
Integrative Effects of Zinc Nanoparticle and PGRs to Mitigate Salt Stress in Maize


Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Conditions, Experimental Design, and Plant Materials
2.2. Nutrient Solution and Treatments
2.3. Evaluation of the Seedling’s Growth Parameters
2.4. Measurement of the Root Parameters
2.5. Measurement of Photosynthetic Traits and Maximum Quantum Yield of PSII
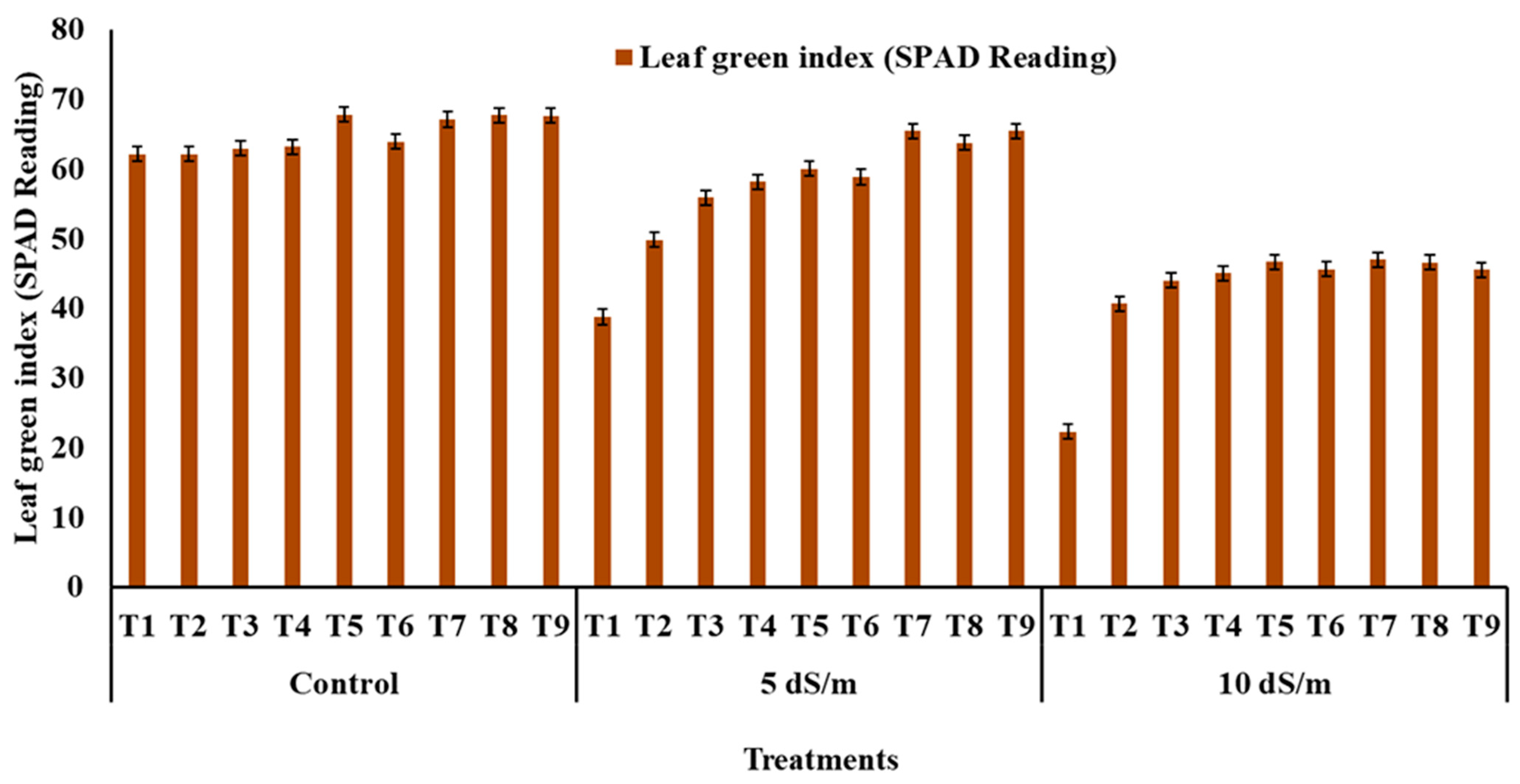
2.6. Determination of the Photosynthesis Pigments and SPAD
2.7. Na+, K+ and Zn2+ Concentration
2.8. Proline and Phenolic Content
2.9. Statistical Analysis
3. Results and Discussion
3.1. Root Growth and Development
3.2. Seedling Growth Performance
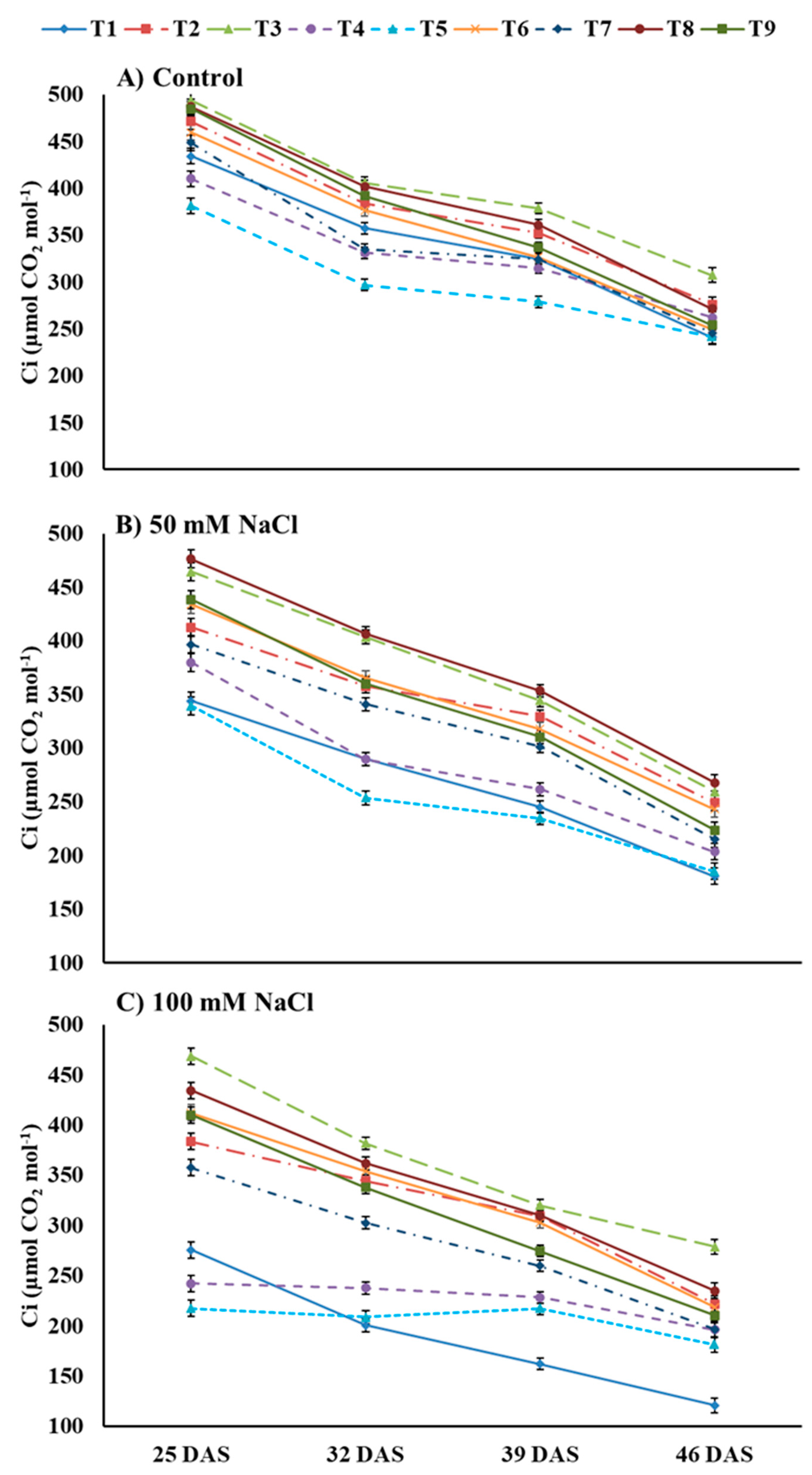
3.3. Photosynthesis and Contributing Traits
3.4. Photosynthetic Pigments, Phenolic, and Proline Content
3.5. Na, K, and Zn Uptake
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Beltran-Peña, A.; Rosa, L.; D’Odorico, P. Global food self-sufficiency in the 21st century under sustainable intensification of agriculture. Environ. Res. Lett. 2020, 15, 095004. [Google Scholar] [CrossRef]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [Green Version]
- Hunter, M.C.; Smith, R.G.; Schipanski, M.E.; Atwood, L.W.; Mortensen, D.A. Agriculture in 2050: Recalibrating targets for sustainable intensification. BioScience 2017, 67, 386–391. [Google Scholar] [CrossRef] [Green Version]
- Hemathilake, D.M.K.S.; Gunathilake, D.M.C.C. Agricultural productivity and food supply to meet increased demands. In Future Foods; Academic Press: Cambridge, MA, USA, 2022; pp. 539–553. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Selim, S.; Alhammad, B.A.; Alharbi, B.M.; Juliatti, F.C. Will novel coronavirus (COVID-19) pandemic impact agriculture, food security and animal sectors? Biosci. J. 2020, 36, 1315–1326. [Google Scholar] [CrossRef]
- Cowan, N.; Ferrier, L.; Spears, B.; Drewer, J.; Reay, D.; Skiba, U. CEA systems: The means to achieve future food security and environmental sustainability? Front. Sustain. Food Syst. 2022, 6, 891256. [Google Scholar] [CrossRef]
- Ojo, M.O.; Zahid, A. Deep Learning in Controlled Environment Agriculture: A Review of Recent Advancements, Challenges and Prospects. Sensors 2022, 22, 7965. [Google Scholar] [CrossRef]
- Ragaveena, S.; Shirly Edward, A.; Surendran, U. Smart controlled environment agriculture methods: A holistic review. Rev. Environ. Sci. Bio/Technol. 2021, 20, 887–913. [Google Scholar] [CrossRef]
- Abusin, S.A.; Mandikiana, B.W. Towards sustainable food production systems in Qatar: Assessment of the viability of aquaponics. Glob. Food Secur. 2020, 25, 100349. [Google Scholar] [CrossRef]
- Ahamed, M.S.; Sultan, M.; Shamshiri, R.R.; Rahman, M.M.; Aleem, M.; Balasundram, S.K. Present status and challenges of fodder production in controlled environments: A review. Smart Agric. Technol. 2022, 3, 100080. [Google Scholar] [CrossRef]
- Bione, M.A.A.; Soares, T.M.; Cova, A.M.W.; da Silva Paz, V.P.; Gheyi, H.R.; Rafael, M.R.S.; Modesto, F.J.N.; da Anunciação Santana, J.; das Neves, B.S.L. Hydroponic production of ‘Biquinho’pepper with brackish water. Agric. Water Manag. 2021, 245, 106607. [Google Scholar] [CrossRef]
- Costan, A.; Stamatakis, A.; Chrysargyris, A.; Petropoulos, S.A.; Tzortzakis, N. Interactive effects of salinity and silicon application on Solanum lycopersicum growth, physiology and shelf-life of fruit produced hydroponically. J. Sci. Food Agric. 2020, 100, 732–743. [Google Scholar] [CrossRef]
- Loudari, A.; Benadis, C.; Naciri, R.; Soulaimani, A.; Zeroual, Y.; Gharous, M.E.; Kalaji, H.M.; Oukarroum, A. Salt stress affects mineral nutrition in shoots and roots and chlorophyll a fluorescence of tomato plants grown in hydroponic culture. J. Plant Interact. 2020, 15, 398–405. [Google Scholar] [CrossRef]
