

Assaying the Use of a Leonardite-Suspension Concentrate-Based Product as a Potential Biostimulant to Enhance Growth, NPK Use Efficiency, and Antioxidant Capacity in Lactuca sativa L.
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Experimental Design and Biostimulant Application
2.3. Lettuce Sampling
2.4. Determination of NPK Concentration and Use Efficiency
2.5. Antioxidant Quality Analysis
2.6. Statistical Procedures
3. Results
3.1. Shoot Growth
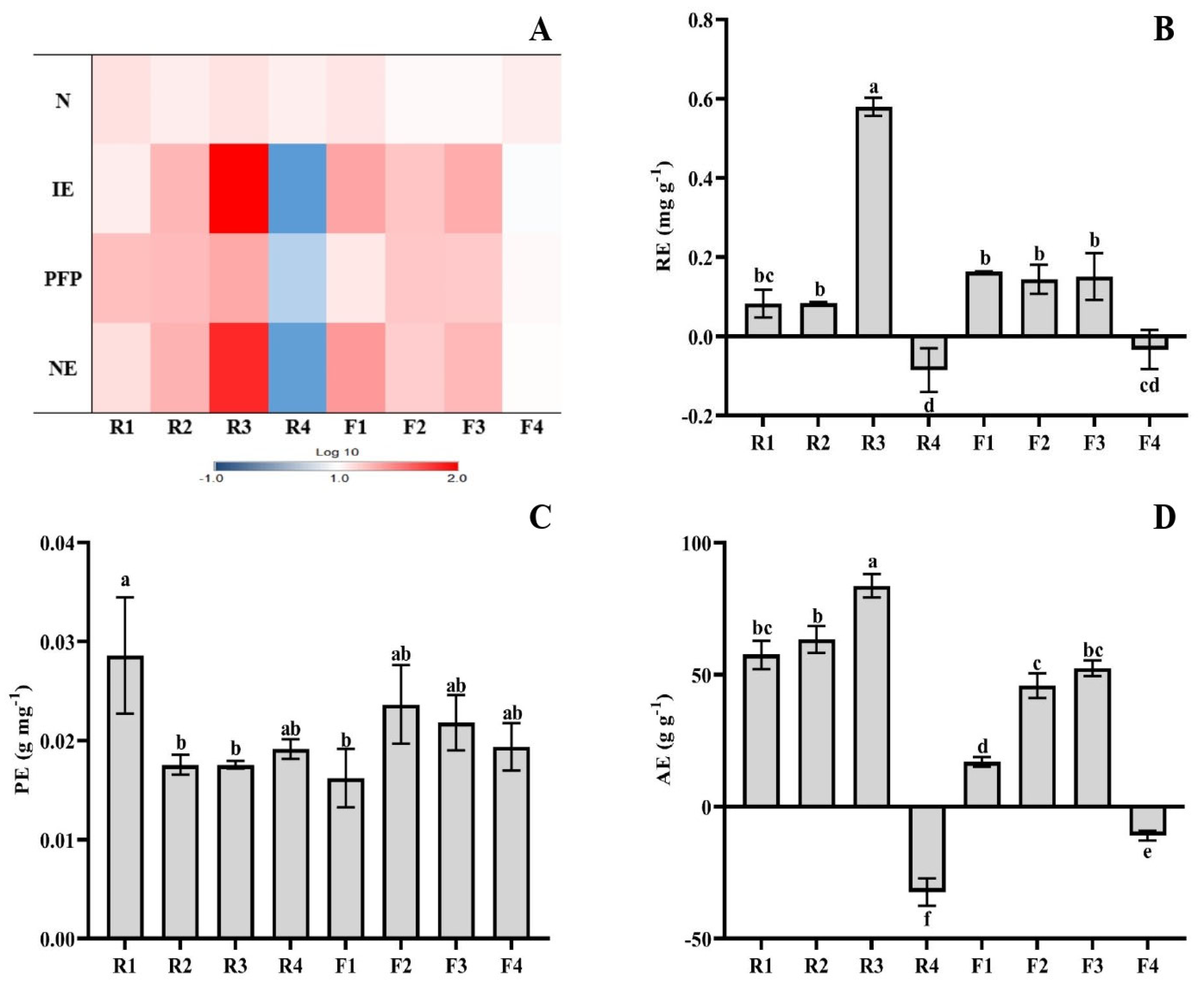
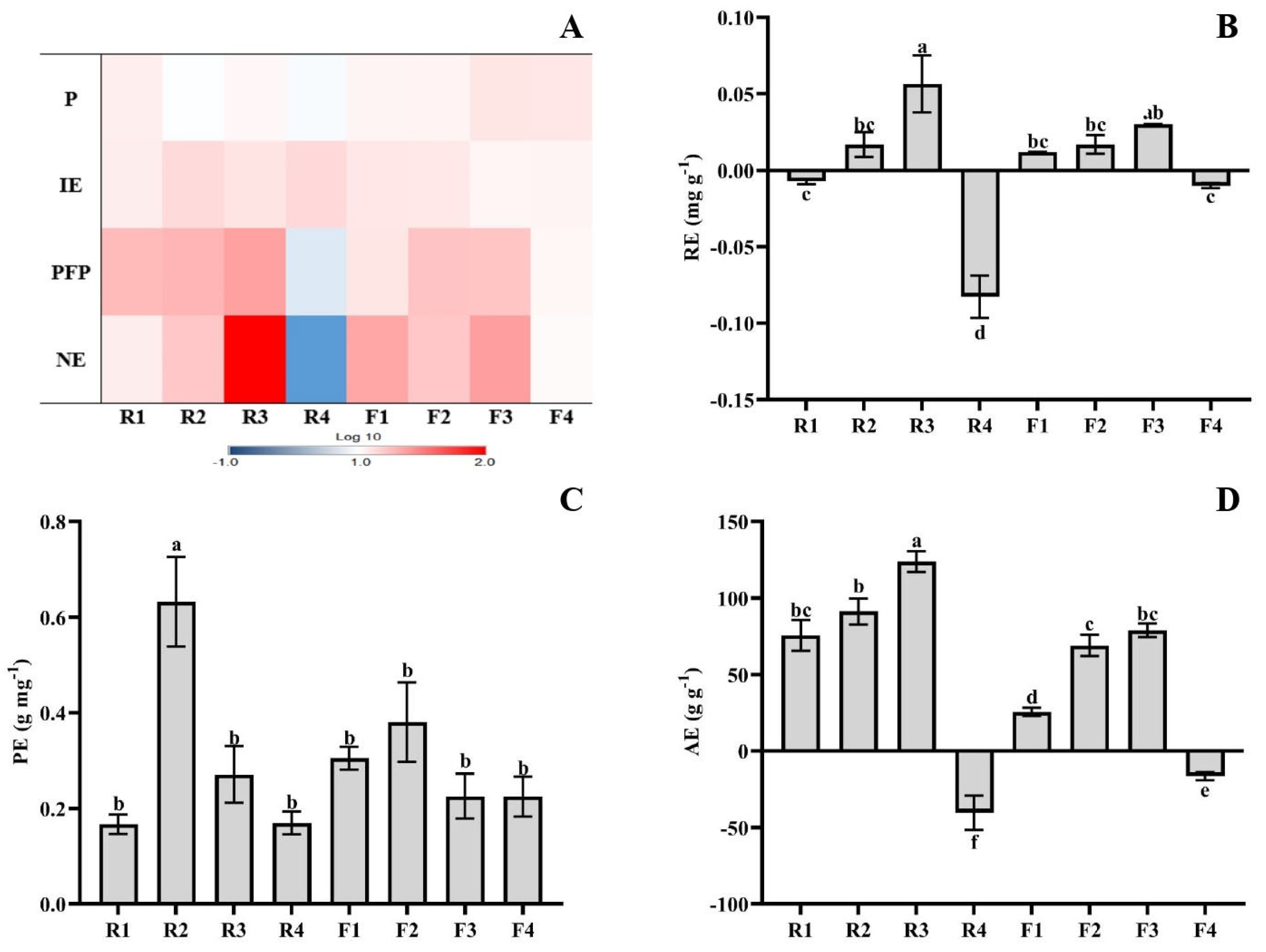
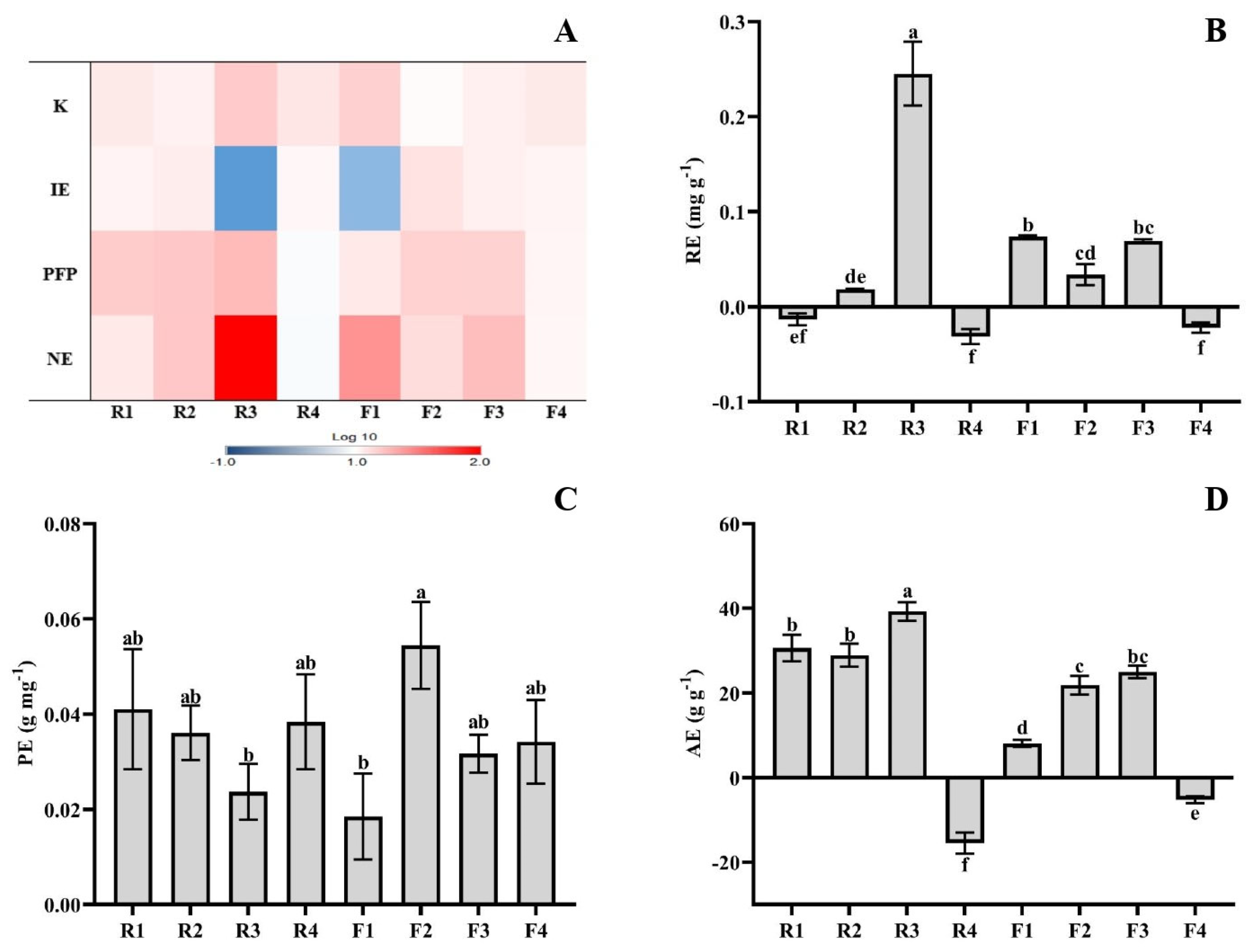
3.2. NPK Use Efficiency
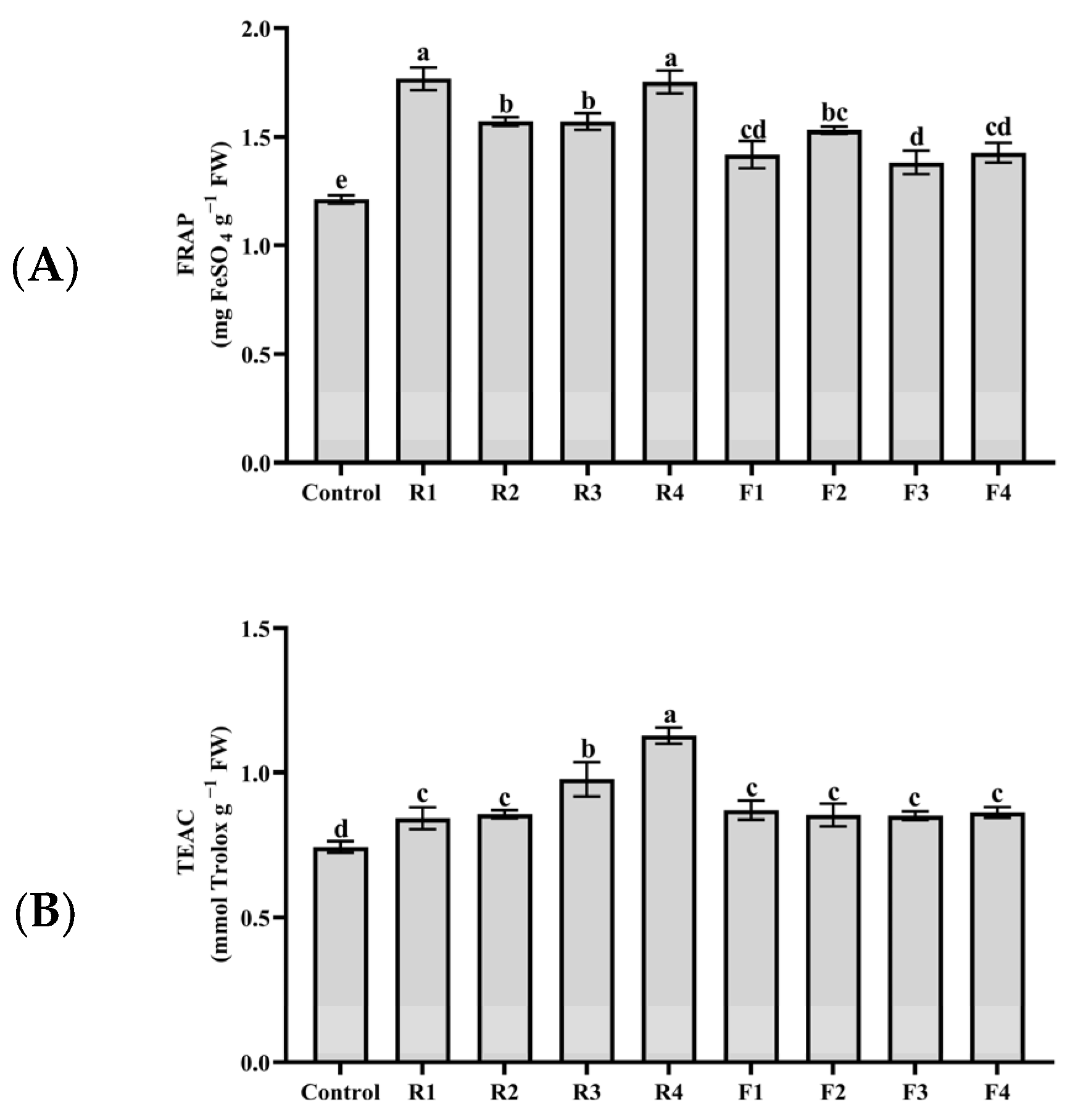
3.3. Antioxidant Capacity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tahery, S.; Munroe, P.; Marjo, C.E.; Rawal, A.; Horvat, J.; Mohammed, M.; Webber, J.B.W.; Arns, J.Y.; Arns, C.H.; Pan, G.; et al. A comparison between the characteristics of a biochar-NPK granule and a commercial NPK granule for application in the soil. Sci. Total Environ. 2022, 832, 155021. [Google Scholar] [CrossRef] [PubMed]
- Anas, M.; Liao, F.; Verma, K.K.; Sarwar, M.A.; Mahmood, A.; Chen, Z.L.; Li, Q.; Zeng, X.P.; Liu, Y.; Li, Y.R. Fate of nitrogen in agriculture and environment: Agronomic, eco-physiological and molecular approaches to improve nitrogen use efficiency. Biol. Res. 2020, 53, 47. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, Y.F.; Wu, W.H. Potassium and phosphorus transport and signaling in plants. J. Integr. Plant Biol. 2021, 63, 34–52. [Google Scholar] [CrossRef] [PubMed]
- Abdo, A.I.; El-Sobky, E.S.E.A.; Zhang, J. Optimizing maize yields using growth stimulants under the strategy of replacing chemicals with biological fertilizers. Front. Plant Sci. 2022, 13, 1069624. [Google Scholar] [CrossRef] [PubMed]
- Agbede, T.M. Effect of tillage, biochar, poultry manure and NPK 15-15-15 fertilizer, and their mixture on soil properties, growth and carrot (Daucus carota L.) yield under tropical conditions. Heliyon 2021, 7, e07391. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Elshayb, O.M.; Nada, A.M.; El-Leithy, S.A.; Baz, L.; Alhammad, B.A.; Mahdi, A.H.A. Azolla compost as an approach for enhancing growth, productivity and nutrient uptake of Oryza sativa L. Agronomy 2022, 12, 416. [Google Scholar] [CrossRef]
