

Diversity Assessment of Winged Bean [Psophocarpus tetragonolobus (L.) DC.] Accessions from IITA Genebank

 ,
,  , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Germplasm and Experimental Site
Seed Viability Assessment
2.2. Agro-Morphological Characterization
2.3. Data Collection
2.4. Statistical Analyses
2.5. Molecular Characterization
3. Results
3.1. Variability in Agronomic Traits of 15 Winged Bean Accession
3.2. Genetic Variances and Broad-Sense Heritability of Agronomic Traits
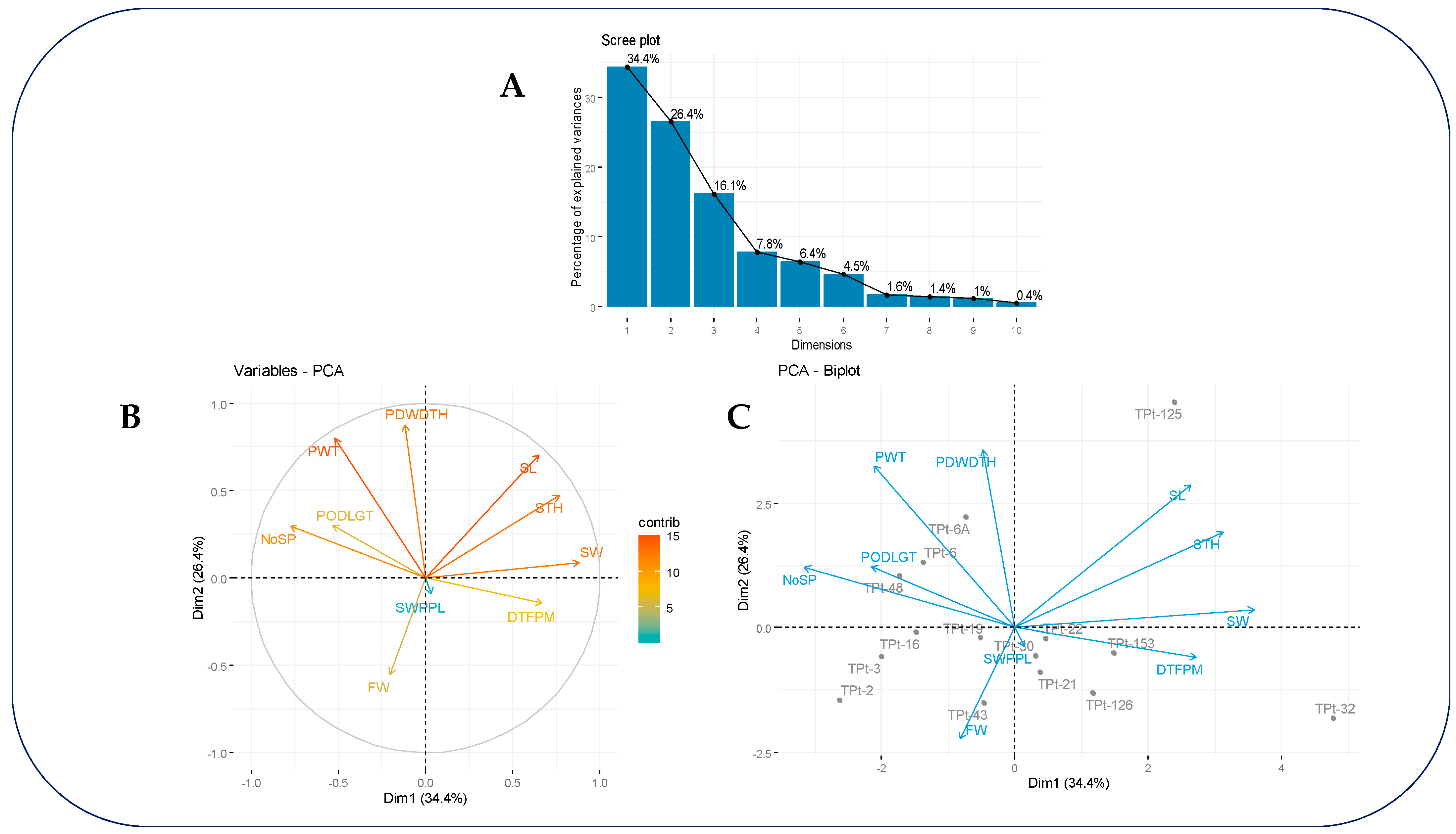
3.3. Dimension Reduction Analysis of Agronomic Traits
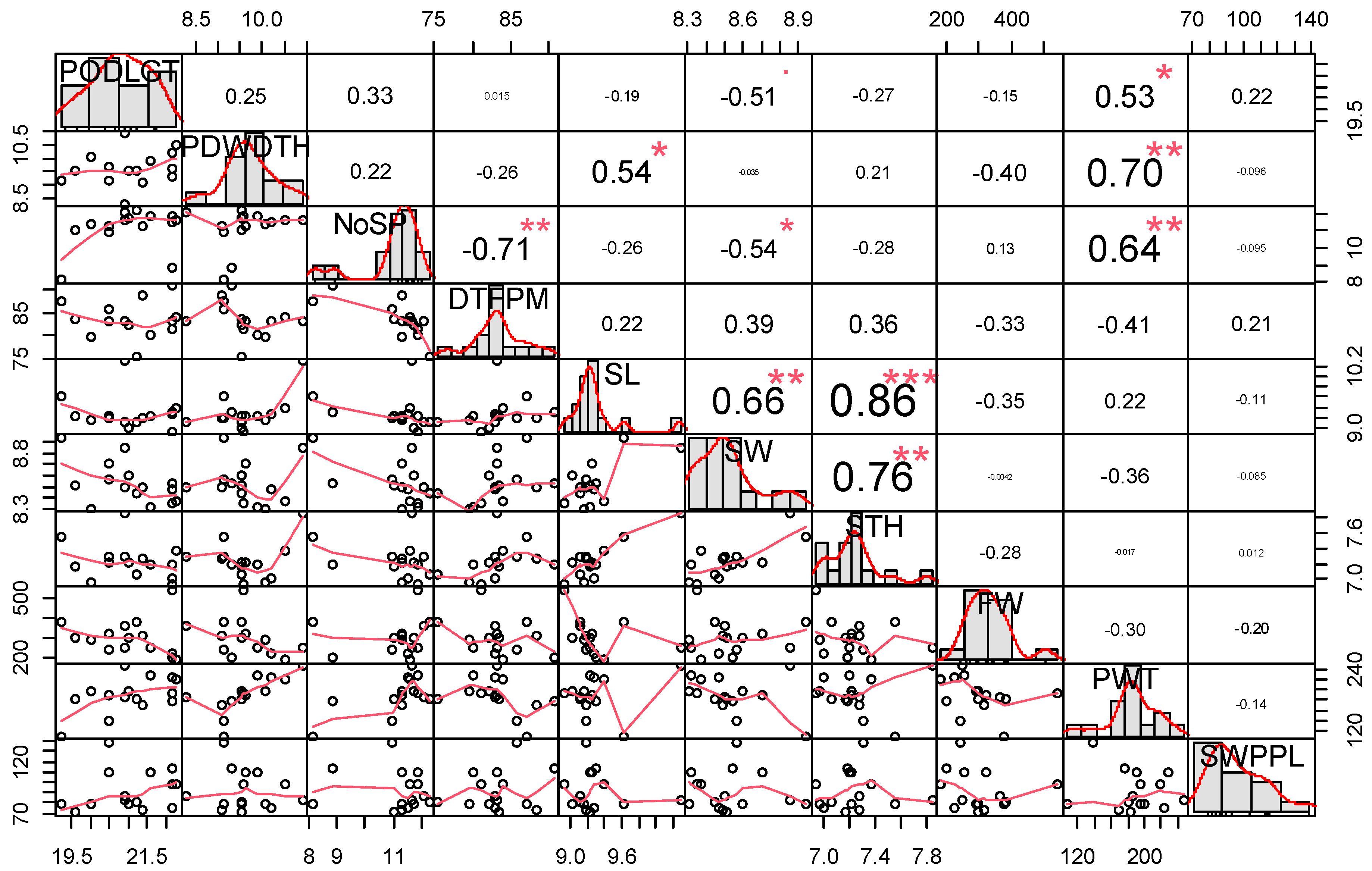
3.4. Phenotypic Correlation among Measured Agronomic Traits
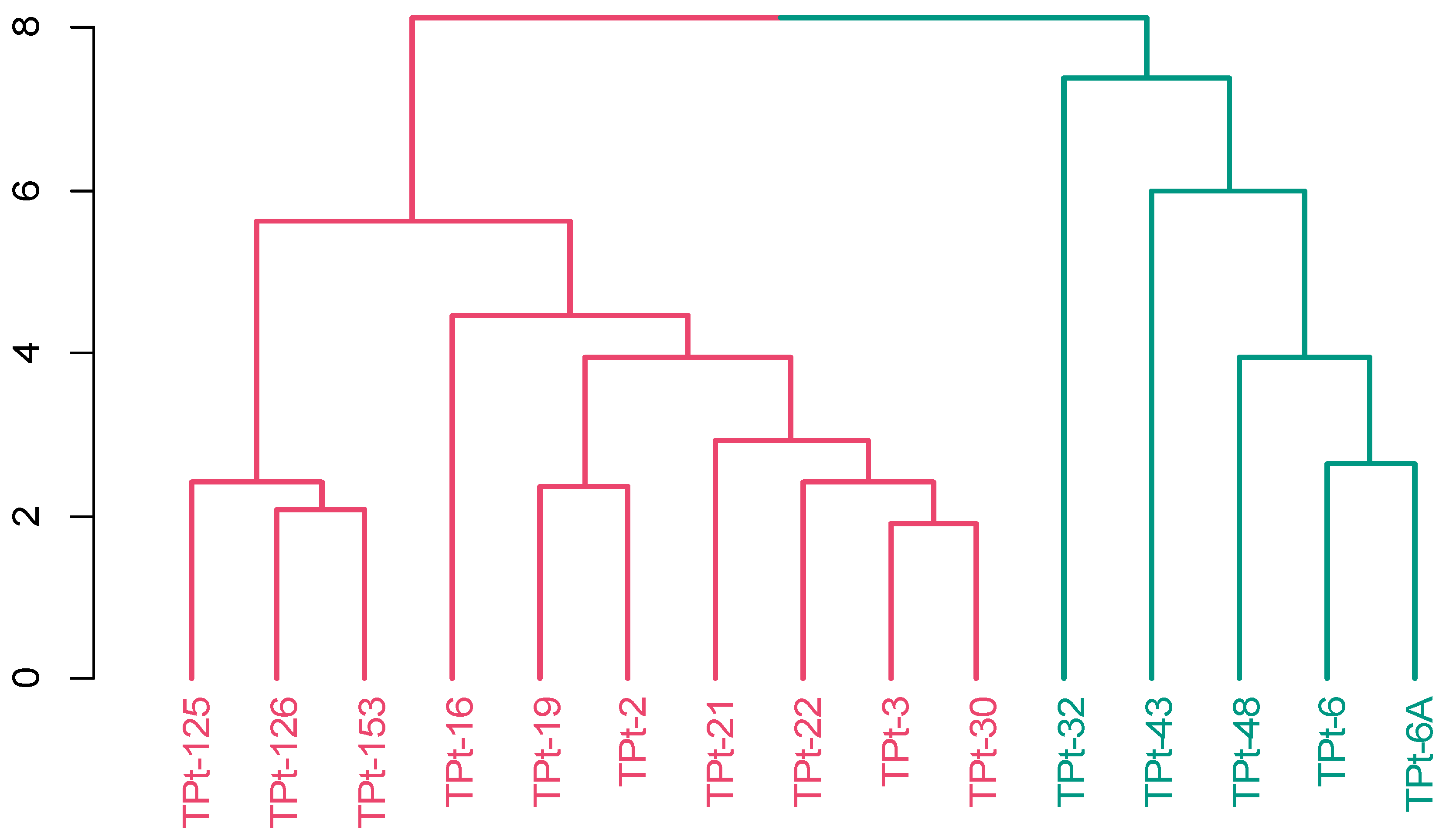
3.5. Phenotypic Diversity among 15 Studied Winged Bean Accessions Based on Gower’s Distance
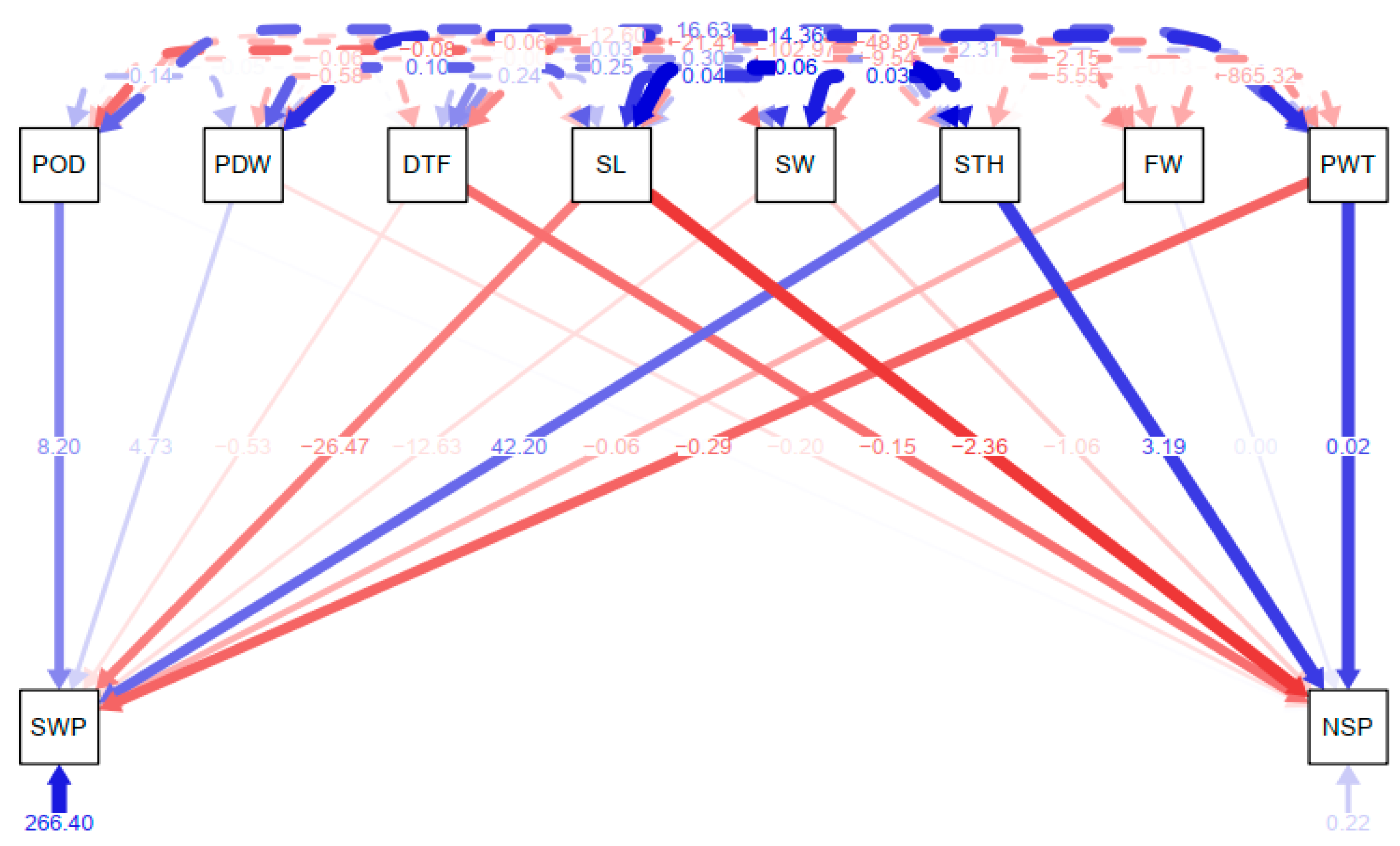
3.6. Path Coefficient Analysis for Correlated Agronomic Traits
3.7. Genetic Diversity among the 15 Winged Bean Accessions
3.7.1. Polymorphisms Detected by Simple Sequence Repeats (SSRs)
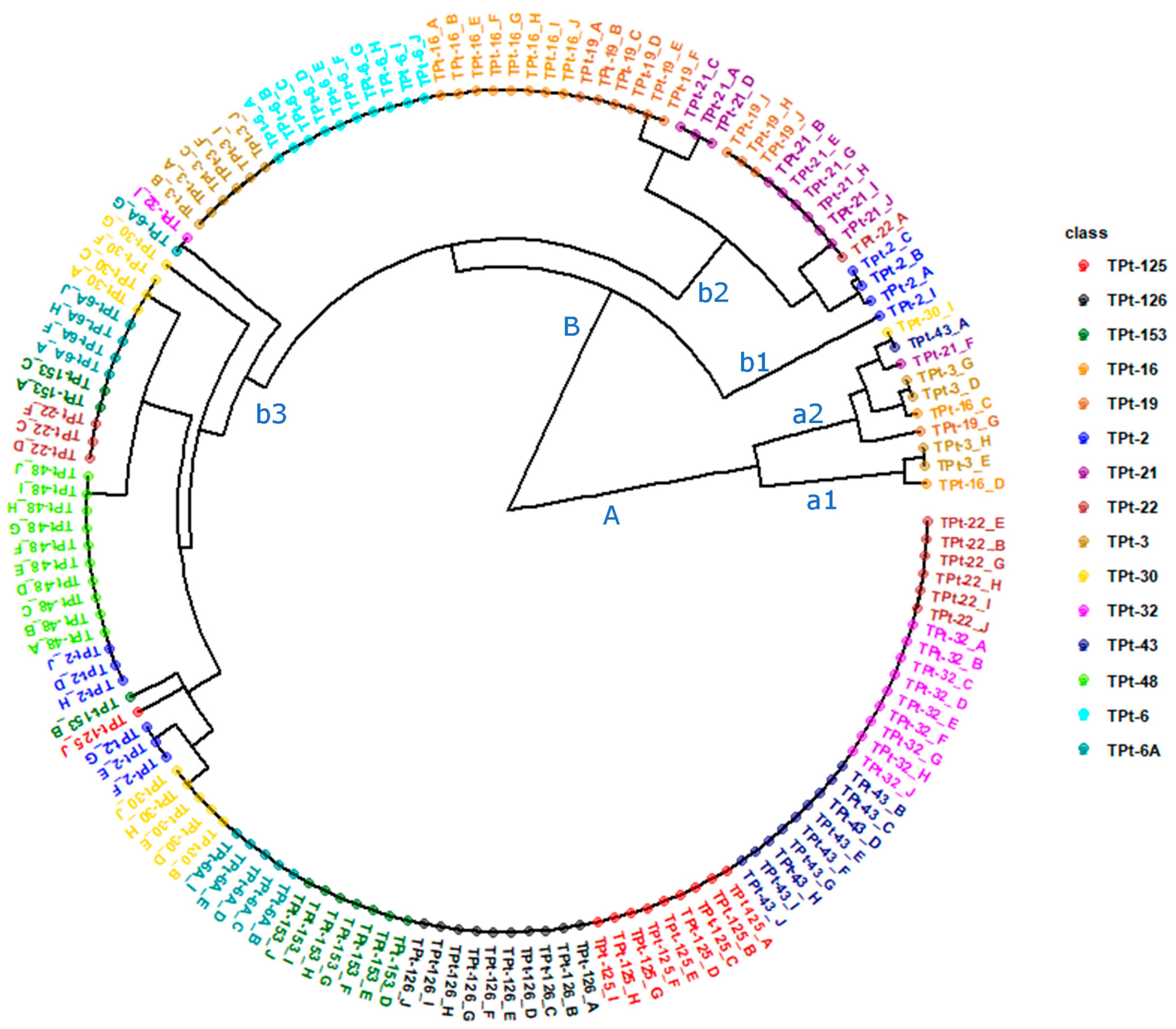