- Tola, E.; Al-Gaadi, K.A.; Madugundu, R.; Patil, V.C.; Sygrimis, N. Impact of water salinity levels on the spectral behavior and yield of tomatoes in hydroponics. J. King Saud Univ.-Sci. 2023, 35, 102515. [Google Scholar] [CrossRef]
- Moncada, A.; Vetrano, F.; Miceli, A. Alleviation of salt stress by plant growth-promoting bacteria in hydroponic leaf lettuce. Agronomy 2020, 10, 1523. [Google Scholar] [CrossRef]
- Harizanova, A.; Koleva-Valkova, L. Effect of silicon on photosynthetic rate and the chlorophyll fluorescence parameters at hydroponically grown cucumber plants under salinity stress. J. Cent. Eur. Agric. 2019, 20, 953–960. [Google Scholar] [CrossRef]
- Kaloterakis, N.; van Delden, S.H.; Hartley, S.; De Deyn, G.B. Silicon application and plant growth promoting rhizobacteria consisting of six pure Bacillus species alleviate salinity stress in cucumber (Cucumis sativus L.). Sci. Hortic. 2021, 288, 110383. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Selwey, W.A.; Ibrahim, A.A.; Shady, M.; Alsadon, A.A. Foliar Applications of ZnO and SiO2 Nanoparticles Mitigate Water Deficit and Enhance Potato Yield and Quality Traits. Agronomy 2023, 13, 466. [Google Scholar] [CrossRef]
- Singh, R.P.; Handa, R.; Manchanda, G. Nanoparticles in sustainable agriculture: An emerging opportunity. J. Control. Release 2021, 329, 1234–1248. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Ahmad, A.; Battaglia, M.L.; Bilal, H.M.; Alhammad, B.A.; Khan, K. Zinc oxide nanoparticles: A unique saline stress mitigator with the potential to increase future crop production. S. Afr. J. Bot. 2023, 159, 208–218. [Google Scholar] [CrossRef]
- Alhammad, B.A.; Ahmad, A.; Seleiman, M.F.; Tola, E. Seed Priming with Nanoparticles and 24-Epibrassinolide Improved Seed Germination and Enzymatic Performance of Zea mays L. in Salt-Stressed Soil. Plants 2023, 12, 690. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Tola, E.; Alshahrani, T.S.; Seleiman, M.F. Enhancement of morphological and physiological performance of Zea Mays L. under saline stress using ZnO nanoparticles and 24-epibrassinolide seed priming. Agronomy 2023, 13, 771. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, R.; Chen, Z.; Cui, P.; Lu, H.; Yang, Y.; Zhang, H. The effect of zinc oxide nanoparticles for enhancing rice (Oryza sativa L.) yield and quality. Agriculture 2021, 11, 1247. [Google Scholar] [CrossRef]
- Adrees, M.; Khan, Z.S.; Hafeez, M.; Rizwan, M.; Hussain, K.; Asrar, M.; Alyemeni, M.N.; Wijaya, L.; Ali, S. Foliar exposure of zinc oxide nanoparticles improved the growth of wheat (Triticum aestivum L.) and decreased cadmium concentration in grains under simultaneous Cd and water deficient stress. Ecotoxicol. Environ. Saf. 2021, 208, 111627. [Google Scholar] [CrossRef] [PubMed]
- Faizan, M.; Bhat, J.A.; Chen, C.; Alyemeni, M.N.; Wijaya, L.; Ahmad, P.; Yu, F. Zinc oxide nanoparticles (ZnO-NPs) induce salt tolerance by improving the antioxidant system and photosynthetic machinery in tomato. Plant Physiol. Biochem. 2021, 161, 122–130. [Google Scholar] [CrossRef]
- Adil, M.; Bashir, S.; Bashir, S.; Aslam, Z.; Ahmad, N.; Younas, T.; Asghar, R.M.A.; Alkahtani, J.; Dwiningsih, Y.; Elshikh, M.S. Zinc oxide nanoparticles improved chlorophyll contents, physical parameters, and wheat yield under salt stress. Front. Plant Sci. 2022, 13, 932861. [Google Scholar] [CrossRef]
- Aqeel, U.; Aftab, T.; Khan, M.M.A.; Naeem, M.; Khan, M.N. A comprehensive review of impacts of diverse nanoparticles on growth, development and physiological adjustments in plants under changing environment. Chemosphere 2022, 291, 132672. [Google Scholar] [CrossRef]