- Xu, D.P.; Li, Y.; Meng, X.; Zhou, T.; Zhou, Y.; Zheng, J.; Zhang, J.J.; Li, H. Bin Natural antioxidants in foods and medicinal plants: Extraction, assessment and resources. Int. J. Mol. Sci. 2017, 18, 96. [Google Scholar] [CrossRef]
- Michalak, M. Plant-derived antioxidants: Significance in skin health and the ageing process. Int. J. Mol. Sci. 2022, 23, 585. [Google Scholar] [CrossRef]
- Celi, G.E.A.; Gratão, P.L.; Lanza, M.G.D.B.; Dos Reis, A.R. Physiological and biochemical roles of ascorbic acid on mitigation of abiotic stresses in plants. Plant Physiol. Biochem. 2023, 202, 107970. [Google Scholar] [CrossRef]
- Ito, T.; Ohkama-Ohtsu, N. Degradation of glutathione and glutathione conjugates in plants. J. Exp. Bot. 2023, 74, 3313–3327. [Google Scholar] [CrossRef]
- Li, Z.; Ahammed, G.J. Hormonal regulation of anthocyanin biosynthesis for improved stress tolerance in plants. Plant Physiol. Biochem. 2023, 201, 107835. [Google Scholar] [CrossRef] [PubMed]
- Antonio, M.; Ortega-regules, A.E.; De Parrodi, C.A.; Lozada-Ram, D. Encapsulation of carotenoids—A review. Plants 2023, 12, 1–22. [Google Scholar]
- Khutami, C.; Sumiwi, S.A.; Khairul Ikram, N.K.; Muchtaridi, M. The effects of antioxidants from natural products on obesity, dyslipidemia, diabetes and their molecular signaling mechanism. Int. J. Mol. Sci. 2022, 23, 2056. [Google Scholar] [CrossRef] [PubMed]
- Mannino, G.; Campobenedetto, C.; Vigliante, I.; Contartese, V.; Gentile, C.; Bertea, C.M. The application of a plant biostimulant based on seaweed and yeast extract improved tomato fruit development and quality. Biomolecules 2020, 10, 1662. [Google Scholar] [CrossRef] [PubMed]
- Velasco, P.; Rodríguez, V.M.; Soengas, P.; Poveda, J. Trichoderma hamatum increases productivity, glucosinolate content and antioxidant potential of different leafy Brassica vegetables. Plants 2021, 10, 20449. [Google Scholar] [CrossRef] [PubMed]
