.; Park, K.; Pak, C.H.; Kim, K.D. Biocontrol of Phytophthora blight and anthracnose in pepper by sequentially selected antagonistic rhizobacteria against Phytophthora capsici. Plant Pathol. J. 2013, 29, 154–167. [Google Scholar] [CrossRef]
- Oh, I.S.; In, M.S.; Woo, I.S.; Lee, S.K.; Yu, S.H. Anthracnose of pepper seedling caused by Colletotrichum coccodes (Wallr.) Hughes. Korean J. Mycol. 1988, 16, 151–156. [Google Scholar]
- Kim, K.-H.; Yoon, J.-B.; Park, H.-G.; Park, E.W.; Kim, Y.H.; Kim, J.-B.Y.K.-H.; Mongkolporn, O.; Taylor, P.W.J.; Yan, G.; Baidoo, R.; et al. Structural modifications and programmed cell death of chili pepper fruit related to resistance responses to Colletotrichum gloeosporioides infection. Phytopathology 2004, 94, 1295–1304. [Google Scholar] [CrossRef]
- Kim, S.H.; Yoon, J.B.; Do, J.W.; Park, H.G. Resistance to anthracnose caused by Colletotrichum acutatum in chili pepper (Capsicum annuum L.). J. Crop Sci. Biotechnol. 2007, 10, 277–280. [Google Scholar]
- Madhavan, S.; Paranidharan, V.; Velazhahan, R. RAPD and virulence analyses of Colletotrichum capsici isolates from chilli (Capsicum annuum). J. Plant Dis. Prot. 2010, 117, 253–257. [Google Scholar] [CrossRef]
- Sun, C.; Mao, S.L.; Zhang, Z.H.; Palloix, A.; Wang, L.H.; Zhang, B.X. Resistances to anthracnose (Colletotrichum acutatum) of Capsicum mature green and ripe fruit are controlled by a major dominant cluster of QTLs on chromosome P5. Sci. Hortic. 2015, 181, 81–88. [Google Scholar] [CrossRef]
- de Almeida, C.L.P.; Bento, C.D.S.; Sudré, C.P.; Pimenta, S.; Gonçalves, L.S.A.; Rodrigues, R. Genotype-Ideotype distance index and multivariate analysis to select sources of anthracnose resistance in Capsicum spp. Eur. J. Plant Pathol. 2019, 156, 223–236. [Google Scholar] [CrossRef]
- Oo, M.M.; Oh, S.-K. First report of anthracnose of chili pepper fruit caused by Colletotrichum truncatum in Korea. Plant Dis. 2020, 104, 564. [Google Scholar] [CrossRef]
- Mongkolporn, O.; Taylor, P.W.J. Chili anthracnose: Colletotrichum taxonomy and pathogenicity. Plant Pathol. 2018, 67, 1255–1263. [Google Scholar] [CrossRef]
- Mongkolporn, O. Anthracnose disease in Capsicum. In Capsicum: Breeding Strategies for Anthracnose Resistance; CRC Press: Boca Raton, FL, USA, 2018; pp. 47–71. [Google Scholar]
- Saxena, A.; Raghuwanshi, R.; Gupta, V.K.; Singh, H.B. Chilli anthracnose: The epidemiology and management. Front. Microbiol. 2016, 7, 1527. [Google Scholar] [CrossRef]
- Pakdeevaraporn, P.; Wasee, S.; Taylor, P.W.J.; Mongkolporn, O. Inheritance of resistance to anthracnose caused by Colleto-trichum capsici in Capsicum. Plant Breed. 2005, 124, 206–208. [Google Scholar] [CrossRef]
- Mahasuk, P.; Khumpeng, N.; Wasee, S.; Taylor, P.W.J.; Mongkolporn, O. Inheritance of resistance to anthracnose (Colleto-trichum capsici) at seedling and fruiting stages in chili pepper (Capsicum spp.). Plant Breed. 2009, 128, 701–706. [Google Scholar] [CrossRef]
- Marvel, J.K. Biology and control of pepper anthracnose. Ph.D. Thesis, Virginia Tech, Blacksburg, VA, USA, 2003. [Google Scholar]
- Than, P.P.; Prihastuti, H.; Phoulivong, S.; Taylor, P.W.J.; Hyde, K.D. Chilli anthracnose disease caused by Colletotrichum species. J. Zhejiang Univ. Sci. B 2008, 9, 764–778. [Google Scholar] [CrossRef]