- Tanveer, M.; Shahzad, B.; Sharma, A.; Biju, S.; Bhardwaj, R. 24-Epibrassinolide; an active brassinolide and its role in salt stress tolerance in plants: A review. Plant Physiol. Biochem. 2018, 130, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.M.; Anwar, R.; Malik, A.U.; Khan, A.S.; Ahmad, S.; Hussain, Z.; Hasan, M.U.; Nasir, M.; Chen, F. Plant growth and fruit quality response of strawberry is improved after exogenous application of 24-epibrassinolide. J. Plant Growth Regul. 2022, 41, 1786–1799. [Google Scholar] [CrossRef]
- Maia, C.F.; da Silva, B.R.S.; Batista, B.L.; Bajguz, A.; Lobato, A.K.D.S. 24-Epibrassinolide simultaneously stimulates photosynthetic machinery and biomass accumulation in tomato plants under lead stress: Essential contributions connected to the antioxidant system and anatomical structures. Agronomy 2022, 12, 1985. [Google Scholar] [CrossRef]
- Liu, Z.M.; Faizan, M.; Chen, C.; Zheng, L.H.; Yu, F.Y. The combined analysis of transcriptome and antioxidant enzymes revealed the mechanism of EBL and ZnO NPs enhancing Styrax tonkinensis seed abiotic stress resistance. Genes 2022, 13, 2170. [Google Scholar] [CrossRef]
- Hosseinpour, M.; Ebadi, A.; Habibi, H.; Nabizadeh, E.; Jahanbakhsh, S. Enhancing enzymatic and nonenzymatic response of Echinacea purpurea by exogenous 24-epibrassinolide under drought stress. Ind. Crops Prod. 2020, 146, 112045. [Google Scholar] [CrossRef]
- Faizan, M.; Bhat, J.A.; Noureldeen, A.; Ahmad, P.; Yu, F. Zinc oxide nanoparticles and 24-epibrassinolide alleviates Cu toxicity in tomato by regulating ROS scavenging, stomatal movement and photosynthesis. Ecotoxicol. Environ. Saf. 2021, 218, 112293. [Google Scholar] [CrossRef]
- Hafeez, M.B.; Zahra, N.; Zahra, K.; Raza, A.; Batool, A.; Shaukat, K.; Khan, S. Brassinosteroids: Molecular and physiological responses in plant growth and abiotic stresses. Plant Stress 2021, 2, 100029. [Google Scholar] [CrossRef]
- Tanveer, M. Role of 24-Epibrassinolide in inducing thermo-tolerance in plants. J. Plant Growth Regul. 2019, 38, 945–955. [Google Scholar] [CrossRef]
- Hu, S.; Ma, Y.; Xie, B.; Hou, Y.; Jia, Z.; Zhao, L.; Zheng, Y.; Jin, P. 24-Epibrassinolide improves chilling tolerance by regulating PpCBF5-mediated membrane lipid metabolism in peach fruit. Postharvest Biol. Technol. 2022, 186, 111844. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, B.; Tanveer, M.; Che, Z.; Rehman, A.; Cheema, S.A.; Sharma, A.; Song, H.; ur Rehman, S.; Zhaorong, D. Role of 24-epibrassinolide (EBL) in mediating heavy metal and pesticide induced oxidative stress in plants: A review. Ecotoxicol. Environ. Saf. 2018, 147, 935–944. [Google Scholar] [CrossRef]
- Faizan, M.; Bhat, J.A.; Hessini, K.; Yu, F.; Ahmad, P. Zinc oxide nanoparticles alleviates the adverse effects of cadmium stress on Oryza sativa via modulation of the photosynthesis and antioxidant defense system. Ecotoxicol. Environ. Saf. 2021, 220, 112401. [Google Scholar] [CrossRef]
- Dong, Y.; Wang, W.; Hu, G.; Chen, W.; Zhuge, Y.; Wang, Z.; He, M.R. Role of exogenous 24-epibrassinolide in enhancing the salt tolerance of wheat seedlings. J. Soil Sci. Plant Nutr. 2017, 17, 554–569. [Google Scholar] [CrossRef] [Green Version]
- Liaqat, S.; Umar, S.; Saffeullah, P.; Iqbal, N.; Siddiqi, T.O.; Khan, M.I.R. Protective Effect of 24-epibrassinolide on barley plants growing under combined stress of salinity and potassium deficiency. J. Plant Growth Regul. 2020, 39, 1543–1558. [Google Scholar] [CrossRef]