- Cristofano, F.; El-Nakhel, C.; Pannico, A.; Giordano, M.; Colla, G.; Rouphael, Y. Foliar and root applications of vegetal-derived protein hydrolysates differentially enhance the yield and qualitative attributes of two lettuce cultivars grown in floating system. Agronomy 2021, 11, 1194. [Google Scholar] [CrossRef]
- Yang, F.; Tang, C.; Antonietti, M. Natural and artificial humic substances to manage minerals, ions, water, and soil microorganisms. Chem. Soc. Rev. 2021, 50, 6221–6239. [Google Scholar] [CrossRef]
- Ore, O.T.; Adeola, A.O.; Fapohunda, O.; Adedipe, D.T.; Bayode, A.A.; Adebiyi, F.M. Humic substances derived from unconventional resources: Extraction, properties, environmental impacts, and prospects. Environ. Sci. Pollut. Res. 2023, 30, 59106–59127. [Google Scholar] [CrossRef]
- Jannin, L.; Arkoun, M.; Ourry, A.; Laîné, P.; Goux, D.; Garnica, M.; Fuentes, M.; San Francisco, S.; Baigorri, R.; Cruz, F.; et al. Microarray analysis of humic acid effects on Brassica napus growth: Involvement of N, C and S metabolisms. Plant Soil 2012, 359, 297–319. [Google Scholar] [CrossRef]
- Hatami, E.; Shokouhian, A.A.; Ghanbari, A.R.; Naseri, L.A. Alleviating salt stress in almond rootstocks using of humic acid. Sci. Hortic. 2018, 237, 296–302. [Google Scholar] [CrossRef]
- Matuszak-Slamani, R.; Bejger, R.; Włodarczyk, M.; Kulpa, D.; Sienkiewicz, M.; Gołębiowska, D.; Skórska, E.; Ukalska-Jaruga, A. Effect of humic acids on soybean seedling growth under polyethylene-glycol-6000-induced drought stress. Agronomy 2022, 12, 1109. [Google Scholar] [CrossRef]
- Canellas, L.P.; Olivares, F.L.; Aguiar, N.O.; Jones, D.L.; Nebbioso, A.; Mazzei, P.; Piccolo, A. Humic and fulvic acids as biostimulants in horticulture. Sci. Hort. 2015, 196, 15–27. [Google Scholar] [CrossRef]
- Billard, V.; Etienne, P.; Jannin, L.; Garnica, M.; Cruz, F.; Garcia-Mina, J.M.; Yvin, J.C.; Ourry, A. Two biostimulants derived from algae or humic acid induce similar responses in the mineral content and gene expression of winter oilseed rape (Brassica napus L.). J. Plant Growth Regul. 2014, 33, 305–316. [Google Scholar] [CrossRef]
