
Genome-Wide Association Study (GWAS) of the Agronomic Traits and Phenolic Content in Sorghum (Sorghum bicolor L.) Genotypes
 , ,
, ,  , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and DNA Extraction
2.2. Evaluation of Agronomic Traits and Soluble Solids Content
2.3. Ultra-High-Performance Liquid Chromatography (UPLC) Analysis
2.4. Genotyping-by-Sequencing Analysis
2.5. Genome-Wide Association Study (GWAS) with Agronomic Traits and Phenolic Compounds
3. Results
3.1. Subsection Agronomic Traits in Sorghum Genotypes
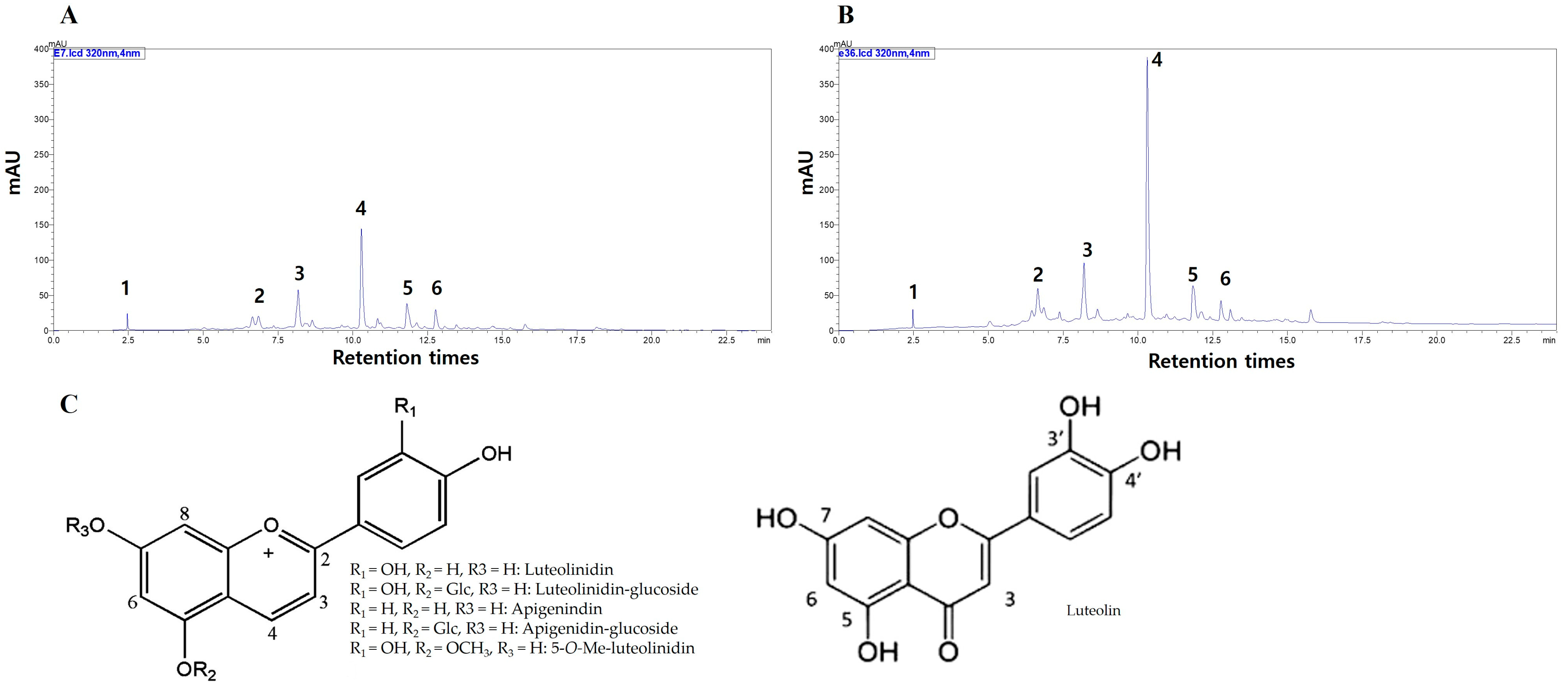
3.2. UPLC Analysis in Sorghum Genotypes
3.3. Correlation Analysis
3.4. Genotyping-by-Sequencing of Sorghum Genotypes
3.5. Identification of SNPs
3.6. GWAS Analysis for Agronomic Traits
3.7. GWAS Analysis for Total Phenolic Content
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Habyarimana, E.; Laureti, D.; De Ninno, M.; Lorenzoni, C. Performances of biomass sorghum [Sorghum bicolor (L.) Moench] under different water regimes in Mediterranean region. Ind. Crops Prod. 2004, 20, 23–28. [Google Scholar] [CrossRef]
- Shiringani, A.L.; Friedt, W. QTL for fibre-related traits in grain × sweet sorghum as a tool for the enhancement of sorghum as a biomass crop. Theor. Appl. Genet. 2011, 123, 999–1011. [Google Scholar] [CrossRef] [PubMed]
- Habyarimana, E.; De Franceschi, P.; Ercisli, S.; Baloch, F.S.; Dall’Agata, M. Genome-wide association study for biomass related traits in a panel of Sorghum bicolor and S. bicolor × S. halepense populations. Front. Plant Sci. 2020, 11, 551305. [Google Scholar] [CrossRef]
- Kawahigashi, H.; Kasuga, S.; Okuizumi, H.; Hiradate, S.; Yonemaru, J.i. Evaluation of Brix and sugar content in stem juice from sorghum varieties. Grassl. Sci. 2013, 59, 11–19. [Google Scholar] [CrossRef]
- Shukla, S.; Felderhoff, T.J.; Saballos, A.; Vermerris, W. The relationship between plant height and sugar accumulation in the stems of sweet sorghum (Sorghum bicolor (L.) Moench). Field Crops Res. 2017, 203, 181–191. [Google Scholar] [CrossRef]
- Sabadin, P.; Malosetti, M.; Boer, M.; Tardin, F.; Santos, F.; Guimaraes, C.; Gomide, R.; Andrade, C.; Albuquerque, P.; Caniato, F. Studying the genetic basis of drought tolerance in sorghum by managed stress trials and adjustments for phenological and plant height differences. Theor. Appl. Genet. 2012, 124, 1389–1402. [Google Scholar] [CrossRef] [PubMed]
- Soleymani, A.; Shahrajabian, M.H.; Esfahan, I. The Effects of Fe, Mn and Zn Foliar Application on Yield, Ash and Protein Percentage of Forage Sorghum in Climatic Condition of Esfahan. Int. J. Biol. 2012, 4, 92. [Google Scholar] [CrossRef] [Green Version]
- Hassan, M.U.; Chattha, M.U.; Barbanti, L.; Mahmood, A.; Chattha, M.B.; Khan, I.; Mirza, S.; Aziz, S.A.; Nawaz, M.; Aamer, M. Cultivar and seeding time role in sorghum to optimize biomass and methane yield under warm dry climate. Ind. Crops Prod. 2020, 145, 111983. [Google Scholar] [CrossRef]
- Kim, H.-J.; Woo, K.S.; Lee, J.Y.; Choe, M.E.; Lee, H.; Lee, Y.-Y.; Lee, B.W.; Kim, M.; Kang, M.S. Physicochemical properties, functional components, and physiological activities of sorghum cultivars. Korean J. Food Sci. Technol. 2020, 49, 1349–1356. [Google Scholar] [CrossRef]
- Bado, S.; Forster, B.P.; Nielen, S.; Ali, A.M.; Lagoda, P.J.; Till, B.J.; Laimer, M. Plant mutation breeding: Current progress and future assessment. Plant Breed. Rev. 2015, 39, 23–88. [Google Scholar]
- Johnson, R.E.; Washington, M.T.; Prakash, S.; Prakash, L. Fidelity of human DNA polymerase η. J. Biol. Chem. 2000, 275, 7447–7450. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.-Y.; Ramachandran, S. Natural and artificial mutants as valuable resources for functional genomics and molecular breeding. Int. J. Biol. Sci. 2010, 6, 228–251. [Google Scholar] [CrossRef]
- Martin, J. Understanding gamma sterilization. Biopharm Int. 2012, 25, 18. [Google Scholar]
- Djezzar, B. Ionizing Radiation Effects and Applications; IntechOpenLo: London, UK, 2018. [Google Scholar]