- Sousa, V.Q.; Messias, W.F.S.; Pereira, Y.C.; da Silva, B.R.S.; Lobato, E.M.S.G.; Alyemeni, M.N.; Ahmad, P.; Lobato, A.K.D.S. Pretreatment with 24-epibrassinolide synergistically protects root structures and chloroplastic pigments and upregulates antioxidant enzymes and biomass in Na+-stressed tomato plants. J. Plant Growth Regul. 2021, 41, 2869–2885. [Google Scholar] [CrossRef]
- Li, W.; Sun, J.; Zhang, X.; Ahmad, N.; Hou, L.; Zhao, C.; Pan, J.; Tian, R.; Wang, X.; Zhao, S. The mechanisms underlying salt resistance mediated by exogenous application of 24-epibrassinolide in peanut. Int. J. Mol. Sci. 2022, 23, 6376. [Google Scholar] [CrossRef]
- Basit, F.; Chen, M.; Ahmed, T.; Shahid, M.; Noman, M.; Liu, J.; An, J.; Hashem, A.; Fahad Al Arjani, A.B.; Alqarawi, A.A.; et al. Seed priming with brassinosteroids alleviates chromium stress in rice cultivars via improving ROS metabolism and antioxidant defense response at biochemical and molecular levels. Antioxidants 2021, 10, 1089. [Google Scholar] [CrossRef] [PubMed]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajput, R.D.; Patil, R.P. The comparative study on spectrophotometric analysis of chlorophyll and carotenoids pigments from non-leguminous fodder crops. Int. J. Innov. Sci. Eng. Technol. 2017, 4, 140–148. [Google Scholar]
- Wolf, B. The comprehensive system of leaf analysis and its use for diagnosing crop nutrient status. Commun. Soil Sci. Plant Anal. 1982, 3, 1035–1059. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.A.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Tawaha, K.; Alali, F.Q.; Gharaibeh, M.; Mohammad, M.; El-Elimat, T. Antioxidant activity and total phenolic content of selected Jordanian plant species. Food Chem. 2017, 104, 1372–1378. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Hao, S.; Wang, Y.; Yan, Y.; Liu, Y.; Wang, J.; Chen, S. A review on plant responses to salt stress and their mechanisms of salt resistance. Horticulturae 2021, 7, 132. [Google Scholar] [CrossRef]
- Yang, Z.; Li, J.L.; Liu, L.N.; Xie, Q.; Sui, N. Photosynthetic regulation under salt stress and salt-tolerance mechanism of sweet sorghum. Front. Plant Sci. 2020, 10, 1722. [Google Scholar] [CrossRef] [Green Version]
- Zahra, N.; Raza, Z.A.; Mahmood, S. Effect of salinity stress on various growth and physiological attributes of two contrasting maize genotypes. Braz. Arch. Biol. Technol. 2020, 63, 72. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, G.; Wang, Y.; Zhou, Z.; Meng, Y.; Chen, B. Effect of soil salinity on physiological characteristics of functional leaves of cotton plants. J. Plant Res. 2013, 126, 293–304. [Google Scholar] [CrossRef]
- West, G.; Inzé, D.; Beemster, G.T. Cell cycle modulation in the response of the primary root of Arabidopsis to salt stress. Plant Physiol. 2004, 135, 1050–1058. [Google Scholar] [CrossRef]
- Shahzad, R.; Harlina, P.W.; Ewas, M.; Zhenyuan, P.; Nie, X.; Gallego, P.P.; Ullah Khan, S.; Nishawy, E.; Khan, A.H.; Jia, H. Foliar applied 24-epibrassinolide alleviates salt stress in rice (Oryza sativa L.) by suppression of ABA levels and upregulation of secondary metabolites. J. Plant Interact. 2021, 16, 533–549. [Google Scholar] [CrossRef]
- Ji, Y.; Zhou, Y.; Ma, C.; Feng, Y.; Hao, Y.; Rui, Y.; Wu, W.; Gui, X.; Han, Y.; Wang, Y.; et al. Jointed toxicity of TiO2 NPs and Cd to rice seedlings: NPs alleviated Cd toxicity and Cd promoted NPs uptake. Plant Physiol. Biochem. 2017, 110, 82–93. [Google Scholar] [CrossRef]
- Faizan, M.; Hayat, S.; Pichtel, J. Effects of zinc oxide nanoparticles on crop plants: A perspective analysis. In Sustainable Agriculture Reviews 41; Springer: Cham, Switzerland, 2020; pp. 83–99. [Google Scholar] [CrossRef]