- Voorrips, R.E.; Finkers, R.; Sanjaya, L.; Groenwold, R. QTL mapping of anthracnose (Colletotrichum spp.) resistance in a cross between Capsicum annuum and C. chinense. Theor. Appl. Genet. 2004, 109, 1275–1282. [Google Scholar] [CrossRef]
- Kim, S.H.; Yoon, J.B.; Do, J.W.; Park, H.G. A major recessive gene associated with anthracnose resistance to Colletotrichum capsici in chili pepper (Capsicum annuum L.). Breed. Sci. 2008, 58, 137–141. [Google Scholar] [CrossRef]
- Mongkolporn, O.; Taylor, P.W.J. Capsicum. In Wild Crop Relatives: Genomic and Breeding Resources; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 43–57. [Google Scholar]
- Mahasuk, P.; Taylor, P.W.J.; Mongkolporn, O. Identification of two new genes conferring resistance to Colletotrichum acutatum in Capsicum baccatum L. Phytopathology 2009, 99, 1100–1104. [Google Scholar] [CrossRef]
- Lee, J.; Hong, J.-H.; Do, J.W.; Yoon, J.B. Identification of QTLs for resistance to anthracnose to two Colletotrichum species in pepper. J. Crop. Sci. Biotechnol. 2010, 13, 227–233. [Google Scholar] [CrossRef]
- Kim, S.-G.; Ro, N.-Y.; Hur, O.-S.; Ko, H.-C.; Gwag, J.-G.; Huh, Y.-C. Evaluation of resistance to Colletotrichum acutatum in pepper genetic resources. Res. Plant Dis. 2012, 18, 93–100. [Google Scholar] [CrossRef]
- Garg, R.; Kumar, S.; Kumar, R.; Loganathan, M.; Saha, S.; Kumar, S.; Rai, A.B.; Roy, B.K. Novel source of resistance and differential reactions on chilli fruit infected by Colletotrichum capsici. Australas. Plant Pathol. 2012, 42, 227–233. [Google Scholar] [CrossRef]
- Yoon, J.; Park, H. Trispecies bridge crosses, (Capsicum annuum × C. chinense) × C. baccatum, as an alternative for introgression of anthracnose resistance from C. baccatum into C. annuum. Hortic. Environ. Biotechnol. 2005, 46, 5–9. [Google Scholar]
- Kim, S.; Kim, K.T.; Yang, E.Y.; Cho, M.C.; Jamal, A.; Chae, Y.; Pae, D.H.; Hwang, J.K. Identification of quantitative trait loci associated with anthracnose resistance in chili pepper (Capsicum spp.). Korean J. Hortic. Sci. Technol. 2010, 28, 1014–1024. [Google Scholar]
- Mahasuk, P.; Struss, D.; Mongkolporn, O. QTLs for resistance to anthracnose identified in two Capsicum sources. Mol. Breed. 2016, 36, 1–10. [Google Scholar] [CrossRef]
- Chang, S.H.; Chung, B.K. Studies on the varietal resistance and effects of nutrients for fungal growth of pepper anthracnose disease caused by Colletotrichum dematium fsp. capsicum. Korean J. Mycol. 1985, 13, 227–233. [Google Scholar]
- Souza, L.C.S.; Assis, L.A.G.; Catarino, A.D.M.; Hanada, R. Screening of chilli pepper genotypes against anthracnose (Colletotrichum brevisporum). Emir. J. Food Agric. 2020, 31, 919–929. [Google Scholar] [CrossRef]
- De Silva, D.D.; Ades, P.K.; Crous, P.W.; Taylor, P.W.J. Colletotrichum species associated with chili anthracnose in Australia. Plant Pathol. 2017, 66, 254–267. [Google Scholar] [CrossRef]
- Auyong, A.S.M.; Ford, R.; Taylor, P.W.J. The role of cutinase and its impact on pathogenicity of Colletotrichum truncatum. J. Plant Pathol. Microbiol. 2015, 6, 259. [Google Scholar] [CrossRef]
- Angadi, H.D.; Naik, M.K.; Patil, M.G. Evaluation of chilli genotypes against anthracnose disease. Veg. Sci. 2003, 30, 164–165. [Google Scholar]
- Vanan, T.; Khirbat, S.K.; Mehra, R. Reaction of detached fruits of chilli (Capsicum annuum L.) varieties to isolates of Colleto-trichum capsici (Syd.) Butler and Bisby. J. Spices Aromat. Crops. 2005, 14, 145–147. [Google Scholar]