- Malek, M.; Rafii, M.Y.; Afroz, S.S.; Nath, U.K.; Mondal, M. Morphological characterization and assessment of genetic variability, character association, and divergence in soybean mutants. Sci. World J. 2014, 2014, 968796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhary, J.; Alisha, A.; Bhatt, V.; Chandanshive, S.; Kumar, N.; Mir, Z.; Kumar, A.; Yadav, S.K.; Shivaraj, S.; Sonah, H. Mutation breeding in tomato: Advances, applicability and challenges. Plants 2019, 8, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girard, A.L.; Awika, J.M. Sorghum polyphenols and other bioactive components as functional and health promoting food ingredients. J. Cereal Sci. 2018, 84, 112–124. [Google Scholar] [CrossRef]
- de Morais Cardoso, L.; Pinheiro, S.S.; Martino, H.S.D.; Pinheiro-Sant’Ana, H.M. Sorghum (Sorghum bicolor L.): Nutrients, bioactive compounds, and potential impact on human health. Crit. Rev. Food Sci. Nutr. 2017, 57, 372–390. [Google Scholar] [CrossRef]
- Pontieri, P.; Pepe, G.; Campiglia, P.; Merciai, F.; Basilicata, M.G.; Smolensky, D.; Calcagnile, M.; Troisi, J.; Romano, R.; Del Giudice, F. Comparison of Content in Phenolic Compounds and Antioxidant Capacity in Grains of White, Red, and Black Sorghum Varieties Grown in the Mediterranean Area. ACS Food Sci. Technol. 2021, 1, 1109–1119. [Google Scholar] [CrossRef]
- Obruca, S.; Benesova, P.; Marsalek, L.; Marova, I. Use of lignocellulosic materials for PHA production. Chem. Biochem. Eng. Q. 2015, 29, 135–144. [Google Scholar] [CrossRef]
- Lee, S.J.; Lee, J.H.; Yang, X.; Kim, S.B.; Lee, J.H.; Yoo, H.Y.; Park, C.; Kim, S.W. Phenolic compounds: Strong inhibitors derived from lignocellulosic hydrolysate for 2, 3-butanediol production by Enterobacter aerogenes. Biotechnol. J. 2015, 10, 1920–1928. [Google Scholar] [CrossRef]
- Dlamini, N.R.; Taylor, J.R.; Rooney, L.W. The effect of sorghum type and processing on the antioxidant properties of African sorghum-based foods. Food Chem. 2007, 105, 1412–1419. [Google Scholar] [CrossRef]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.; Guo, H.; Kong, W.; Chandnani, R.; Shuang, L.-S.; Paterson, A.H. Application of genotyping by sequencing technology to a variety of crop breeding programs. Plant Sci. 2016, 242, 14–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirschhorn, J.N.; Daly, M.J. Genome-wide association studies for common diseases and complex traits. Nat. Rev. Genet. 2005, 6, 95–108. [Google Scholar] [CrossRef]
- Cuevas, H.E.; Rosa-Valentin, G.; Hayes, C.M.; Rooney, W.L.; Hoffmann, L. Genomic characterization of a core set of the USDA-NPGS Ethiopian sorghum germplasm collection: Implications for germplasm conservation, evaluation, and utilization in crop improvement. BMC Genom. 2017, 18, 108. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Ersoz, E.; Lai, C.-Q.; Todhunter, R.J.; Tiwari, H.K.; Gore, M.A.; Bradbury, P.J.; Yu, J.; Arnett, D.K.; Ordovas, J.M. Mixed linear model approach adapted for genome-wide association studies. Nat. Genet. 2010, 42, 355–360. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.; Burke, J.; Chopra, R.; Burow, G.; Chen, J.; Wang, B.; Hayes, C.; Emendack, Y.; Ware, D.; Xin, Z. A sorghum mutant resource as an efficient platform for gene discovery in grasses. Plant Cell 2016, 28, 1551–1562. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.-G.; Lyu, J.I.; Kim, J.M.; Seo, J.S.; Choi, H.-I.; Jo, Y.D.; Kim, S.H.; Eom, S.H.; Ahn, J.-W.; Bae, C.-H. Identification of Loci Governing Agronomic Traits and Mutation Hotspots via a GBS-Based Genome-Wide Association Study in a Soybean Mutant Diversity Pool. Int. J. Mol. Sci. 2022, 23, 10441. [Google Scholar] [CrossRef]
- Ryu, J.; Lyu, J.I.; Kim, D.-G.; Koo, K.M.; Yang, B.; Jo, Y.D.; Kim, S.H.; Kwon, S.-J.; Ha, B.-K.; Kang, S.-Y. Single Nucleotide Polymorphism (SNP) discovery and association study of flowering times, crude fat and fatty acid composition in rapeseed (Brassica napus L.) mutant lines using Genotyping-by-Sequencing (GBS). Agronomy 2021, 11, 508. [Google Scholar] [CrossRef]
- Petti, C.; Kushwaha, R.; Tateno, M.; Harman-Ware, A.E.; Crocker, M.; Awika, J.; DeBolt, S. Mutagenesis breeding for increased 3-deoxyanthocyanidin accumulation in leaves of Sorghum bicolor (L.) Moench: A source of natural food pigment. J. Agric. Food Chem. 2014, 62, 1227–1232. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Cox, M.P.; Peterson, D.A.; Biggs, P.J. SolexaQA: At-a-glance quality assessment of Illumina second-generation sequencing data. BMC Bioinform. 2010, 11, 485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, E.A.; Brenton, Z.W.; Flinn, B.S.; Jenkins, J.; Shu, S.; Flowers, D.; Luo, F.; Wang, Y.; Xia, P.; Barry, K. A new reference genome for Sorghum bicolor reveals high levels of sequence similarity between sweet and grain genotypes: Implications for the genetics of sugar metabolism. BMC Genom. 2019, 20, 420. [Google Scholar] [CrossRef] [Green Version]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-E.; Oh, S.-K.; Lee, J.-H.; Lee, B.-M.; Jo, S.-H. Genome-wide SNP calling using next generation sequencing data in tomato. Mol. Cells 2014, 37, 36. [Google Scholar] [CrossRef] [PubMed]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [Green Version]