- Paramo, L.A.; Feregrino-Pérez, A.A.; Guevara, R.; Mendoza, S.; Esquivel, K. Nanoparticles in agroindustry: Applications, toxicity, challenges, and trends. Nanomaterials 2020, 10, 1654. [Google Scholar] [CrossRef] [PubMed]
- Chaiwanon, J.; Wang, Z.Y. Spatiotemporal brassinosteroid signaling and antagonism with auxin pattern stem cell dynamics in Arabidopsis roots. Curr. Biol. 2015, 25, 1031–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerona, M.E.B.; Deocampo, M.P.; Egdane, J.A.; Ismail, A.M.; Dionisio-Sese, M.L. Physiological responses of contrasting rice genotypes to salt stress at reproductive stage. Rice Sci. 2019, 26, 207–219. [Google Scholar] [CrossRef]
- Guo, Q.; Liu, L.; Barkla, B.J. Membrane lipid remodeling in response to salinity. Int. J. Mol. Sci. 2019, 20, 4264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, D.M.; Oliveira, M.M.; Saibo, N.J. Regulation of Na+ and K+ homeostasis in plants: Towards improved salt stress tolerance in crop plants. Genet. Mol. Biol. 2017, 40, 326–345. [Google Scholar] [CrossRef] [Green Version]
- Yun, P.; Xu, L.; Wang, S.S.; Shabala, L.; Shabala, S.; Zhang, W.Y. Piriformospora indica improves salinity stress tolerance in Zea mays L. plants by regulating Na+ and K+ loading in root and allocating K+ in shoot. Plant Growth Regul. 2018, 86, 323–331. [Google Scholar] [CrossRef]
- Cakmak, I. Enrichment of cereal grains with zinc: Agronomic or genetic biofortification? Plant Soil 2008, 302, 1–17. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Navarro, M.; Ashraf, M.; Akram, N.A.; Munné-Bosch, S. Nanofertilizer use for sustainable agriculture: Advantages and limitations. Plant Sci. 2019, 289, 110270. [Google Scholar] [CrossRef] [PubMed]
- Zafar, S.; Perveen, S.; Kamran Khan, M.; Shaheen, M.R.; Hussain, R.; Sarwar, N.; Rashid, S.; Nafees, M.; Farid, G.; Alamri, S.; et al. Effect of zinc nanoparticles seed priming and foliar application on the growth and physio-biochemical indices of spinach (Spinacia oleracea L.) under salt stress. PLoS ONE 2022, 17, e0263194. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Ahanger, M.A.; Egamberdieva, D.; Alam, P.; Alyemeni, M.N.; Ashraf, M. Modification of osmolytes and antioxidant enzymes by 24-epibrassinolide in chickpea seedlings under mercury (Hg) toxicity. J. Plant Growth Regul. 2018, 37, 309–322. [Google Scholar] [CrossRef]
- Nolan, T.M.; Vukašinović, N.; Liu, D.; Russinova, E.; Yin, Y. Brassinosteroids: Multidimensional regulators of plant growth, development, and stress responses. Plant Cell 2020, 32, 295–318. [Google Scholar] [CrossRef] [Green Version]
- Khan, I.; Awan, S.A.; Ikram, R.; Rizwan, M.; Akhtar, N.; Yasmin, H.; Sayyed, R.Z.; Ali, S.; Ilyas, N. Effects of 24-epibrassinolide on plant growth, antioxidants defense system, and endogenous hormones in two wheat varieties under drought stress. Physiol. Plant. 2021, 172, 696–706. [Google Scholar] [CrossRef]
- Desoky, E.S.M.; Mansour, E.; Ali, M.M.; Yasin, M.A.; Abdul-Hamid, M.I.; Rady, M.M.; Ali, E.F. Exogenously used 24-epibrassinolide promotes drought tolerance in maize hybrids by improving plant and water productivity in an arid environment. Plants 2021, 10, 354. [Google Scholar] [CrossRef]
- Yue, J.; Fu, Z.; Zhang, L.; Zhang, Z.; Zhang, J. The positive effect of different 24-epiBL pretreatments on salinity tolerance in Robinia pseudoacacia L. seedlings. Forests 2018, 10, 4. [Google Scholar] [CrossRef] [Green Version]