- Jindo, K.; Soares, T.S.; Peres, L.E.P.; Azevedo, I.G.; Aguiar, N.O.; Mazzei, P.; Spaccini, R.; Piccolo, A.; Olivares, F.L.; Canellas, L.P. Phosphorus speciation and high-affinity transporters are influenced by humic substances. J. Plant Nutr. Soil Sci. 2016, 179, 206–214. [Google Scholar] [CrossRef]
- Hernandez, O.L.; Calderín, A.; Huelva, R.; Martínez-Balmori, D.; Guridi, F.; Aguiar, N.O.; Olivares, F.L.; Canellas, L.P. Humic substances from vermicompost enhance urban lettuce production. Agron. Sustain. Dev. 2015, 35, 225–232. [Google Scholar] [CrossRef]
- Schiavon, M.; Pizzeghello, D.; Muscolo, A.; Vaccaro, S.; Francioso, O.; Nardi, S. High molecular size humic substances enhance phenylpropanoid metabolism in Maize (Zea mays L.). J. Chem. Ecol. 2010, 36, 662–669. [Google Scholar] [CrossRef] [PubMed]
- Monda, H.; McKenna, A.M.; Fountain, R.; Lamar, R.T. Bioactivity of humic acids extracted from shale ore: Molecular characterization and structure-activity relationship with tomato plant yield under nutritional stress. Front. Plant Sci. 2021, 12, 660224. [Google Scholar] [CrossRef] [PubMed]
- Muscolo, A.; Pizzeghello, D.; Francioso, O.; Cortes, S.S.; Nardi, S. Effectiveness of humic substances and phenolic compounds in regulating plant-biological functionality. Agronomy 2020, 10, 1553. [Google Scholar] [CrossRef]
- Peng, H.; Simko, I. Extending lettuce shelf life through integrated technologies. Curr. Opin. Biotechnol. 2023, 81, 102951. [Google Scholar] [CrossRef]
- Farooq, A.; Khattak, A.M.; Gul, G.; Habib, W.; Ahmad, S.; Asghar, M.; Rashid, T. Effect of moringa leaf extract on the performance of lettuce cultivars. Gesunde Pflanz 2023, 75, 1449–1459. [Google Scholar] [CrossRef]
- Paniagua-López, M.; Aguilar-Garrido, A.; Contero-Hurtado, J.; García-Romera, I.; Sierra-Aragón, M.; Romero-Freire, A. Ecotoxicological assessment of polluted soils one year after the application of different soil remediation rechniques. Toxics 2023, 11, 298. [Google Scholar] [CrossRef] [PubMed]