- Lipka, A.E.; Tian, F.; Wang, Q.; Peiffer, J.; Li, M.; Bradbury, P.J.; Gore, M.A.; Buckler, E.S.; Zhang, Z. GAPIT: Genome association and prediction integrated tool. Bioinformatics 2012, 28, 2397–2399. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Pressoir, G.; Briggs, W.H.; Vroh Bi, I.; Yamasaki, M.; Doebley, J.F.; McMullen, M.D.; Gaut, B.S.; Nielsen, D.M.; Holland, J.B. A unified mixed-model method for association mapping that accounts for multiple levels of relatedness. Nat. Genet. 2006, 38, 203–208. [Google Scholar] [CrossRef]
- Beyaz, R.; Yildiz, M. The use of gamma irradiation in plant mutation breeding. In Plant Engineering; InTech: Rijeka, Croatia, 2017; pp. 34–46. [Google Scholar]
- Micke, A.; Maluszynski, M.; Donini, B. Plant Cultivars Derived from Mutation Induction or the Use of Induced Mutants in Cross Breeding; IAEA: Vienna, Ausrtria, 1985. [Google Scholar]
- Human, S.; Indriatama, W.M. Sorghum improvement program by using mutation breeding in Indonesia. IOP Conf. Ser. Earth Environ. Sci. 2020, 484, 012003. [Google Scholar] [CrossRef]
- Madhusudhana, R.; Patil, J. A major QTL for plant height is linked with bloom locus in sorghum [Sorghum bicolor (L.) Moench]. Euphytica 2013, 191, 259–268. [Google Scholar] [CrossRef]
- Kong, W.; Jin, H.; Franks, C.D.; Kim, C.; Bandopadhyay, R.; Rana, M.K.; Auckland, S.A.; Goff, V.H.; Rainville, L.K.; Burow, G.B. Genetic analysis of recombinant inbred lines for Sorghum bicolor × Sorghum propinquum. G3 Genes Genomes Genet. 2013, 3, 101–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, S.C.; Sharma, A.; Rooney, W.L.; Klein, P.E.; Mullet, J.E.; Mitchell, S.E.; Kresovich, S. Genetic improvement of sorghum as a biofuel feedstock: I. QTL for stem sugar and grain nonstructural carbohydrates. Crop Sci. 2008, 48, 2165–2179. [Google Scholar] [CrossRef]
- Reddy, P.S. Breeding for Abiotic Stress Resistance in Sorghum; Elsevier: Amsterdam, The Netherlands, 2019; pp. 325–340. [Google Scholar]
- Xu, W.; Rosenow, D.; Nguyen, H. Stay green trait in grain sorghum: Relationship between visual rating and leaf chlorophyll concentration. Plant Breed. 2000, 119, 365–367. [Google Scholar] [CrossRef]
- Rama Reddy, N.R.; Ragimasalawada, M.; Sabbavarapu, M.M.; Nadoor, S.; Patil, J.V. Detection and validation of stay-green QTL in post-rainy sorghum involving widely adapted cultivar, M35-1 and a popular stay-green genotype B35. BMC Genom. 2014, 15, 909. [Google Scholar] [CrossRef] [Green Version]
- Kiranmayee, K.U.; Hash, C.T.; Sivasubramani, S.; Ramu, P.; Amindala, B.P.; Rathore, A.; Kishor, P.K.; Gupta, R.; Deshpande, S.P. Fine-mapping of sorghum stay-green QTL on chromosome10 revealed genes associated with delayed senescence. Genes 2020, 11, 1026. [Google Scholar] [CrossRef]
- Sun, X.-Z.; Minowa, T.; Yamaguchi, K.; Genchi, Y. Evaluation of energy consumption and greenhouse gas emissions from poly (phenyllactic acid) production using sweet sorghum. J. Clean. Prod. 2015, 87, 208–215. [Google Scholar] [CrossRef]
- Ebrahimian, F.; Karimi, K.; Kumar, R. Sustainable biofuels and bioplastic production from the organic fraction of municipal solid waste. Waste Manag. 2020, 116, 40–48. [Google Scholar] [CrossRef]
- Li, J.; Tang, W.; Zhang, Y.-W.; Chen, K.-N.; Wang, C.; Liu, Y.; Zhan, Q.; Wang, C.; Wang, S.-B.; Xie, S.-Q. Genome-wide association studies for five forage quality-related traits in sorghum (Sorghum bicolor L.). Front. Plant Sci. 2018, 9, 1146. [Google Scholar] [CrossRef]
- Boatright, J.; Negre, F.; Chen, X.; Kish, C.M.; Wood, B.; Peel, G.; Orlova, I.; Gang, D.; Rhodes, D.; Dudareva, N. Understanding in vivo benzenoid metabolism in petunia petal tissue. Plant Physiol. 2004, 135, 1993–2011. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Liu, X.; Sheng, Y.; Yang, H.; Xu, X.; Tao, Y.; Zhang, M. Optimization of different acid-catalyzed pretreatments on co-production of xylooligosaccharides and glucose from sorghum stalk. Polymers 2022, 14, 830. [Google Scholar] [CrossRef]