- Giacomin, R.M.; Ruas, C.D.F.; Moreira, A.F.P.; Guidone, G.H.M.; Baba, V.Y.; Rodrigues, R.; Gonçalves, L.S.A. Inheritance of anthracnose resistance (Colletotrichum scovillei) in ripe and unripe Capsicum annuum fruits. J. Phytopathol. 2020, 168, 184–192. [Google Scholar] [CrossRef]
- Kanchana-udomkan, C.; Taylor, P.W.J.; Mongkolporn, O. Development of a bioassay to study anthracnose infection of chili fruit caused by Colletotrichum capsici. Thai J. Agric. Sci. 2004, 37, 293–297. [Google Scholar]
- Mahasuk, P.; Chinthaisong, J.; Mongkolporn, O. Differential resistances to anthracnose in Capsicum baccatum as responding to two Colletotrichum pathotypes and inoculation methods. Breed. Sci. 2013, 63, 333–338. [Google Scholar] [CrossRef]
- Mongkolporn, O.; Montri, P.; Supakaew, T.; Taylor, P.W.J. Differential reactions on mature green and ripe chili fruit infected by three Colletotrichum spp. Plant Dis. 2010, 94, 306–310. [Google Scholar] [CrossRef]
- Hasyim, A.; Setiawati, W.; Sutarya, R. Screening for resistance to anthracnose caused by Colletotrichum acutatum in chili pepper (Capsicum annuum L.) in Kediri, East Java. Adv. Agric. Bot. 2014, 6, 104–118. [Google Scholar]
- Wasee, S.; Pongpisutta, R.; Patarapuwadol, S.; Saridnirun, P.; Tangchitsomkid, N. Pepper (Capsicum spp.) germplasm management under Thailand conditions. In Proceedings of the SEAVEG 2012 High Value Vegetables in Southeast Asia: Production, Supply and Demand, Chiang Mai, Thailand, 24–26 January 2012; Volume 44. [Google Scholar]
- Kim, S.H.; Yoon, J.B.; Park, H.G. Inheritance of anthracnose resistance in a new genetic resource, Capsicum baccatum PI594137. J. Crop. Sci. Biotechnol. 2008, 11, 13–16. [Google Scholar]
- Silva, S.A.M.; Rodrigues, R.; Gonçalves, L.S.; Sudré, C.P.; Bento, C.S.; Carmo, M.G.; Medeiros, A.M. Resistance in Capsicum spp. to anthracnose affected by different stages of fruit development during pre- and post-harvest. Trop. Plant Pathol. 2014, 39, 335–341. [Google Scholar] [CrossRef]
- Mishra, R.; Rout, E.; Joshi, R.K. Identification of resistant sources against anthracnose disease caused by Colletotrichum truncatum and Colletotrichum gloeosporioides in Capsicum annuum L. Proc. Natl. Acad. Sci. India Sect. B Boil. Sci. 2019, 89, 517–524. [Google Scholar] [CrossRef]
- Mishra, R.; Mohanty, J.N.; Mahanty, B.; Joshi, R.K. A single transcript CRISPR/Cas9 mediated mutagenesis of CaERF28 confers anthracnose resistance in chilli pepper (Capsicum annuum L.). Planta 2021, 254, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Prasath, D.; Ponnuswami, V.; Muralidharan, V. Source of resistance to anthracnose (Colletotrichum capsici) disease in Capsicum species. Indian J. Agric. Sci. 2007, 77, 473–474. [Google Scholar]
- AVRDC. Genetic enhancement: Pepper unit. In AVRDC Report 2004; Kalb, T., Ed.; Asian Vegetable Research and Development Center: Shanhua, Taiwan, 2004; pp. 41–66. [Google Scholar]
- Cho, M.C.; Lin, S.W.; Shieh, S.C.; Suwor, P.; Kim, S.Y.; Techawongstien, S.; Kumar, S. Revisiting anthracnose-resistant Capsicum germplasm: Preliminary field evaluation. In Breakthroughs in the Genetics and Breeding of Capsicum and Eggplant, Proceedings of the XV EUCARPIA Meeting on Genetics and Breeding of Capsicum and Eggplant, Torino, Italy, 2–4 September 2013; Lanteri, S., Rotino, G.L., Eds.; Comitato per L’organizzazione degli Eventi (COE) DISAFA; Università degli Studi di Torino: Torino, Italy, 2013; pp. 363–366. [Google Scholar]