- Shin, Y.K.; Bhandari, S.R.; Jo, J.S.; Song, J.W.; Cho, M.C.; Yang, E.Y.; Lee, J.G. Response to salt stress in lettuce: Changes in chlorophyll fluorescence parameters, phytochemical contents, and antioxidant activities. Agronomy 2020, 10, 1627. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Rizwan, M.; Ali, S.; Ali, B.; Adrees, M.; Arshad, M.; Hussain, A.; ur Rehman, M.Z.; Waris, A.A. Zinc and iron oxide nanoparticles improved the plant growth and reduced the oxidative stress and cadmium concentration in wheat. Chemosphere 2019, 214, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Abou-Zeid, H.M.; Ismail, G.S.M.; Abdel-Latif, S.A. Influence of seed priming with ZnO nanoparticles on the salt-induced damages in wheat (Triticum aestivum L.) plants. J. Plant Nutr. 2021, 44, 629–643. [Google Scholar] [CrossRef]
- Aravind, P.; Prasad, M.N.V. Zinc protects chloroplasts and associated photochemical functions in cadmium exposed Ceratophyllum demersum L., a freshwater macrophyte. Plant Sci. 2004, 166, 1321–1327. [Google Scholar] [CrossRef]
- Raliya, R.; Nair, R.; Chavalmane, S.; Wang, W.N.; Biswas, P. Mechanistic evaluation of translocation and physiological impact of titanium dioxide and zinc oxide nanoparticles on the tomato (Solanum lycopersicum L.) plant. Metallomics 2015, 7, 1584–1594. [Google Scholar] [CrossRef]
- Rai-Kalal, P.; Jajoo, A. Priming with zinc oxide nanoparticles improve germination and photosynthetic performance in wheat. Plant Physiol. Biochem. 2021, 160, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Srivastav, A.; Ganjewala, D.; Singhal, R.K.; Rajput, V.D.; Minkina, T.; Voloshina, M.; Srivastava, S.; Shrivastava, M. Effect of ZnO nanoparticles on growth and biochemical responses of wheat and maize. Plants 2021, 10, 2556. [Google Scholar] [CrossRef]
- Wu, F.; Fang, Q.; Yan, S.; Pan, L.; Tang, X.; Ye, W. Effects of zinc oxide nanoparticles on arsenic stress in rice (Oryza sativa L.): Germination, early growth, and arsenic uptake. Environ. Sci. Pollut. Res. 2020, 27, 26974–26981. [Google Scholar] [CrossRef]
- Wang, X.; Yang, X.; Chen, S.; Li, Q.; Wang, W.; Hou, C.; Gao, X.; Wang, L.; Wang, S. Zinc oxide nanoparticles affect biomass accumulation and photosynthesis in Arabidopsis. Front. Plant Sci. 2016, 6, 1243. [Google Scholar] [CrossRef] [Green Version]
- Kohli, S.K.; Handa, N.; Sharma, A.; Kumar, V.; Kaur, P.; Bhardwaj, R. Synergistic effect of 24-epibrassinolide and salicylic acid on photosynthetic efficiency and gene expression in Brassica juncea L. under Pb stress. Turk. J. Biol. 2017, 41, 943–953. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.K. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Biol. 2003, 6, 441–445. [Google Scholar] [CrossRef]
- Abdelaziz, M.E.; Kim, D.; Ali, S.; Fedoroff, N.V.; Al-Babili, S. The endophytic fungus Piriformospora indica enhances Arabidopsis thaliana growth and modulates Na+/K+ homeostasis under salt stress conditions. Plant Sci. 2017, 263, 107–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seleiman, M.F.; Almutairi, K.F.; Alotaibi, M.; Shami, A.; Alhammad, B.A.; Battaglia, M.L. Nano-Fertilization as an Emerging Fertilization Technique: Why Can Modern Agriculture Benefit from Its Use? Plants 2020, 10, 2. [Google Scholar] [CrossRef] [PubMed]
- Noohpisheh, Z.; Amiri, H.; Mohammadi, A.; Farhadi, S. Effect of the foliar application of zinc oxide nanoparticles on some biochemical and physiological parameters of Trigonella foenum-graecum under salinity stress. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2021, 155, 267–280. [Google Scholar] [CrossRef]
- Dey, A.; Somaiah, S. Green synthesis and characterization of zinc oxide nanoparticles using leaf extract of Thryallis glauca (Cav.) Kuntze and their role as antioxidant and antibacterial. Microsc. Res. Tech. 2022, 85, 2835–2847. [Google Scholar] [CrossRef]