- Perrin, R.M.; DeRocher, A.E.; Bar-Peled, M.; Zeng, W.; Norambuena, L.; Orellana, A.; Raikhel, N.V.; Keegstra, K. Xyloglucan fucosyltransferase, an enzyme involved in plant cell wall biosynthesis. Science 1999, 284, 1976–1979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mace, E.S.; Hunt, C.H.; Jordan, D. Supermodels: Sorghum and maize provide mutual insight into the genetics of flowering time. Theor. Appl. Genet. 2013, 126, 1377–1395. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, D.; Hu, J.; Salas Fernandez, M.G. Genetic architecture of photosynthesis in Sorghum bicolor under non-stress and cold stress conditions. J. Exp. Bot. 2017, 68, 4545–4557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nijhawan, A.; Jain, M.; Tyagi, A.K.; Khurana, J.P. Genomic survey and gene expression analysis of the basic leucine zipper transcription factor family in rice. Plant Physiol. 2008, 146, 333. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Park, M.-R.; Kitazumi, A.; Herath, V.; Mohanty, B.; Yun, S.J.; de los Reyes, B.G. Cis-regulatory signatures of orthologous stress-associated bZIP transcription factors from rice, sorghum and Arabidopsis based on phylogenetic footprints. BMC Genom. 2012, 13, 497. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Mantilla Perez, M.B.; Hu, J.; Salas Fernandez, M.G. Genome-wide association study for nine plant architecture traits in Sorghum. Plant Genome 2016, 9. [Google Scholar] [CrossRef] [Green Version]
- Tao, Y.; Zhao, X.; Wang, X.; Hathorn, A.; Hunt, C.; Cruickshank, A.W.; van Oosterom, E.J.; Godwin, I.D.; Mace, E.S.; Jordan, D.R. Large-scale GWAS in sorghum reveals common genetic control of grain size among cereals. Plant Biotechnol. J. 2020, 18, 1093–1105. [Google Scholar] [CrossRef] [Green Version]
- Kevany, B.M.; Tieman, D.M.; Taylor, M.G.; Cin, V.D.; Klee, H.J. Ethylene receptor degradation controls the timing of ripening in tomato fruit. Plant J. 2007, 51, 458–467. [Google Scholar] [CrossRef]
- Klee, H.J. Control of ethylene-mediated processes in tomato at the level of receptors. J. Exp. Bot. 2002, 53, 2057–2063. [Google Scholar] [CrossRef] [Green Version]
- Klee, H.; Tieman, D. The tomato ethylene receptor gene family: Form and function. Physiol. Plant. 2002, 115, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.R.; Ramirez, M.V.; Miller, N.D.; Vallabhaneni, P.; Ray, W.K.; Helm, R.F.; Winkel, B.S.; Muday, G.K. Auxin and ethylene induce flavonol accumulation through distinct transcriptional networks. Plant Physiol. 2011, 156, 144–164. [Google Scholar] [CrossRef] [Green Version]
- Tovar-Méndez, A.; Miernyk, J.A.; Randall, D.D. Regulation of pyruvate dehydrogenase complex activity in plant cells. Eur. J. Biochem. 2003, 270, 1043–1049. [Google Scholar] [CrossRef]
- DeBrosse, S.D.; Kerr, D.S. Pyruvate Dehydrogenase Complex Deficiency; Elsevier: Amsterdam, The Netherlands, 2016; pp. 93–101. [Google Scholar]
- Noel, J.P.; Austin, M.B.; Bomati, E.K. Structure–function relationships in plant phenylpropanoid biosynthesis. Curr. Opin. Plant Biol. 2005, 8, 249–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenton, Z.W.; Cooper, E.A.; Myers, M.T.; Boyles, R.E.; Shakoor, N.; Zielinski, K.J.; Rauh, B.L.; Bridges, W.C.; Morris, G.P.; Kresovich, S. A genomic resource for the development, improvement, and exploitation of sorghum for bioenergy. Genetics 2016, 204, 21–33. [Google Scholar] [CrossRef] [Green Version]
- Niu, H.; Ping, J.; Wang, Y.; Lv, X.; Li, H.; Zhang, F.; Chu, J.; Han, Y. Population genomic and genome-wide association analysis of lignin content in a global collection of 206 forage sorghum accessions. Mol. Breed. 2020, 40, 73. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lines | Origin | Types | Accession Numbers | HD 1 (days) | PH 2 (cm) | SC 3 (brix°) | DY 4 (ton/ha) | TPC 5 (mg/100 g) |
|---|---|---|---|---|---|---|---|---|
| Gangwonsamcheok-2001-40 | Republic of Korea | Accession | IT218409 * | 70.0 | 311.0 | 18.5 | 9.5 | 7.31 |
| High-land-sweet | Republic of Korea | Accession | IR139445 * | 100.0 | 307.0 | 14.6 | 10.0 | 11.54 |