3.7.2. Analysis of Molecular Variance (AMOVA) and Cluster Analysis
4. Discussion
4.1. Variability in Measured Traits as Identifiers of Gene Reservoirs for Winged Bean Improvement
4.2. Potentials of Measured Traits for Indirect Selection in Winged Bean Improvement
4.3. SSR Markers Revealed Intra-Accession Genetic Variation within the Winged Bean Germplasm
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tanzito, G.; Ibanda, P.A.; Ocan, D.; Lejoly, J. Use of charcoal (biochar) to enhance tropical soil fertility: A case of Masako in Democratic Republic of Congo. J. Soil Sci. Environ. Manag. 2020, 11, 17–29. [Google Scholar] [CrossRef]
- Sriwichai, S.; Monkham, T.; Sanitchon, J.; Jogloy, S.; Chankaew, S. Dual-Purpose of the Winged Bean (Psophocarpus tetragonolobus (L.) DC.), the Neglected Tropical Legume, Based on Pod and Tuber Yields. Plants 2021, 10, 1746. [Google Scholar] [CrossRef] [PubMed]
- Aletor, V.A.; Aladetimin, O.O. Proximate composition of some underutilized Nigeria legumes. Nahrung 1989, 33, 999–1007. [Google Scholar] [CrossRef] [PubMed]