- Wolf, B. A comprehensive system of leaf analyses and its use for diagnosing crop nutrient status. Commun. Soil Sci. Plant Anal. 1982, 13, 1035–1059. [Google Scholar] [CrossRef]
- Dobermann, A. Nutrient use efficiency—Measurement and management. In Proceedings of the International Fertilizer Industry Association, Brussels, Belgium, 7–9 March 2007; pp. 1–22. [Google Scholar]
- Buturi, C.V.; Sabatino, L.; Mauro, R.P.; Navarro-León, E.; Blasco, B.; Leonardi, C.; Giuffrida, F. Iron biofortification of greenhouse soilless lettuce: An effective agronomic tool to improve the dietary mineral intake. Agronomy 2022, 12, 1793. [Google Scholar] [CrossRef]
- López-Moreno, F.J.; Atero-Calvo, S.; Navarro-León, E.; Blasco, B.; Soriano, T.; Ruiz, J.M. Evaluation of physiological and quality parameters of green asparagus spears subjected to three treatments against the decline syndrome. Agronomy 2021, 11, 937. [Google Scholar] [CrossRef]
- Rivero, R.M.; Ruiz, J.M.; García, P.C.; López-Lefebre, L.R.; Sánchez, E.; Romero, L. Resistance to cold and heat stress: Accumulation of phenolic compounds in tomato and watermelon plants. Plant Sci. 2001, 160, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Flores, P.; Navarro, J.M.; Garrido, C.; Rubio, J.S.; Martínez, V. Influence of Ca2+, K+ and NO3− fertilisation on nutritional quality of pepper. J. Sci. Food Agric. 2004, 84, 569–574. [Google Scholar] [CrossRef]
- Bishopp, A.; Lynch, J.P. The hidden half of crop yields. Nat. Plants. 2015, 1, 15117. [Google Scholar] [CrossRef]
- Deolu-Ajayi, A.O.; van der Meer, I.M.; van der Werf, A.; Karlova, R. The power of seaweeds as plant biostimulants to boost crop production under abiotic stress. Plant Cell Environ. 2022, 45, 2537–2553. [Google Scholar] [CrossRef]
- du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Tan, W.K.; Goenadie, V.; Lee, H.W.; Liang, X.; Loh, C.S.; Ong, C.N.; Tan, H.T.W. Growth and glucosinolate profiles of a common Asian green leafy vegetable, Brassica rapa subsp. chinensis var. parachinensis (choy sum), under LED lighting. Sci. Hortic. 2020, 261, 108922. [Google Scholar] [CrossRef]