- El-Badri, A.M.; Batool, M.; Mohamed, I.A.; Khatab, A.; Sherif, A.; Wang, Z.; Salah, A.; Nishawy, E.; Ayaad, M.; Kuai, J.; et al. Modulation of salinity impact on early seedling stage via nano-priming application of zinc oxide on rapeseed (Brassica napus L.). Plant Physiol. Biochem. 2021, 166, 376–392. [Google Scholar] [CrossRef]
- Kapoor, D.; Rattan, A.; Gautam, V.; Bhardwaj, R. Mercury-induced changes in growth, metal & ions uptake, photosynthetic pigments, osmoprotectants and antioxidant defence system in Raphanus sativus L. seedlings and role of steroid hormone in stress amelioration. J. Pharmacogn. Phytochem. 2016, 5, 259. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Salinity | Root Application Treatments | Total Root Length (cm) | Total Surface Area (cm2) | Average Diameter (mm) | Total Root Volume (cm3) | Number of Root Tips (Number) |
|---|---|---|---|---|---|---|
| Control | T1 | 296.90 | 256.91 | 1.84 | 10.96 | 1237.33 |
| T2 | 326.38 | 291.12 | 1.94 | 12.89 | 1341.33 | |
| T3 | 421.72 | 320.55 | 2.11 | 16.92 | 1681.33 | |
| T4 | 365.01 | 310.44 | 1.82 | 15.40 | 1504.33 | |
| T5 | 443.80 | 343.66 | 2.25 | 17.46 | 1788.67 | |
| T6 | 456.95 | 377.89 | 2.47 | 28.41 | 2061.67 | |
| T7 | 744.46 | 605.64 | 3.03 | 37.02 | 2875.00 | |
| T8 | 531.32 | 482.50 | 2.53 | 31.90 | 2734.00 | |
| T9 | 617.61 | 582.55 | 2.77 | 30.02 | 2680.00 | |
| Mean | 467.13 A | 396.81 A | 2.31 A | 22.33 A | 1989.30 A | |
| 5 dS m−1 | T1 | 267.45 | 238.95 | 1.62 | 9.71 | 987.67 |
| T2 | 294.13 | 258.94 | 1.77 | 11.16 | 1107.33 | |
| T3 | 367.04 | 300.16 | 1.82 | 14.96 | 1411.00 | |
| T4 | 340.61 | 270.88 | 1.56 | 13.74 | 1239.33 | |
| T5 | 403.98 | 316.25 | 1.96 | 14.72 | 1537.67 | |
| T6 | 421.57 | 341.63 | 2.07 | 23.48 | 1781.33 | |
| T7 | 676.89 | 521.96 | 2.62 | 33.19 | 2601.00 | |
| T8 | 490.44 | 407.14 | 2.18 | 26.51 | 2373.00 | |
| T9 | 565.68 | 495.65 | 2.23 | 27.03 | 2225.33 | |
| Mean | 425.31 B | 350.17 B | 1.98 B | 19.39 B | 1695.96 B | |
| 10 dS m−1 | T1 | 173.29 | 125.80 | 1.13 | 6.66 | 787.67 |
| T2 | 235.30 | 157.41 | 1.37 | 9.68 | 914.00 | |
| T3 | 304.28 | 244.85 | 1.56 | 12.26 | 1168.00 | |
| T4 | 283.29 | 235.25 | 1.28 | 11.54 | 1025.33 | |
| T5 | 377.46 | 268.16 | 1.67 | 12.82 | 1366.67 | |
| T6 | 378.36 | 296.04 | 1.64 | 18.71 | 1457.00 | |
| T7 | 607.78 | 469.62 | 2.35 | 28.65 | 2340.00 | |
| T8 | 448.04 | 323.14 | 1.78 | 20.72 | 1954.67 | |
| T9 | 450.56 | 342.41 | 1.89 | 18.75 | 1740.00 | |
| Mean | 362.04 C | 273.63 C | 1.63 C | 15.53 C | 1417.04 C | |
| Significance ANOVA | ||||||
| Saline Stress | *** | ** | *** | *** | *** | |
| Seed Priming Treatment | *** | *** | ** | ** | * | |
| Interaction | *** | ** | * | *** | ** | |
| SEM0.05 | ||||||
| Saline Stress | 3.040 | 3.279 | 0.017 | 0.161 | 12.969 | |
| Seed Priming Treatment | 5.265 | 5.679 | 0.029 | 0.279 | 22.462 | |
| Interaction | 9.119 | 9.837 | 0.051 | 0.483 | 38.906 | |
| Salinity | Root Application Treatments | Stem Diameter (mm) | Shoot Length (cm) | SLW (mg/cm2) | Leaf Area (cm2) | RWC (%) |
|---|---|---|---|---|---|---|
| Control | T1 | 9.77 | 98.90 | 3.55 | 80.46 | 86.23 |
| T2 | 8.80 | 105.67 | 2.80 | 107.33 | 87.48 | |
| T3 | 8.10 | 99.77 | 2.95 | 118.64 | 88.98 | |
| T4 | 11.03 | 103.07 | 3.44 | 153.81 | 87.12 | |
| T5 | 12.27 | 108.40 | 3.26 | 188.35 | 88.29 | |
| T6 | 12.27 | 118.87 | 3.45 | 182.20 | 90.05 | |
| T7 | 13.07 | 126.33 | 3.04 | 233.06 | 90.55 | |
| T8 | 12.57 | 121.60 | 3.38 | 202.46 | 89.22 | |
| T9 | 10.03 | 115.03 | 2.66 | 212.88 | 90.04 | |
| Mean | 10.88 A | 110.85 A | 3.17 A | 164.35 A | 88.66 A | |
| 5 dS m−1 | T1 | 9.00 | 70.60 | 3.70 | 94.64 | 82.14 |
| T2 | 8.47 | 81.70 | 1.99 | 172.51 | 83.89 | |
| T3 | 8.57 | 88.07 | 1.85 | 142.00 | 85.18 | |
| T4 | 10.23 | 94.47 | 3.47 | 128.15 | 87.07 | |
| T5 | 10.43 | 84.73 | 2.32 | 155.81 | 89.41 | |