- Koshy, E.P.; John, P.; Scaria, S. Winged bean: The wings that carry away malnutrition. Acad. Rev. 1999, 3, 77–80. [Google Scholar]
- Adegboyega, T.T.; Abberton, M.T.; AbdelGadir, A.H.; Dianda, M.; Maziya-Dixon, B.; Oyatomi, O.A.; Ofodile, S.; Babalola, O.O. Evaluation of Nutritional and Antinutritional Properties of African Yam Bean (Sphenostylis stenocarpa (Hochst ex. A. Rich.) Harms.) Seeds. J. Food Qual. 2020, 2020, 6569420. [Google Scholar] [CrossRef]
- Wong, Q.N.; Tanzi, A.S.; Ho, W.K.; Malla, S.; Blythe, M.; Karunaratne, A.; Massawe, F.; Mayes, S. Development of Gene-Based SSR Markers in Winged Bean (Psophocarpus tetragonolobus (L.) DC.) for Diversity Assessment. Genes 2017, 8, 100. [Google Scholar] [CrossRef]
- Mohanty, C.S.; Verma, S.; Singh, V.; Khan, S.; Gaur, P.; Gupta, P.; Nizar, M.A.; Dikshit, N.; Pattanayak, R.; Shukla, A.; et al. Characterization of winged bean (Psophocarpus tetragonolobus (L.) DC.) based on molecular, chemical and physiological parameters. Am. J. Mol. Biol. 2013, 03, 187–197. [Google Scholar] [CrossRef]