| DINE-A-MITE | Republic of Korea | Accession | IR100992 * | 97.0 | 362.0 | 9.3 | 17.4 | 5.87 |
| Pioneer931 | Republic of Korea | Accession | IT033846* | 115.0 | 348.0 | 16.7 | 15.2 | 8.04 |
| Kingsorgo | Republic of Korea | Accession | IT033841 * | 96.0 | 248.0 | 12.5 | 9.5 | 7.80 |
| IS645 | United State | Accession | IT124065 * | 90.0 | 330.0 | 18.1 | 11.9 | 6.67 |
| Ikumba | Kenya | Accession | IT262644 * | 98.0 | 331.0 | 15.8 | 17.4 | 3.36 |
| Olusi | Kenya | Accession | IT262629 * | 97.0 | 330.0 | 17.2 | 14.2 | 4.84 |
| Sorghum-medovoe | Russia | Accession | IT199372 * | 74.0 | 234.0 | 13.3 | 8.8 | 6.59 |
| IS2868 | South Africa | Accession | IT124094 * | 89.0 | 273.0 | 8.0 | 13.5 | 8.29 |
| IS5718 | India | Accession | IT124108 * | 90.0 | 253.0 | 5.0 | 5.9 | 8.13 |
| Andiwo-ma-rabour | Kenya | Accession | IT262529 * | 80.0 | 331.0 | 15.5 | 24.9 | 5.45 |
| Sabina | Kenya | Accession | IT262628 * | 90.0 | 340.0 | 15.2 | 14.2 | 7.09 |
| IS14131 | Portugal | Accession | IT143764 * | 91.0 | 333.0 | 11.2 | 9.5 | 3.77 |
| IS1211 | China | Accession | IT143846 * | 77.0 | 295.0 | 7.0 | 7.1 | 6.64 |
| KLSo79168 | Republic of Korea | Accession | IT028417 * | 75.0 | 347.0 | 8.5 | 10.3 | 7.03 |
| KLSo79125 | Republic of Korea | Accession | IT028385 * | 90.0 | 338.0 | 15.7 | 10.7 | 10.83 |
| KLSo79075 | Republic of Korea | Accession | IT028358 * | 71.0 | 275.0 | 12.2 | 6.6 | 6.02 |
| JM4621 | South Africa | Accession | IS27887 ** | 110.0 | 307.0 | 17.2 | 23.7 | 4.01 |
| JM 4682 | South Africa | Accession | IS27912 ** | 85.0 | 307.0 | 17.2 | 16.6 | 4.33 |
| Muansusu | Republic of Korea | Cultivar | IT028258 * | 69.0 | 284.0 | 9.8 | 13.5 | 8.69 |
| Nulsusu | Republic of Korea | Cultivar | IT185794 * | 102.0 | 354.0 | 11.0 | 11.1 | 10.50 |
| Chalsusu1 | Republic of Korea | Cultivar | IT191187 * | 99.0 | 343.0 | 10.2 | 11.9 | 7.72 |
| Shikyoung | Republic of Korea | Cultivar | IT105551 * | 100.0 | 341.0 | 11.5 | 9.5 | 6.27 |
| Jangmok | Republic of Korea | Cultivar | IT103274 * | 100.0 | 323.0 | 11.3 | 8.9 | 8.13 |
| Hansan | Republic of Korea | Cultivar | IT101381 * | 95.0 | 338.0 | 12.6 | 10.3 | 4.57 |
| Moktak | Republic of Korea | Cultivar | IT124114 * | 95.0 | 302.0 | 13.0 | 17.8 | 6.34 |
| Banwoldang | Republic of Korea | Cultivar | IT124115 * | 75.0 | 301.0 | 9.0 | 14.2 | 7.96 |
| Chalsusu2 | Republic of Korea | Cultivar | IT028260 * | 67.0 | 308.0 | 13.1 | 14.2 | 7.69 |
| Bitjaru | Republic of Korea | Cultivar | IT104110 * | 95.0 | 348.0 | 9.3 | 11.1 | 8.70 |
| Mesusu | Republic of Korea | Cultivar | IT028269 * | 98.0 | 326.0 | 12.7 | 11.9 | 3.75 |
| SOG102 | Republic of Korea | Accession | IS30507 ** | 85.0 | 229.0 | 18.8 | 9.5 | 9.11 |
| SOG103 | Republic of Korea | Accession | IS30508 ** | 82.0 | 233.0 | 16.0 | 13.5 | 4.47 |
| SOG129 | Republic of Korea | Accession | IS30533 ** | 80.0 | 252.0 | 18.1 | 15.9 | 3.46 |
| SOG132 | Republic of Korea | Accession | IS30536 ** | 75.0 | 320.0 | 18.8 | 19.0 | 5.27 |
| SOG159 | Republic of Korea | Accession | IS30562 ** | 92.0 | 260.0 | 15.0 | 14.2 | 6.51 |
| HDW501 | Indonesia | Accession | IS20956 ** | 100.0 | 265.0 | 12.9 | 11.9 | 3.27 |
| Banwoldang-1 | Banwoldang | Mutant | Gamma-ray 200 Gy *** | 100.0 | 329.0 | 15.3 | 11.1 | 6.76 |
| Banwoldang-2 | Banwoldang | Mutant | Gamma-ray 400 Gy | 62.0 | 137.0 | 12.1 | 9.5 | 8.22 |
| Banwoldang-3 | Banwoldang | Mutant | Gamma-ray 400 Gy | 62.0 | 126.0 | 16.3 | 9.5 | 10.64 |
| Banwoldang-4 | Banwoldang | Mutant | Gamma-ray 400 Gy | 100.0 | 175.0 | 12.5 | 11.9 | 10.95 |
| Banwoldang-5 | Banwoldang | Mutant | Gamma-ray 400 Gy | 110.0 | 135.0 | 13.1 | 9.5 | 5.24 |
| Banwoldang-6 | Banwoldang | Mutant | Gamma-ray 400 Gy | 110.0 | 155.0 | 15.2 | 11.9 | 9.81 |
| Banwoldang-7 | Banwoldang | Mutant | Gamma-ray 400 Gy | 58.0 | 125.0 | 13.0 | 9.5 | 8.19 |
| Banwoldang-8 | Banwoldang | Mutant | Gamma-ray 400 Gy | 62.0 | 215.0 | 17.2 | 14.2 | 3.86 |
| Banwoldang-9 | Banwoldang | Mutant | Gamma-ray 400 Gy | 59.0 | 112.0 | 15.3 | 11.9 | 7.70 |
| Banwoldang-10 | Banwoldang | Mutant | Gamma-ray 400 Gy | 110.0 | 369.0 | 14.5 | 11.9 | 4.00 |
| Banwoldang-11 | Banwoldang | Mutant | Gamma-ray 400 Gy | 110.0 | 374.0 | 14.2 | 11.9 | 2.59 |
| Banwoldang-12 | Banwoldang | Mutant | Gamma-ray 400 Gy | 70.0 | 252.0 | 14.8 | 16.6 | 6.12 |
| Banwoldang-13 | Banwoldang | Mutant | Gamma-ray 400 Gy | 80.0 | 376.0 | 10.8 | 16.6 | 7.39 |