| T6 | 9.57 | 81.83 | 2.90 | 174.21 | 88.27 | |
| T7 | 10.27 | 100.10 | 3.08 | 206.80 | 90.12 | |
| T8 | 9.50 | 85.70 | 2.79 | 187.54 | 88.05 | |
| T9 | 9.13 | 85.03 | 2.92 | 138.59 | 89.69 | |
| Mean | 9.46 B | 85.80 B | 2.78 B | 155.58 B | 87.09 B | |
| 10 dS m−1 | T1 | 5.10 | 50.93 | 2.69 | 34.47 | 69.97 |
| T2 | 5.07 | 67.43 | 2.02 | 64.35 | 73.66 | |
| T3 | 7.03 | 62.13 | 2.49 | 66.68 | 72.44 | |
| T4 | 7.23 | 57.43 | 2.72 | 74.04 | 77.16 | |
| T5 | 7.30 | 60.50 | 2.00 | 105.08 | 78.62 | |
| T6 | 7.53 | 77.63 | 3.09 | 137.13 | 82.62 | |
| T7 | 7.87 | 80.03 | 3.25 | 148.57 | 87.37 | |
| T8 | 7.23 | 88.53 | 2.78 | 156.37 | 86.16 | |
| T9 | 6.57 | 80.30 | 1.69 | 152.81 | 84.78 | |
| Mean | 6.77 C | 69.44 C | 2.53 C | 104.39 C | 79.20 C | |
| Significance ANOVA | ||||||
| Saline Stress | ** | *** | *** | *** | ** | |
| Seed Priming | *** | ** | ** | ** | *** | |
| Interaction | *** | *** | * | *** | ** | |
| SEM0.05 | ||||||
| Saline Stress | 0.093 | 0.574 | 0.045 | 2.537 | 0.231 | |
| Seed Priming Treatment | 0.161 | 0.994 | 0.077 | 4.395 | 0.399 | |
| Interaction | 0.278 | 1.721 | 0.134 | 7.612 | 0.692 | |
| Salinity | NPs/EBL | Total Chlorophyll (mg/gFW) | Carotenoids (mg/gFW) | Phenolic Content (mgGAE/gFM) | Proline (mg/gFW) |
|---|---|---|---|---|---|
| Control | T1 | 4.53 | 0.35 | 40.25 | 0.37 |
| T2 | 4.57 | 0.36 | 41.56 | 0.37 | |
| T3 | 4.89 | 0.38 | 40.99 | 0.37 | |
| T4 | 4.77 | 0.37 | 43.04 | 0.37 | |
| T5 | 4.95 | 0.39 | 45.44 | 0.36 | |
| T6 | 4.89 | 0.37 | 43.11 | 0.36 | |
| T7 | 5.18 | 0.40 | 45.85 | 0.34 | |
| T8 | 4.97 | 0.38 | 42.59 | 0.36 | |
| T9 | 5.00 | 0.37 | 42.78 | 0.36 | |
| Mean | 4.86 A | 0.37 A | 42.85 B | 0.36 C | |
| 5 dS m−1 | T1 | 3.79 | 0.29 | 47.28 | 2.05 |
| T2 | 4.29 | 0.31 | 50.91 | 1.35 | |
| T3 | 4.61 | 0.36 | 54.61 | 0.76 | |
| T4 | 4.68 | 0.34 | 55.87 | 0.85 | |
| T5 | 4.80 | 0.36 | 64.13 | 0.70 | |
| T6 | 4.63 | 0.35 | 56.87 | 0.67 | |
| T7 | 5.03 | 0.38 | 69.71 | 0.62 | |
| T8 | 4.83 | 0.35 | 66.23 | 0.71 | |
| T9 | 4.76 | 0.34 | 66.82 | 0.76 | |
| Mean | 4.60 B | 0.34 B | 59.16 A | 0.94 B | |
| 10 dS m−1 | T1 | 3.03 | 0.21 | 24.33 | 2.60 |
| T2 | 3.80 | 0.28 | 28.58 | 1.67 | |
| T3 | 4.44 | 0.31 | 32.36 | 1.19 | |
| T4 | 4.14 | 0.30 | 35.15 | 0.95 | |
| T5 | 4.50 | 0.32 | 36.17 | 0.82 | |
| T6 | 4.11 | 0.29 | 35.79 | 0.75 | |
| T7 | 4.84 | 0.34 | 41.27 | 0.69 | |
| T8 | 4.54 | 0.31 | 35.20 | 0.75 | |
| T9 | 4.28 | 0.31 | 37.37 | 0.73 | |
| Mean | 4.19 C | 0.30 C | 34.03 C | 1.13 A | |
| Significance ANOVA | |||||
| Saline Stress | ** | *** | *** | *** | |
| Seed Priming | *** | ** | *** | *** | |
| Interaction | * | ** | *** | ** | |
| SEM0.05 | |||||
| Saline Stress | 0.014 | 0.001 | 0.297 | 0.007 | |
| Seed Priming | 0.024 | 0.002 | 0.514 | 0.013 | |
| Interaction | 0.042 | 0.004 | 0.890 | 0.022 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seleiman, M.F.; Ahmad, A.; Alshahrani, T.S. Integrative Effects of Zinc Nanoparticle and PGRs to Mitigate Salt Stress in Maize. Agronomy 2023, 13, 1655. https://doi.org/10.3390/agronomy13061655
Seleiman MF, Ahmad A, Alshahrani TS. Integrative Effects of Zinc Nanoparticle and PGRs to Mitigate Salt Stress in Maize. Agronomy. 2023; 13(6):1655. https://doi.org/10.3390/agronomy13061655
Chicago/Turabian StyleSeleiman, Mahmoud F., Awais Ahmad, and Thobayet S. Alshahrani. 2023. "Integrative Effects of Zinc Nanoparticle and PGRs to Mitigate Salt Stress in Maize" Agronomy 13, no. 6: 1655. https://doi.org/10.3390/agronomy13061655
APA StyleSeleiman, M. F., Ahmad, A., & Alshahrani, T. S. (2023). Integrative Effects of Zinc Nanoparticle and PGRs to Mitigate Salt Stress in Maize. Agronomy, 13(6), 1655. https://doi.org/10.3390/agronomy13061655