- Gupta, S.K.; Bansal, R.; Gopalakrishna, T. Development and characterization of genic SSR markers for mungbean (Vigna radiata (L.) Wilczek). Euphytica 2013, 195, 245–258. [Google Scholar] [CrossRef]
- Egan, A.N.; Schlueter, J.; Spooner, D.M. Applications of next-generation sequencing in plant biology. Am. J. Bot. 2012, 99, 175–185. [Google Scholar] [CrossRef]
- Omena, B.O.; Nkang, N.A.; Odesola, A.; Kafilat, D.; Okeh, I. Genetic diversity assessment of winged bean (pso-phorcarpus tetragonolobus) accessions revealed by Inter-Simple Sequence Repeat (ISSR) markers. J. Plant Biol. Crop Res. 2020, 3, 1014. [Google Scholar]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Kresovich, S. Quantitative genetics in maize breeding. Field Crops Res. 1990, 23, 78–79. [Google Scholar] [CrossRef]
- Johnson, H.W.; Robinson, H.F.; Comstock, R.E. Genotypic and Phenotypic Correlations in Soybeans and Their Implications in Selection 1. Agron. J. 1955, 47, 477–483. [Google Scholar] [CrossRef]
- Deshmukh, S.; Basu, M.; Reddy, P. Genetic variability, character association and path coefficients of quantitative traits in Virginia bunch varieties of groundnut. Indian J. Agric. Sci. 1986, 56, 816–821. [Google Scholar]
- Kassambara, A.; Mundt, F. Package ‘Factoextra’, CRAN-R Packag. 2020. Available online: http://cran.nexr.com/web/packages/factoextra/factoextra.pdf (accessed on 4 April 2023).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Galili, T. Dendextend: An R package for visualizing, adjusting, and comparing trees of hierarchical clustering. Bioinformatics 2015, 31, 3718–3720. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Gu, L.; Eils, R.; Schlesner, M.; Brors, B. circlize implements and enhances circular visualization in R. Bioinformatics 2014, 30, 2811–2812. [Google Scholar] [CrossRef]
- Charrad, M.; Ghazzali, N.; Boiteau, V.; Maintainer, A.N. Determining the Best Number of Clusters in a Data Set. Cran. 2015. Submitted 13 April 2015; published 2nd May 2022 13:01:42 UTC. Available online: https://www.google.com.hk/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&ved=2ahUKEwiPko76jI2AAxVO6WEKHR5aCDIQFnoECBoQAQ&url=https%3A%2F%2Fcran.r-project.org%2Fweb%2Fpackages%2FNbClust%2FNbClust.pdf&usg=AOvVaw0_6opCPIS9Hd4qstjJ_HW0&opi=89978449 (accessed on 4 April 2023).