| Banwoldang-14 | Banwoldang | Mutant | Gamma-ray 400 Gy | 80.0 | 365.0 | 12.5 | 16.6 | 7.06 |
| Banwoldang-15 | Banwoldang | Mutant | Gamma-ray 400 Gy | 100.0 | 342.0 | 11.9 | 16.6 | 3.89 |
| Banwoldang-16 | Banwoldang | Mutant | Gamma-ray 400 Gy | 100.0 | 412.0 | 12.4 | 23.7 | 5.09 |
| Banwoldang-17 | Banwoldang | Mutant | Gamma-ray 400 Gy | 100.0 | 402.0 | 11.6 | 23.7 | 5.55 |
| Banwoldang-18 | Banwoldang | Mutant | Gamma-ray 400 Gy | 102.0 | 242.0 | 13.9 | 8.3 | 5.34 |
| Dansusu2-1 | SOG103 | Mutant | Proton beam 300 Gy | 100.0 | 294.0 | 16.8 | 19.0 | 7.78 |
| Dansusu2-2 | SOG103 | Mutant | Proton beam 300 Gy | 100.0 | 308.0 | 15.8 | 16.6 | 9.21 |
| Dansusu2-3 | SOG103 | Mutant | Proton beam 300 Gy | 79.0 | 282.0 | 12.6 | 9.5 | 11.05 |
| Dansusu2-4 | SOG103 | Mutant | Proton beam 300 Gy | 79.0 | 290.0 | 13.4 | 9.5 | 4.97 |
| Dansusu2-5 | SOG103 | Mutant | Proton beam 300 Gy | 110.0 | 268.0 | 14.2 | 9.0 | 5.02 |
| Dansusu2-6 | SOG103 | Mutant | Proton beam 300 Gy | 100.0 | 308.0 | 16.2 | 10.7 | 6.14 |
| Dansusu2-7 | SOG103 | Mutant | Proton beam 300 Gy | 100.0 | 310.0 | 16.8 | 14.2 | 4.10 |
| Dansusu2-8 | SOG103 | Mutant | Proton beam 300 Gy | 100.0 | 89.0 | 5.0 | 2.4 | 4.04 |
| Dansusu2-9 | SOG103 | Mutant | Proton beam 300 Gy | 90.0 | 138.0 | 13.2 | 7.1 | 2.85 |
| Dansusu2-10 | SOG103 | Mutant | Gamma-ray 200 Gy | 105.0 | 253.0 | 16.5 | 9.5 | 5.87 |
| Dansusu2-11 | SOG103 | Mutant | Gamma-ray 200 Gy | 105.0 | 308.0 | 16.1 | 16.6 | 8.83 |
| Dansusu2-12 | SOG103 | Mutant | Gamma-ray 150 Gy | 105.0 | 282.0 | 14.8 | 11.9 | 3.03 |
| KLSo79125-1 | KLSo79125 | Mutant | Gamma-ray 400 Gy | 69.0 | 270.0 | 12.3 | 14.2 | 8.63 |
| KLSo79125-2 | KLSo79125 | Mutant | Gamma-ray 400 Gy | 69.0 | 214.0 | 16.3 | 11.9 | 8.87 |
| KLSo79125-3 | KLSO79125 | Mutant | Gamma-ray 200 Gy | 77.0 | 270.0 | 13.4 | 11.9 | 4.48 |
| KLSo79125-4 | KLSO79125 | Mutant | Gamma-ray 200 Gy | 100.0 | 352.0 | 13.3 | 14.2 | 3.94 |
| KLSo79125-5 | KLSO79125 | Mutant | Gamma-ray 200 Gy | 110.0 | 350.0 | 13.4 | 16.6 | 5.84 |
| Pahat-1 | HDW501 | Mutant | Gamma-ray 200 Gy | 90.0 | 240.0 | 17.8 | 14.2 | 3.21 |
| Pahat-2 | HDW501 | Mutant | Gamma-ray 200 Gy | 90.0 | 239.0 | 18.6 | 9.5 | 6.01 |
| Pahat-3 | HDW501 | Mutant | Gamma-ray 200 Gy | 69.0 | 100.0 | 10.7 | 7.1 | 4.23 |
| Pahat-4 | HDW501 | Mutant | Gamma-ray 200 Gy | 68.0 | 108.0 | 9.5 | 9.5 | 1.92 |
| Pahat-5 | HDW501 | Mutant | Gamma-ray 200 Gy | 110.0 | 95.0 | 10.1 | 7.6 | 5.34 |
| Pahat-6 | HDW501 | Mutant | Gamma-ray 200 Gy | 71.0 | 132.0 | 12.1 | 9.5 | 2.21 |
| IS5718-1 | IS5718 | Mutant | Gamma-ray 200 Gy | 62.0 | 220.0 | 14.2 | 8.1 | 9.00 |
| IS5718-2 | IS5718 | Mutant | Gamma-ray 200 Gy | 61.0 | 192.0 | 15.0 | 8.3 | 11.11 |
| IS5718-3 | IS5718 | Mutant | Gamma-ray 100 Gy | 61.0 | 240.0 | 12.6 | 8.3 | 6.55 |
| IS5718-4 | IS5718 | Mutant | Gamma-ray 100 Gy | 62.0 | 242.0 | 13.5 | 8.3 | 6.11 |
| IS645-1 | IS645 | Mutant | Gamma-ray 200 Gy | 99.0 | 373.0 | 14.5 | 21.3 | 9.08 |
| IS645-2 | IS645 | Mutant | Gamma-ray 200 Gy | 75.0 | 371.0 | 11.0 | 11.9 | 13.10 |
| IS645-3 | IS645 | Mutant | Gamma-ray 200 Gy | 61.0 | 348.0 | 12.8 | 26.1 | 3.95 |
| DINE-A-MITE-1 | DINE-A-MITE | Mutant | Gamma-ray 100 Gy | 102.0 | 465.0 | 6.0 | 23.7 | 4.22 |
| DINE-A-MITE-2 | DINE-A-MITE | Mutant | Gamma-ray 100 Gy | 77.0 | 319.0 | 10.5 | 11.9 | 9.16 |
| DINE-A-MITE-3 | DINE-A-MITE | Mutant | Gamma-ray 100 Gy | 105.0 | 410.0 | 8.5 | 16.6 | 5.99 |
| Moktak-1 | Moktak | Mutant | Gamma-ray 100 Gy | 68.0 | 289.0 | 15.2 | 14.2 | 7.17 |
| Moktak-2 | Moktak | Mutant | Gamma-ray 100 Gy | 100.0 | 390.0 | 15.3 | 26.1 | 6.96 |
| Chalsusu1-1 | Chalsusu1 | Mutant | Gamma-ray 200 Gy | 110.0 | 408.0 | 16.4 | 16.6 | 5.57 |
| High-land-sweet-1 | High-land-sweet | Mutant | Gamma-ray 200 Gy | 71.0 | 270.0 | 16.3 | 11.9 | 4.80 |
| IS2868-1 | IS2868 | Mutant | Gamma-ray 100 Gy | 90.0 | 345.0 | 16.9 | 21.3 | 9.28 |
| Mesusu-1 | Mesusu | Mutant | Gamma-ray 100 Gy | 105.0 | 375.0 | 12.8 | 14.2 | 2.85 |
| IS2864-1 | South Africa | Mutant | Gamma-ray 100 Gy | 69.0 | 108.0 | 8.2 | 7.1 | 2.81 |
| IS2864-2 | South Africa | Mutant | Gamma-ray 100 Gy | 90.0 | 240.0 | 12.6 | 8.3 | 2.25 |
| Trait 1 | Min 2 | Max 3 | Mean | Skew 4 | Kurt 5 | CV 6 |
|---|---|---|---|---|---|---|
| HD | 58.0 | 115.0 | 87.8 | −0.29 | −1.19 | 0.18 |
| PH | 89.0 | 465.0 | 282.0 | −0.66 | −0.06 | 0.30 |