- Tahir, M.M.; Khurshid, M.; Khan, M.Z.; Abbasi, M.K.; Kazmi, M.H. Lignite-derived humic acid effect on growth of wheat plants in different soils. Pedosphere 2011, 21, 124–131. [Google Scholar] [CrossRef]
- Bayat, H.; Shafie, F.; Aminifard, M.H.; Daghighi, S. Comparative effects of humic and fulvic acids as biostimulants on growth, antioxidant activity and nutrient content of yarrow (Achillea millefolium L.). Sci. Hortic. 2021, 279, 109912. [Google Scholar] [CrossRef]
- Nardi, S.; Schiavon, M.; Francioso, O. Chemical structure and biological activity of humic substances define their role as plant growth promoters. Molecules 2021, 26, 2256. [Google Scholar] [CrossRef] [PubMed]
- Shah, Z.H.; Rehman, H.M.; Akhtar, T.; Alsamadany, H.; Hamooh, B.T.; Mujtaba, T.; Daur, I.; Al Zahrani, Y.; Alzahrani, H.A.S.; Ali, S.; et al. Humic substances: Determining potential molecular regulatory processes in plants. Front. Plant Sci. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Haghighi, M.; Kafi, M.; Fang, P. Photosynthetic activity and N metabolism of lettuce as affected by humic acid. Int. J. Veg. Sci. 2012, 18, 182–189. [Google Scholar] [CrossRef]
- Olaetxea, M.; Mora, V.; Bacaicoa, E.; Baigorri, R.; Garnica, M.; Fuentes, M.; Zamarreño, A.M.; Spíchal, L.; García-Mina, J.M. Root ABA and H+-ATPase are key players in the root and shoot growth-promoting action of humic acids. Plant Direct 2019, 3, e00175. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Tang, C.; Jin, Y.; Cheng, K.; Yang, F. Contribution of exogenous humic substances to phosphorus availability in soil-plant ecosystem: A review. Crit. Rev. Environ. Sci. Technol. 2023, 53, 1085–1102. [Google Scholar] [CrossRef]
- Gerke, J. Review article: The effect of humic substances on phosphate and iron acquisition by higher plants: Qualitative and quantitative aspects. J. Plant Nutr. Soil Sci. 2021, 184, 329–338. [Google Scholar] [CrossRef]