- Rosseel, Y. lavaan: AnRPackage for Structural Equation Modeling. J. Stat. Softw. 2012, 48, 1–36. [Google Scholar] [CrossRef]
- Epskamp, S. semPlot: Unified Visualizations of Structural Equation Models. Struct. Equ. Model. A Multidiscip. J. 2015, 22, 474–483. [Google Scholar] [CrossRef]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A plant DNA mini preparation: Version II. Plant Mol. Biol. Report. 1983, 1, 19–21. [Google Scholar] [CrossRef]
- Vatanparast, M.; Shetty, P.; Chopra, R.; Doyle, J.J.; Sathyanarayana, N.; Egan, A.N. Transcriptome sequencing and marker development in winged bean (Psophocarpus tetragonolobus; Leguminosae). Sci. Rep. 2016, 6, 29070. [Google Scholar] [CrossRef]
- Chatterji, S.; Pachter, L. Reference based annotation with GeneMapper. Genome Biol. 2006, 7, R29. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Muse, S.V. Power Marker integrated analysis environment for genetic marker data. Bioinformatics 2005, 29, 2128–2129. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Smouse, P. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Tanzi, A.S.; Ho, W.K.; Massawe, F.; Mayes, S. Development and interaction between plant architecture and yield-related traits in winged bean (Psophocarpus tetragonolobus (L.) DC.). Euphytica 2019, 215, 36. [Google Scholar] [CrossRef]
- Schiavinato, I.V.M.A. Influence of photoperiod and temperature on the development of winged bean plants. RBrasFisiolVeg 1996, 8, 105–110. [Google Scholar]
- Adejumobi, I.; Agre, A.P.; Onautshu, O.D.; Adheka, G.J.; Cipriano, M.I.; Jean-Claude, L.K.; Joseph, L.M. Assessment of yam landraces (Dioscorea spp.) of DR Congo for reaction to pathological diseases, yield potential and tuber quality characteristics. Agronomy 2022, 12, 599. [Google Scholar] [CrossRef]
- Amoo, I.A.; Adebayo, O.T.; Oyeleye, A.O. Chemical evaluation of winged beans. Afr. J. Food Agric. Nutr. Dev. 2006, 6. [Google Scholar] [CrossRef]
- Gupta, S.K.; Gopalakrishna, T. Development of unigene-derived SSR markers in cowpea (Vigna unguiculata) and their transferability to other Vigna species. Genome 2010, 53, 508–523. [Google Scholar] [CrossRef]
- Li, G.; Ra, W.-H.; Park, J.-W.; Kwon, S.-W.; Lee, J.-H.; Park, C.-B.; Park, Y.-J. Developing EST-SSR markers to study molecular diversity in Liriope and Ophiopogon. Biochem. Syst. Ecol. 2011, 39, 241–252. [Google Scholar] [CrossRef]
- Dutta, S.; Kumawat, G.; Singh, B.P.; Gupta, D.K.; Singh, S.; Dogra, V.; Gaikwad, K.; Sharma, T.R.; Raje, R.S.; Bandhopadhya, T.K.; et al. Development of genic-SSR markers by deep transcriptome sequencing in pigeonpea [Cajanus cajan (L.) Millspaugh]. BMC Plant Biol. 2011, 11, 17. [Google Scholar] [CrossRef]
- Blair, M.W.; Hurtado, N.; Chavarro, C.M.; Muñoz-Torres, M.C.; Giraldo, M.C.; Pedraza, F.; Tomkins, J.; Wing, R. Gene-based SSR markers for common bean (Phaseolus vulgaris L.) derived from root and leaf tissue ESTs: An integration of the BMc series. BMC Plant Biol. 2011, 11, 50. [Google Scholar] [CrossRef] [PubMed]
- Molosiwa, O.O.; Aliyu, S.; Stadler, F.; Mayes, K.; Massawe, F.; Kilian, A.; Mayes, S. SSR marker development, genetic diversity and population structure analysis of Bambara groundnut [Vigna subterranea (L.) Verdc.] landraces. Genet. Resour. Crop Evol. 2015, 62, 1225–1243. [Google Scholar] [CrossRef]
- Adebayo, M.A.; Kolawole, A.O.; Adebayo, T.A. Assessment of new generation of drought-tolerant maize (Zea mays L.) hybrids for agronomic potential and adaptation in the derived savanna agro-ecology of Nigeria. Int. J. Agron. Agric. Res. 2015, 7, 45–54. [Google Scholar]
- Adebayo, M.A.; Menkir, A.; Gedil, M.; Blay, E.; Gracen, V.; Danquah, E.; Funmilayo, L. Diversity assessment of drought tolerant exotic and adapted maize (Zea mays L.) inbred lines with microsatellite markers. J. Crop Sci. Biotechnol. 2015, 18, 147–154. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S/N | Accession No | Source | SC | SS | FLC | STEMCLR | PPS | PPS | LSS |
|---|---|---|---|---|---|---|---|---|---|
| 1 | TPt-2 | Nigeria | Brownish orange | Oval | Pastel violet | Green | absent | Flat on suture | Deltoid-large |
| 2 | TPt-3 | Nigeria | Yellowish brown | Oval | Light violet | purple | present | Flat on suture | Ovate Lanceolate-medium |