| SC | 5.0 | 18.8 | 13.4 | −0.53 | 0.08 | 0.23 |
| DY | 2.4 | 26.1 | 13.0 | 0.90 | 0.65 | 0.37 |
| Type | Min 1 | Max 2 | Mean | Skew 3 | Kurt 4 | CV 5 |
|---|---|---|---|---|---|---|
| Luteolinidin diglucoside | 0.09 | 0.46 | 0.23 | 1.08 | 2.13 | 0.29 |
| Luteolin glucoside | 0.00 | 1.24 | 0.30 | 1.33 | 2.02 | 0.90 |
| Apigeninidin glucoside | 0.02 | 1.18 | 0.40 | 1.01 | 2.29 | 0.49 |
| Luteolinidin | 0.06 | 5.20 | 1.71 | 0.76 | 0.39 | 0.67 |
| Apigeninidin | 0.00 | 0.75 | 0.32 | 0.25 | −0.13 | 0.52 |
| 5-O-Me-luteolinidin | 0.01 | 0.43 | 0.16 | 0.16 | −0.81 | 0.50 |
| Total phenolic content (TPC) | 1.92 | 13.10 | 6.37 | 0.36 | −0.44 | 0.38 |
| Total | Average/Plant | |
|---|---|---|
| Raw data | ||
| Reads | 684,426,636 | 7,129,444 |
| Bases (bp) | 103,348,422,036 | 1,076,546,063 |
| After trimming | ||
| Reads | 620,196,808 | 6,460,383 |
| Bases (bp) | 64,640,661,227 | 673,340,221 |
| Mapped reads on reference genome 1 | ||
| Reads | 599,168,188 | 6,241,335 |
| Bases (bp) | 4,968,266,855 | 51,752,780 |
| Reference genome coverage (%) | 7.0954% | |
| Chromosome | Length (bp) | No. of SNPs | Kb/SNP | SNPs/Mb |
|---|---|---|---|---|
| Chromosome 1 | 81,498,373 | 25,265 | 3.23 | 310.0 |
| Chromosome 2 | 89,798,109 | 23,970 | 3.75 | 266.9 |
| Chromosome 3 | 75,771,322 | 20,189 | 3.75 | 266.4 |
| Chromosome 4 | 66,264,056 | 19,532 | 3.39 | 294.8 |
| Chromosome 5 | 74,474,820 | 20,707 | 3.60 | 278.0 |
| Chromosome 6 | 69,324,445 | 18,302 | 3.79 | 264.0 |
| Chromosome 7 | 70,271,347 | 13,830 | 5.08 | 196.8 |
| Chromosome 8 | 70,703,592 | 17,045 | 4.15 | 241.1 |
| Chromosome 9 | 60,147,662 | 15,641 | 3.85 | 260.0 |
| Chromosome 10 | 61,107,423 | 16,986 | 3.60 | 278.0 |
| Scaffolds | 10,018,713 | 573 | 17.48 | 57.2 |
| Total | 729,379,862 | 192,040 | ||
| Mean | 5.06 | 246.7 |
| SNP | Trait 1 | Chr. | Position (bp) | Effect | −log10(p) | MAF 2 | Allele | Method 3 | Candidate Gene 4 | Description |
|---|---|---|---|---|---|---|---|---|---|---|
| Sb02_6876523 | HD | 2 | 6,876,523 | −8.14 | 4.15 | 0.42 | G/A | 1, 2 | SbRio.02G064100 | benzyl alcohol O-benzoyltransferase |
| Sb02_6876524 | HD | 2 | 6,876,524 | 8.14 | 4.15 | 0.42 | G/T | 1, 2 | SbRio.02G064100 | benzyl alcohol O-benzoyltransferase |
| Sb06_8705823 | HD | 6 | 8,705,823 | −9.74 | 4.16 | 0.47 | T/C | 1, 2 | SbRio.06G036000 | endo−1,4-beta-xylanase 4-like isoform X2 |
| Sb10_7471984 | HD | 10 | 7,471,984 | −12.67–12.19 | 4.20–4.93 | 0.16 | A/G | 1, 2, 3, 4 | SbRio.10G099600 | galactoside 2-alpha-L-fucosyltransferase-like isoform X2 |
| Sb07_53523852 | PH | 7 | 53,523,852 | −82.68–54.15 | 4.12–4.18 | 0.11 | T/C | 3, 4 | SbRio.07G123800 | nudix hydrolase 15, mitochondrial |
| Sb08_63291752 | PH | 8 | 63,291,752 | −49.51–38.37 | 7.80–8.08 | 0.19 | G/A | 1, 2 | SbRio.08G141500 | hypothetical protein BDA96_08G141500 |
| Sb09_50399847 | PH | 9 | 50,399,847 | −127.68–117.14 | 5.77–11.70 | 0.18 | G/A | 3, 4 | SbRio.09G149200 | hypothetical protein BDA96_09G149200 |
| Sb04_2143594 | DY | 4 | 2,143,594 | 1.32–3.97 | 4.40–6.82 | 0.15 | G/C | 1, 2, 4 | SbRio.04G031300 | beta-amylase 8 isoform X2 |
| Sb06_62687750 | DY | 6 | 62,687,750 | 1.79–4.02 | 4.20–6.08 | 0.17 | G/A | 2, 4 | SbRio.06G211400 | transcription factor MafB |
| SNP | Trait 1 | Chr | Position (bp) | Effect | −log10(p) | MAF 2 | Allele | Method 3 | Candidate gene 4 | Description |
|---|---|---|---|---|---|---|---|---|---|---|
| Sb04_1305322 | 1 | 4 | 1,305,322 | −1.68–1.05 | 4.03–6.69 | 0.33 | T/C | 2, 3 | SbRio.04G019000 | choline/ethanolaminephosphotransferase 1 |
| Sb04_64461978 | 1 | 4 | 64,461,978 | −0.88–1.16 | 7.28–7.67 | 0.30 | C/A | 1, 2 | SbRio.04G361300 | glycosyltransferase family protein 2 |
| Sb04_56584914 | 1 | 4 | 56,584,914 | −3.49 | 4.06 | 0.08 | C/G | 4 | SbRio.04G259800 | pyruvate dehydrogenase E1 component subunit alpha-1, mitochondrial |
| 2 | 4 | −0.11 | 4.40 | 0.08 | C/G | 4 | SbRio.04G259800 | pyruvate dehydrogenase E1 component subunit alpha-1, mitochondrial | ||
| Sb02_81796960 | 1 | 2 | 81,796,960 | 1.85 | 4.10 | 0.28 | T/C | 4 | SbRio.02G343600 | ethylene receptor 4 |
| 5 | 0.80 | 4.64 | 0.28 | T/C | 3 | SbRio.02G343600 | ethylene receptor 4 | |||
| Sb02_81797062 | 1 | 2 | 81,797,062 | −2.29–1.43 | 6.55–14.04 | 0.24 | A/G | 1, 2, 3 | SbRio.02G343600 | ethylene receptor 4 |
| 5 | −0.87 | 4.58 | 0.24 | A/G | 3 | SbRio.02G343600 | ethylene receptor 4 | |||