- Hong, J.K.; Hwang, B.K. Influence of inoculum density, wetness duration, plant age, inoculation method, and cultivar resistance on infection of pepper plants by Colletotrichum coccodes. Plant Dis. 1998, 82, 1079–1083. [Google Scholar] [CrossRef]
- Baba, V.Y.; Constantino, L.V.; Ivamoto-Suzuki, S.T.; Moreira, A.F.P.; Madeira, T.B.; Nixdorf, S.L.; Rodrigues, R.; Gonçalves, L.S.A. Capsicum-Colletotrichum interaction: Identification of resistance sources and quantification of secondary metabolites in unripe and ripe fruits in response to anthracnose infection. Sci. Hortic. 2019, 246, 469–477. [Google Scholar] [CrossRef]
- AVRDC. Vegetables in cereal-based systems: Off-season tomato, pepper and eggplant. In AVRDC Report 1998; Stares, J., Ed.; Asian Vegetable Research and Development Center: Shanhua, Taiwan, 1999; pp. 2–36. [Google Scholar]
- Montri, P.; Taylor, P.W.J.; Mongkolporn, O. Pathotypes of Colletotrichum capsici, the causal agent of chili anthracnose, in Thailand. Plant Dis. 2009, 93, 17–20. [Google Scholar] [CrossRef]
- Padilha, H.K.M.; Madruga, N.D.A.; Aranha, B.C.; Hoffmann, J.F.; Crizel, R.L.; Barbieri, R.L.; Chaves, F.C. Defense responses of Capsicum spp. genotypes to post-harvest Colletotrichum sp. inoculation. Phytoparasitica 2019, 47, 557–573. [Google Scholar] [CrossRef]
- Park, H.K.; Kim, B.S.; Lee, W.S. Inheritance of resistance to anthracnose (Colletotrichum spp.) in pepper (Capsicum annuum L.) I. Genetic analysis of anthracnose resistance by diallel crosses. J. Korean Soc. Hortic. Sci. 1990, 31, 91–105. [Google Scholar]
- Ahmed, N.; Dey, S.K.; Hundal, J.S. Inheritance of resistance to anthracnose in chilli. Indian Phytopathol. 1991, 44, 402–403. [Google Scholar]
- Hirdaypal, S.; Jarnail, S.; Kaur, S. Resistant sources to fruit rot (Colletotrichum capsici) in pepper (Capsicum annuum L.). J. Res. Punjab Agric. Univ. 1990, 27, 419–420. [Google Scholar]
- Henz, G.P.; Boiteux, L.S.; Lima, M.F. Reaction of Capsicum spp. fruits to Colletotrichum gloeosporioides. Capsicum Eggplant Nwsl. 1993, 12, 79–80. [Google Scholar]
- AVRDC. Vegetables in cereal-based systems: Off-season tomato, pepper and eggplant. In AVRDC Report 1999; Koizumi, M., Abbass, D., Stares, J., Eds.; Asian Vegetable Research and Development Center: Shanhua, Taiwan, 2000; pp. 2–36. [Google Scholar]
- AVRDC. Annual Report 2013; AVRDC-The World Vegetable Center: Shanhua, Taiwan, 2014; pp. 14–27. [Google Scholar]
- Kaur, N.; Dhiman, J.S.; Khurana, D.S. Physiological and biochemical traits analysis of Capsicum annuum L. germplasm for resistance to Colletotrichum capsici. J. Cell Plant Sci. 2011, 2, 12–21. [Google Scholar]
- Pereira, M.J.Z.; Junior, N.S.M.; Sussel, A.A.B.; Sala, F.C.; de Costa, C.P.; Boiteux, L.S. Reação de acessos de Capsicum e de progênies de cruzamentos interespecíficos a isolados de Colletotrichum acutatum. Hortic. Bras. 2011, 29, 569–576. [Google Scholar] [CrossRef]
- Babu, B.S.; Pandravada, S.; Rao, R.P.; Anitha, K.; Chakrabarty, S.; Varaprasad, K. Global sources of pepper genetic resources against arthropods, nematodes and pathogens. Crop. Prot. 2011, 30, 389–400. [Google Scholar] [CrossRef]