| 3 | TPt-6 | Nigeria | Yellowish brown | Oval | Pastel violet | Green | present | Flat on side | Ovate-large |
| 4 | TPt-16 | Indonesia | Brownish orange | Round | Light violet | Green | absent | Flat on side | Ovate Lanceolate-large |
| 5 | TPt-19 | Nigeria | Yellowish brown | Oval | Pale blue | Green | absent | Flat on suture | Deltoid-large |
| 6 | TPt-21 | Papua New Guinea | Violet brown | Round | Light violet | Green | absent | Flat on sides | Deltoid-medium |
| 7 | TPt-22 | Papua New Guinea | Brownish Yellow | Round | Pastel violet | purple | absent | Flat on sides | Deltoid-large |
| 8 | TPt-32 | Unknown | Yellowish brown | Oval | Pale violet | Green | absent | Flat on suture | Deltoid-large |
| 9 | TPt-43 | Unknown | Tan | Oval | Light violet | Greenish purple | present | Flat on sides | Deltoid-large |
| 10 | TPt-48 | Unknown | Yellowish brown | Oval | Pale violet | Green | present | Flat on suture | Deltoid-large |
| 11 | TPt-125 | Unknown | Tan | Oval | Pastel violet | Green | absent | Flat on suture | Deltoid-large |
| 12 | TPt-126 | Unknown | Yellowish brown | Oval | Light violet | Green | absent | Flat on sides | Deltoid-large |
| 13 | TPt-153 | Unknown | Light brown | Oval | Light violet | Greenish purple | absent | Flat on sides | Deltoid-large |
| 14 | TPt-6A | Nigeria | Brownish orange | Oval | Light violet | Green | absent | Flat on suture | Deltoid-large |
| 15 | TPt-30 | Unknown | Brownish orange | Round | Pastel violet | Green | absent | Flat on sides | Deltoid-large |
| S/N | Traits | Description of Measurement | Collection Period |
|---|---|---|---|
| 1 | Days to First Flower (DTFF) | number of days from planting to when a plant in a plot emerged first flower | 6 WAP |
| 2 | Days to First Pod (DTFP), | number of days from planting to when a plant in a plot emerged first Pod | 8 WAP |
| 3 | Days to 50% Flower (DT5F) | number of days from planting to when 50% of the plants in a plot emerged flower | 6–8 WAP |
| 4 | Vine length (VL7WAP) | measured as the distance between the stem and the last leaf at the top node | 6–7 WAP |
| 5 | Number of pods per peduncle (NPPP) | counting the number of pods for tagged plant on a plot | 8–12 WAP |
| 6 | Pod length (PODLGTH) | measured from the point of attachment to the tip of the pod | At Maturity |
| 7 | Pod width (PODWDTH) | measured from the edge of one wing to that of the opposite wing at the middle of the pod | At Maturity |
| 8 | Number of seeds per pods (NSP) | Counted and averaged over ten tagged plants in a plot. | At Harvest |
| 9 | Seed weight (SW) | measured using a sensitive digital scale as mean weight of ten dry seeds | At Harvest |
| 10 | Seed thickness (STH) | measured using a Vennier caliper as mean thickness of ten dry seeds | At Harvest |
| 11 | Seed length (SL) | measured using a Vennier caliper as mean length of ten dry seeds | At Harvest |
| 12 | Seed width (SDTHW) | measured using a Vennier caliper as mean width of ten dry seeds | At Harvest |
| 13 | Fodder weight (FW) | measured as the weight of leaf mass or abundance of leaf mass at maturity | At Harvest |
| SSR Primer Name | Dyes | 5′ Forward Sequence 3′ | 5′ Reverse Sequence 3′ |
|---|---|---|---|
| SSR-24 | 6-Fam | ACC TCA TAG AGG AAT ACG AC | CAA TAT GTG GAG GAA GTA GA |
| SSR-704 | Atto-532 | GAT TGT TGT GAG ATT GAA GT | ATG CAA ATA GCT TAC AAA AG |
| SSR-747 | 6-Fam | ACT TTG TGA AAA TGA AGG TA | AAT TTA ATA TGG CTG CTA AA |
| SSR-854 | Atto-532 | CTC TAA AAT TCT CAC ACT CG | CGA ATT TCT TTC AAT TCT TA |
| SSR-860 | Atto-532 | TGA GGA AAA TAA AAA GAA AA | CGA GTG TGA GAA TTT TAG AG |
| SSR-879 | Atto-565 | GCA ACA CTT TAG CTC ATT AT | GAA CTT CAA CAC TAT TCC AA |
| SSR-1104 | Atto-565 | CTT CAA CTG CTT GTT CTA CT | TAA AGA AGA AAG AGG AAA GG |
| SSR-3111 | 6-Fam | AGT TGG AAA GTA GCA GAG TT | GGT GTG AGA AGC ATA ATA AA |
| SSR-5819 | Atto-550 | AAT AAT GTC AAT TAC GCA GT | GAA CTG AAG CCA TGT AGT AG |
| SSR-11100 | Atto-550 | AAT AGA AGG CTT GGT GTC | CTT CCT CTT CTC TTC GTC T |
| Source | DF | PODLGT | PDWDTH | NoSP | DTFPM | SL | SW | STH | FW | PWT | SWPPL |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Accessions | 14 | 5.45 * | 2.42 ** | 3.12 ** | 83.78 * | 0.62 * | 0.20 ** | 0.33 * | 49,038 * | 7227.3 *** | 2265.16 *** |
| Year | 1 | 2.99 | 1736.4 *** | 2.46 | 551.19 * | 0.337 | 0.024 | 0.10 | 36 | 788 | 46.06 |