- Suh, H.Y.; Yoo, K.S.; Suh, S.G. Effect of foliar application of fulvic acid on plant growth and fruit quality of tomato (Lycopersicon esculentum L.). Hortic. Environ. Biotechnol. 2014, 55, 455–461. [Google Scholar] [CrossRef]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Jan, R.; Kang, S.M.; Kim, K.M.; Lee, I.J. Extending thermotolerance to tomato seedlings by inoculation with SA1 isolate of Bacillus cereus and comparison with exogenous humic acid application. PLoS ONE 2020, 15, e0232228. [Google Scholar] [CrossRef]
- Yazdani, B.; Nikbakht, A.; Etemadi, N. Physiological effects of different combinations of humic and fulvic acid on Gerbera. Commun. Soil Sci. Plant Anal. 2014, 45, 1357–1368. [Google Scholar] [CrossRef]
- Akladious, S.A.; Mohamed, H.I. Ameliorative effects of calcium nitrate and humic acid on the growth, yield component and biochemical attribute of pepper (Capsicum annuum L.) plants grown under salt stress. Sci. Hortic. 2018, 236, 244–250. [Google Scholar] [CrossRef]
- Leite, J.M.; Pitumpe Arachchige, P.S.; Ciampitti, I.A.; Hettiarachchi, G.M.; Maurmann, L.; Trivelin, P.C.O.; Prasad, P.V.V.; Sunoj, S.V.J. Co-addition of humic substances and humic acids with urea enhances foliar nitrogen use efficiency in sugarcane (Saccharum officinarum L.). Heliyon 2020, 6, e05100. [Google Scholar] [CrossRef]
- Congreves, K.A.; Otchere, O.; Ferland, D.; Farzadfar, S.; Williams, S.; Arcand, M.M. Nitrogen use efficiency definitions of today and tomorrow. Front. Plant Sci. 2021, 12, 637108. [Google Scholar] [CrossRef] [PubMed]
- The, S.V.; Snyder, R.; Tegeder, M. Targeting nitrogen metabolism and transport processes to improve plant nitrogen use efficiency. Front. Plant Sci. 2021, 11, 628366. [Google Scholar] [CrossRef] [PubMed]
- Tshewang, S.; Rengel, Z.; Siddique, K.H.M.; Solaiman, Z.M. Microbial consortium inoculant increases pasture grasses yield in low-phosphorus soil by influencing root morphology, rhizosphere carboxylate exudation and mycorrhizal colonisation. J. Sci. Food Agric. 2022, 102, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Farzadfar, S.; Knight, J.D.; Congreves, K.A. Soil organic nitrogen: An overlooked but potentially significant contribution to crop nutrition. Plant Soil 2021, 462, 7–23. [Google Scholar] [CrossRef] [PubMed]