- Garg, R.; Loganathan, M.; Saha, S.; Roy, B.K. Chilli anthracnose: A review of causal organism, resistance source and mapping of gene. In Microbial Diversity and Biotechnology in Food Security; Kharwar, R., Upadhyay, R., Dubey, N., Raghuwanshi, R., Eds.; Springer: New Delhi, India, 2014; pp. 589–610. [Google Scholar]
- Machenahalli, S.R.; Nargund, V.; Sridevi, O. Reaction of chilli genotypes against fruit rot complex. Int. J. Life Sci. 2016, 11, 715–719. [Google Scholar]
- Maracahipes, C.; Taveira, G.B.; Mello, E.O.; Carvalho, A.O.; Rodrigues, R.; Perales, J.; Teixeira-Ferreira, A.; Silva, M.S.; Rocha, G.L.; Fernandes, K.V.S.; et al. Biochemical analysis of antimicrobial peptides in two different Capsicum genotypes after fruit infection by Colletotrichum gloeosporioides. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef]
- Manzur, J.P.; Fita, A.; Prohens, J.; Rodríguez-Burruezo, A. Successful wide hybridization and introgression breeding in a diverse set of common peppers (Capsicum annuum) using different cultivated Ají (C. baccatum) accessions as donor parents. PLoS ONE 2015, 10, e0144142. [Google Scholar] [CrossRef]
- Kethom, W.; Mongkolporn, O. New QTLs for anthracnose resistance identified in Capsicum baccatum ‘PBC80’-derived recombinant inbred lines. Euphytica 2021, 217, 1–12. [Google Scholar] [CrossRef]
- Lin, Q.; Kanchana-udomkan, C.; Jaunet, T.; Mongkolporn, O. Genetic analysis of resistance to pepper anthracnose caused by Colletotrichum capsici. Thai J. Agric. Sci. 2002, 35, 259–264. [Google Scholar]
- Ridzuan, R.; Rafii, M.Y.; Ismail, S.I.; Yusoff, M.M.; Miah, G.; Usman, M. Breeding for anthracnose disease resistance in chili: Progress and prospects. Int. J. Mol. Sci. 2018, 19, 3122. [Google Scholar] [CrossRef]
- Setiawati, W.; Udiarto, B.K.; Soetiarso, T.A. The effect of variety and planting system of chili pepper on incidence of whiteflies. J. Hortic. 2008, 18, 55–61. [Google Scholar]
- Reddy, M.K.; Srivastava, A.; Kumar, S.; Kumar, R.; Chawda, N.; Ebert, A.W.; Vishwakarma, M. Chilli (Capsicum annuum L.) breeding in India: An overview. SABRAO J. Breed Genet. 2014, 46, 160–173. [Google Scholar]
- Garg, R. Genetics of Host Pathogen Interaction Resistance to Anthracnose in Chilli. Ph.D. Thesis, Banaras Hindu University, Uttar Pradesh, India, 2011. [Google Scholar]
- Yoon, J.B.; Do, J.W.; Kim, S.H.; Park, H.G. Inheritance of anthracnose (Colletotrichum acutatum) resistance in Capsicum using interspecific hybridization. Korean J. Agric. Sci. 2009, 27, 140–144. [Google Scholar]
- Chen, C.; Chen, H.; He, Y.; Xia, R. TBtools, a toolkit for biologists integrating various biological data handling tools with a user-friendly interface. BioRxiv 2018, 1, 289660. [Google Scholar]
- Wang, Y.W. Development of Sequence Characterized Amplified Region (SCAR) Markers Associated with Pepper Anthracnose (Colletotrichum acutatum) Resistance. Master’s Thesis, National Chiayi University, Chiayi, Taiwan, 2011. [Google Scholar]
- Suwor, P.; Sanitchon, J.; Thummabenjapone, P.; Kumar, S.; Techawongstien, S. Inheritance analysis of anthracnose resistance and marker-assisted selection in introgression populations of chili (Capsicum annuum L.). Sci. Hortic. 2017, 220, 20–26. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, Y.; Zhang, Z.; Cao, Y.; Yu, H.; Ma, W.; Zhang, B.; Wang, R.; Gao, J.; Wang, L. Fine mapping of the major anthracnose resistance QTL AnRGO5 in Capsicum chinense ‘PBC932’. BMC Plant Biol. 2020, 20, 189. [Google Scholar] [CrossRef]