| Accessions * Year | 14 | 3.21 | 1.71 | 1.74 | 75.14 * | 0.515 | 0.122 | 0.27 | 10,927 | 2033.2 | 705.95 * |
| Residual | 1.54 | 0.99 | 1.59 | 6.16 | 0.574 | 0.283 | 0.41 | 145.06 | 41.546 | 18.75 | |
| CV | 7.36 | 10.52 | 14.63 | 7.39 | 6.16 | 3.29 | 5.76 | 48.04 | 21.95 | 20.85 | |
| Mean | 21.04 | 9.63 | 11.16 | 83.39 | 9.28 | 8.53 | 7.24 | 304.32 | 189.39 | 91.39 | |
| δ2g | 0.37 | 0.12 | 0.59 | 1.44 | 0.02 | 0.01 | 0.01 | 5003.26 | 875.00 | 260.87 | |
| δ2p | 0.91 | 0.40 | 1.32 | 13.96 | 0.10 | 0.03 | 0.06 | 8173.12 | 1204.00 | 377.53 | |
| GCV (%) | 2.90 | 3.57 | 6.86 | 1.44 | 1.51 | 1.40 | 1.42 | 23.24 | 15.62 | 17.67 | |
| PCV (%) | 4.53 | 6.60 | 10.31 | 4.48 | 3.47 | 2.15 | 3.26 | 29.71 | 18.32 | 21.26 | |
| H2 (%) | 40.99 | 29.26 | 44.31 | 10.31 | 18.92 | 42.75 | 18.83 | 61.22 | 72.67 | 69.10 |
| Variable | PC1 | PC2 | PC3 |
|---|---|---|---|
| PODLGT | −0.529 | 0.301 | 0.559 |
| PDWDTH | −0.118 | 0.877 | −0.046 |
| NoSP | −0.775 | 0.295 | −0.262 |
| DTFPM | 0.667 | −0.147 | 0.574 |
| SL | 0.648 | 0.703 | −0.168 |
| SW | 0.881 | 0.088 | −0.271 |
| STH | 0.767 | 0.472 | −0.115 |
| FW | −0.202 | −0.550 | −0.550 |
| PWT | −0.519 | 0.799 | −0.010 |
| SWPPL | 0.035 | −0.093 | 0.694 |
| Eigen value | 3.436 | 2.642 | 1.612 |
| Percentage of variance (%) | 34.357 | 26.424 | 16.117 |
| Cumulative of variance (%) | 34.357 | 60.781 | 76.898 |
| Traits | Cluster One—Red (10) | Cluster Two—Green (5) | |||||
|---|---|---|---|---|---|---|---|
| Min | Max | Mean | Min | Max | Mean | F-Value | |
| PODLGT | 19.20 | 22.20 | 20.66 a | 19.60 | 22.30 | 21.24 a | 1.20 ns |
| PDWDTH | 9.15 | 10.92 | 9.64 a | 8.30 | 10.51 | 9.63 a | 0.00 ns |
| NoSP | 8.13 | 11.63 | 10.18 b | 11.00 | 12.25 | 11.65 a | 8.30 * |
| DTFPM | 83.20 | 90.80 | 86.04 a | 75.50 | 88.50 | 82.08 b | 4.78 * |
| SL | 9.19 | 10.29 | 9.53 a | 8.92 | 9.39 | 9.16 b | 5.82 * |
| SW | 8.54 | 8.94 | 8.72 a | 8.31 | 8.60 | 8.44 b | 16.65 ** |
| STH | 7.18 | 7.85 | 7.42 a | 6.93 | 7.70 | 7.14 b | 6.44 * |
| FW | 203.00 | 380.00 | 280.40 a | 185.00 | 546.00 | 316.50 a | 0.51 ns |
| PWT | 108.00 | 247.00 | 172.20 a | 170.00 | 228.00 | 198.00 a | 1.98 ns |
| SWPPL | 78.50 | 138.90 | 104.64 a | 70.40 | 109.40 | 84.77 a | 4.31 ns |
| Locus No | Allele Frequency | No of Alleles | Gene Diversity | PIC † |
|---|---|---|---|---|
| SSR-24 | 0.9400 | 3 | 0.1144 | 0.1109 |
| SSR-704 | 0.9333 | 3 | 0.1263 | 0.1218 |
| SSR-747 | 0.9400 | 3 | 0.1144 | 0.1109 |
| SSR-854 | 0.9333 | 3 | 0.1263 | 0.1218 |
| SSR-860 | 0.9200 | 4 | 0.1508 | 0.1462 |
| SSR-879 | 0.6267 | 6 | 0.5041 | 0.4229 |
| SSR-1104 | 0.9267 | 4 | 0.1387 | 0.1342 |
| SSR-3111 | 0.5800 | 6 | 0.5419 | 0.4596 |
| SSR-5819 | 0.9533 | 5 | 0.0903 | 0.0888 |
| SSR-11100 | 0.5400 | 5 | 0.5522 | 0.4606 |
| Mean | 0.8293 | 4.2 | 0.2459 | 0.2178 |
| Variation | df | SS | MS | Est. Var. | % |
|---|---|---|---|---|---|
| Among Accession | 2 | 1.733 | 0.867 | 0.013 | 5% |
| Within Accession | 147 | 33.180 | 0.226 | 0.226 | 95% |
| Total | 149 | 34.913 | 0.239 | 100% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shonde, T.E.O.; Adebayo, M.A.; Bhadmus, A.A.; Adejumobi, I.I.; Oyatomi, O.A.; Faloye, B.; Abberton, M.T. Diversity Assessment of Winged Bean [Psophocarpus tetragonolobus (L.) DC.] Accessions from IITA Genebank. Agronomy 2023, 13, 2150. https://doi.org/10.3390/agronomy13082150
Shonde TEO, Adebayo MA, Bhadmus AA, Adejumobi II, Oyatomi OA, Faloye B, Abberton MT. Diversity Assessment of Winged Bean [Psophocarpus tetragonolobus (L.) DC.] Accessions from IITA Genebank. Agronomy. 2023; 13(8):2150. https://doi.org/10.3390/agronomy13082150
Chicago/Turabian StyleShonde, Temitope Esther Olatunde, Moses Adeolu Adebayo, Ayomide Ajoke Bhadmus, Idris Ishola Adejumobi, Olaniyi A. Oyatomi, Benjamin Faloye, and Michael T. Abberton. 2023. "Diversity Assessment of Winged Bean [Psophocarpus tetragonolobus (L.) DC.] Accessions from IITA Genebank" Agronomy 13, no. 8: 2150. https://doi.org/10.3390/agronomy13082150
APA StyleShonde, T. E. O., Adebayo, M. A., Bhadmus, A. A., Adejumobi, I. I., Oyatomi, O. A., Faloye, B., & Abberton, M. T. (2023). Diversity Assessment of Winged Bean [Psophocarpus tetragonolobus (L.) DC.] Accessions from IITA Genebank. Agronomy, 13(8), 2150. https://doi.org/10.3390/agronomy13082150