| Sb02_81797139 | 1 | 2 | 81,797,139 | 2.18 | 4.56 | 0.26 | G/C | 4 | SbRio.02G343600 | ethylene receptor 4 |
| 5 | 2 | 0.91 | 4.23–4.98 | 0.26 | G/C | 3, 4 | SbRio.02G343600 | ethylene receptor 4 | ||
| Sb02_79905727 | 2 | 2 | 79,905,727 | 0.03–0.04 | 4.52–5.39 | 0.17 | A/T | 1, 2, 3 | SbRio.02G316600 | NEP1-interacting protein-like 1 |
| Sb04_59016630 | 2 | 4 | 59,016,630 | −0.14–0.06 | 4.04–22.41 | 0.12 | G/T | 1, 2, 4 | SbRio.04G289300 | glutamic acid-rich protein-like isoform X1 |
| Sb06_1797783 | 2 | 6 | 1,797,783 | 0.05–0.06 | 4.10–4.39 | 0.16 | T/G | 1, 3, 4 | SbRio.06G011700 | fe-S cluster assembly factor HCF101, chloroplastic |
| Sb06_59065337 | 2 | 6 | 59,065,337 | −0.03 | 4.56–4.76 | 0.35 | C/T | 1, 3 | SbRio.06G167800 | wall-associated receptor kinase 4 |
| Sb06_59065351 | 2 | 6 | 59,065,351 | 0.03 | 4.81–4.85 | 0.36 | T/G | 1, 3 | SbRio.06G167800 | wall-associated receptor kinase 4 |
| Sb06_59065380 | 2 | 6 | 59,065,380 | 0.03 | 4.30–4.65 | 0.34 | G/C | 1, 3 | SbRio.06G167800 | wall-associated receptor kinase 4 |
| Sb06_59065407 | 2 | 6 | 59,065,407 | 0.03 | 4.07–4.42 | 0.34 | T/A | 1, 3 | SbRio.06G167800 | wall-associated receptor kinase 4 |
| Sb01_14431763 | 3 | 1 | 14,431,763 | 0.16 | 4.01 | 0.38 | G/A | 1, 2 | SbRio.01G175100 | ARM repeat superfamily protein | calcium-transporting ATPase 3, plasma membrane-type |
| Sb03_4939603 | 3 | 3 | 4,939,603 | 0.17 | 4.18–4.27 | 0.30 | A/G | 1, 2, 3 | SbRio.03G056200 | hypothetical protein BDA96_03G056200 |
| Sb05_9038134 | 3 | 5 | 9,038,134 | 0.18 | 4.03 | 0.20 | G/A | 1, 2 | SbRio.05G076500 | probable kinase CHARK |
| Sb05_9038126 | 3 | 5 | 9,038,126 | −0.20 | 4.12 | 0.18 | C/G | 1, 2 | SbRio.05G076500 | probable kinase CHARK |
| Sb01_1229036 | 4 | 1 | 1,229,036 | 0.09 | 4.08 | 0.18 | G/C | 1, 3 | SbRio.01G011000 | U2 small nuclear ribonucleoprotein A’ |
| Sb01_1229046 | 4 | 1 | 1,229,046 | −0.09 | 4.08 | 0.18 | A/C | 1, 3 | SbRio.01G011000 | U2 small nuclear ribonucleoprotein A’ |
| Sb06_55785073 | 4 | 6 | 55,785,073 | 0.16–0.25 | 5.52–11.40 | 0.09 | G/C | 1, 2, 3, 4 | SbRio.06G125400 | disease resistance protein Pik-1 |
| Sb03_68395304 | 4 | 3 | 68,395,304 | 0.10 | 4.58 | 0.23 | C/T | 1, 3 | SbRio.03G375900 | ruvB-like protein 1 |
| Sb03_68358847 | 4 | 3 | 68,358,847 | 0.09 | 4.46 | 0.27 | G/T | 1, 3 | SbRio.03G375100 | protein GPR107 |
| Sb03_68358815 | 4 | 3 | 68,358,815 | 0.09 | 4.72 | 0.26 | C/T | 1, 3 | SbRio.03G375100 | protein GPR107 |
| Sb03_68358771 | 4 | 3 | 68,358,771 | −0.09 | 4.41 | 0.27 | T/C | 1, 3 | SbRio.03G375100 | protein GPR107 |
| Sb10_4609482 | 4 | 10 | 4,609,482 | −0.16–0.27 | 4.65–7.73 | 0.08 | C/G | 2, 3, 4 | SbRio.10G064200 | proteasome subunit alpha type-4-2 |
| Sb06_68347889 | 6 | 6 | 68,347,889 | −0.09–0.07 | 4.51–7.23 | 0.17 | A/G | 1, 2, 3, 4 | SbRio.06G295300 | multiple RNA-binding domain-containing protein 1 |
| Sb08_65531987 | 6 | 8 | 65,531,987 | −0.07–0.05 | 4.23–5.94 | 0.31 | A/G | 1, 2, 4 | SbRio.08G160200 | Os04g0380500 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.-J.; Yang, B.; Kim, W.J.; Kim, J.; Kwon, S.-J.; Kim, J.H.; Ahn, J.-W.; Kim, S.H.; Rha, E.-S.; Ha, B.-K.; et al. Genome-Wide Association Study (GWAS) of the Agronomic Traits and Phenolic Content in Sorghum (Sorghum bicolor L.) Genotypes. Agronomy 2023, 13, 1449. https://doi.org/10.3390/agronomy13061449
Lee Y-J, Yang B, Kim WJ, Kim J, Kwon S-J, Kim JH, Ahn J-W, Kim SH, Rha E-S, Ha B-K, et al. Genome-Wide Association Study (GWAS) of the Agronomic Traits and Phenolic Content in Sorghum (Sorghum bicolor L.) Genotypes. Agronomy. 2023; 13(6):1449. https://doi.org/10.3390/agronomy13061449
Chicago/Turabian StyleLee, Ye-Jin, Baul Yang, Woon Ji Kim, Juyoung Kim, Soon-Jae Kwon, Jae Hoon Kim, Joon-Woo Ahn, Sang Hoon Kim, Eui-Shik Rha, Bo-Keun Ha, and et al. 2023. "Genome-Wide Association Study (GWAS) of the Agronomic Traits and Phenolic Content in Sorghum (Sorghum bicolor L.) Genotypes" Agronomy 13, no. 6: 1449. https://doi.org/10.3390/agronomy13061449
APA StyleLee, Y. -J., Yang, B., Kim, W. J., Kim, J., Kwon, S. -J., Kim, J. H., Ahn, J. -W., Kim, S. H., Rha, E. -S., Ha, B. -K., Bae, C. -H., & Ryu, J. (2023). Genome-Wide Association Study (GWAS) of the Agronomic Traits and Phenolic Content in Sorghum (Sorghum bicolor L.) Genotypes. Agronomy, 13(6), 1449. https://doi.org/10.3390/agronomy13061449