- Sun, C.Y.; Mao, S.L.; Zhang, Z.H. QTLs analysis of anthracnose (Colletotrichum acutatum) resistance in pepper (Capsicum spp.). In Breakthroughs in the Genetics and Breeding of Capsicum and Eggplant, Proceedings of the XV EUCARPIA Meeting on Genetics and Breeding of Capsicum and Eggplant, Torino, Italy, 2–4 September 2013; Lanteri, S., Rotino, G.L., Eds.; Comitato per L’organizzazione degli Eventi (COE) DISAFA; Università degli Studi di Torino: Torino, Italy, 2013; pp. 169–176. [Google Scholar]
- Mongkolporn, O. Mapping genes conferring resistance to anthracnose (Colletotrichum capsici) and development of molecular markers for the selection of anthracnose resistance trait in chili. 2008; unpublished. [Google Scholar]
- Hong, J.H.; Do, J.W.; Park, H.G. Species-Specific QTLs Associated with Anthracnose Resistance to Different Colletotrichum spp. in an Introgressed BC1F2 Population from Capsicum annuum × C. baccatum. Master’s Thesis, Seoul National University, Seoul, Republic of Korea, 2006. [Google Scholar]
- Lee, J.; Do, J.W.; Yoon, J.B. Development of STS markers linked to the major QTLs for resistance to the pepper anthracnose caused by Colletotrichum acutatum and C. capsici. Hortic. Environ. Biotechnol. 2011, 52, 596–601. [Google Scholar] [CrossRef]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed]
- Ranathunge, N.P.; Mongkolporn, O.; Ford, R.; Taylor, P.W.J. Colletotrichum truncatum pathosystem on Capsicum spp.: Infection, colonization and defence mechanisms. Australas. Plant Pathol. 2012, 41, 463–473. [Google Scholar] [CrossRef]
- Mishra, R.; Nanda, S.; Rout, E.; Chand, S.K.; Mohanty, J.N.; Joshi, R.K. Differential expression of defense-related genes in chilli pepper infected with anthracnose pathogen Colletotrichum truncatum. Physiol. Mol. Plant Pathol. 2017, 97, 1–10. [Google Scholar] [CrossRef]
- Soh, H.C.; Park, A.R.; Park, S.; Back, K.; Yoon, J.B.; Park, H.G.; Kim, Y.S. Comparative analysis of pathogenesis-related protein 10 (PR10) genes between fungal resistant and susceptible peppers. Eur. J. Plant Pathol. 2011, 132, 37–48. [Google Scholar] [CrossRef]
- Oh, B.-J.; Ko, M.K.; Kostenyuk, I.; Shin, B.; Kim, K.S. Coexpression of a defensin gene and a thionin-like gene via different signal transduction pathways in pepper and Colletotrichum gloeosporioides interactions. Plant Mol. Biol. 1999, 41, 313–319. [Google Scholar] [CrossRef]
- Oh, B.-J.; Ko, M.-K.; Kim, K.S.; Kim, Y.S.; Lee, H.H.; Jeon, W.B.; Im, K.H. Isolation of defense-related genes differentially expressed in the resistance interaction between pepper fruits and the anthracnose fungus Colletotrichum gloeosporioides. Mol. Cells 2003, 15, 349–355. [Google Scholar]
- Ko, M.K.; Jeon, W.B.; Kim, K.S.; Lee, H.H.; Seo, H.H.; Kim, Y.S.; Oh, B.-J. A Colletotrichum gloeosporioides-induced esterase gene of nonclimacteric pepper (Capsicum annuum) fruit during ripening plays a role in resistance against fungal infection. Plant Mol. Biol. 2005, 58, 529–541. [Google Scholar] [CrossRef]
- Lee, S.; Hong, J.-C.; Jeon, W.B.; Chung, Y.-S.; Sung, S.; Choi, D.; Joung, Y.H.; Oh, B.-J. The salicylic acid-induced protection of non-climacteric unripe pepper fruit against Colletotrichum gloeosporioides is similar to the resistance of ripe fruit. Plant Cell Rep. 2009, 28, 1573–1580. [Google Scholar] [CrossRef]
- Lee, S.; Hwang, B. Identification of the pepper SAR8.2 gene as a molecular marker for pathogen infection, abiotic elicitors and environmental stresses in Capsicum annuum. Planta 2003, 216, 387–396. [Google Scholar] [CrossRef]
- Yi, G.; Lee, J.M.; Lee, S.; Choi, D.; Kim, B.-D. Exploitation of pepper EST–SSRs and an SSR-based linkage map. Theor. Appl. Genet. 2006, 114, 113–130. [Google Scholar] [CrossRef]
- Portis, E.; Nagy, I.; Sasvári, Z.; Stágel, A.; Barchi, L.; Lanteri, S. The design of Capsicum spp. SSR assays via analysis of in silico DNA sequence, and their potential utility for genetic mapping. Plant Sci. 2007, 172, 640–648. [Google Scholar] [CrossRef]
- Minamiyama, Y.; Tsuro, M.; Hirai, M. An SSR-based linkage map of Capsicum annuum. Mol. Breed. 2006, 18, 157–169. [Google Scholar] [CrossRef]
- Lee, J.M.; Nahm, S.H.; Kim, Y.M.; Kim, B.D. Characterization and molecular genetic mapping of microsatellite loci in pepper. Theor. Appl. Genet. 2003, 108, 619–627. [Google Scholar] [CrossRef]
- Li, G.; Quiros, C.F. Sequence-related amplified polymorphism (SRAP), a new marker system based on a simple PCR reaction: Its application to mapping and gene tagging in Brassica. Theor. Appl. Genet. 2001, 103, 455–461. [Google Scholar] [CrossRef]
- Kim, Y.S.; Park, J.Y.; Kim, K.S.; Ko, M.K.; Cheong, S.J.; Oh, B.-J. A thaumatin-like gene in nonclimacteric pepper fruits used as molecular marker in probing disease resistance, ripening, and sugar accumulation. Plant Mol. Biol. 2002, 49, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.H.; Park, S.; Park, S.; Oh, B.J.; Back, K.; Han, O.; Kim, J.; Kim, Y.S. Overexpression of a defensin enhances resistance to a fruit-specific anthracnose fungus in pepper. PLoS ONE 2014, 21, e97936. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, S.H.; Park, H.G.; Yoon, J.B. Capsicum germplasm resistant to pepper anthracnose differentially interact with Colletotrichum isolates. Hortic. Environ. Biotechnol. 2009, 50, 17–23. [Google Scholar]

| Colletotrichum Species a | Capsicum Species b | |||||
|---|---|---|---|---|---|---|
| C. annuum | C. baccatum | C. chacoense | C. chinense | C. frutescens | Capsicum spp. | |
| C. scovillei | 18 | 46 | 4 | 21 | 7 | - |
| C. capsici | 76 | 22 | - | 13 | 8 | 4 |
| C. siamense | 31 | 18 | - | 41 | - | 15 |
| C. coccodes | 1 | - | - | - | 1 | - |
| C. brevisporum | - | - | - | 15 | - | - |
| C. dematium | 7 | - | - | - | - | - |
| C. truncatum | 9 | - | - | - | - | - |
| Colletotrichum spp. | 18 | - | - | - | - | - |
| Total | 160 | 86 | 4 | 90 | 16 | 19 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, L.; van den Munckhof, M.C.; Bai, Y.; Voorrips, R.E. Resistance to Anthracnose Rot Disease in Capsicum. Agronomy 2023, 13, 1434. https://doi.org/10.3390/agronomy13051434
Cui L, van den Munckhof MC, Bai Y, Voorrips RE. Resistance to Anthracnose Rot Disease in Capsicum. Agronomy. 2023; 13(5):1434. https://doi.org/10.3390/agronomy13051434
Chicago/Turabian StyleCui, Lei, Michiel C. van den Munckhof, Yuling Bai, and Roeland E. Voorrips. 2023. "Resistance to Anthracnose Rot Disease in Capsicum" Agronomy 13, no. 5: 1434. https://doi.org/10.3390/agronomy13051434
APA StyleCui, L., van den Munckhof, M. C., Bai, Y., & Voorrips, R. E. (2023). Resistance to Anthracnose Rot Disease in Capsicum. Agronomy, 13(5), 1434. https://doi.org/10.3390/agronomy13051434