- Selladurai, R.; Jyoti Purakayastha, T. Effect of humic acid multinutrient fertilizers on yield and nutrient use efficiency of potato. J. Plant. Nutr. 2016, 39, 949–956. [Google Scholar] [CrossRef]
- Niaz, A.; Yaseen, M.; Shakar, M.; Sultana, S.; Ehsan, M.; Nazarat, A. Maize production and nitrogen use efficiency in response to nitrogen application with and without humic acid. J. Anim. Plant Sci. 2016, 26, 1641–1651. [Google Scholar]
- Kong, B.; Wu, Q.; Li, Y.; Zhu, T.; Ming, Y.; Li, C.; Li, C.; Wang, F.; Jiao, S.; Shi, L.; et al. The application of humic acid urea improves nitrogen use efficiency and crop yield by reducing the nitrogen loss compared with urea. Agriculture 2022, 12, 1996. [Google Scholar] [CrossRef]
- Santoro, V.; Della Lucia, M.C.; Francioso, O.; Stevanato, P.; Bertoldo, G.; Borella, M.; Ferrari, E.; Zaccone, C.; Schiavon, M.; Pizzeghello, D.; et al. Phosphorus acquisition efficiency and transcriptomic changes in maize plants treated with two lignohumates. Plants 2023, 12, 3291. [Google Scholar] [CrossRef] [PubMed]
- Scharff, L.B.; Saltenis, V.L.R.; Jensen, P.E.; Baekelandt, A.; Burgess, A.J.; Burow, M.; Ceriotti, A.; Cohan, J.P.; Geu-Flores, F.; Halkier, B.A.; et al. Prospects to improve the nutritional quality of crops. Food Energy Secur. 2022, 11, e327. [Google Scholar] [CrossRef]
- Soares, J.C.; Santos, C.S.; Carvalho, S.M.P.; Pintado, M.M.; Vasconcelos, M.W. Preserving the nutritional quality of crop plants under a changing climate: Importance and strategies. Plant Soil 2019, 443, 1–26. [Google Scholar] [CrossRef]
- Noperi-Mosqueda, L.C.; López-Moreno, F.J.; Navarro-León, E.; Sánchez, E.; Blasco, B.; Moreno, D.A.; Soriano, T.; Ruiz, J.M. Effects of asparagus decline on nutrients and phenolic compounds, spear quality, and allelopathy. Sci. Hortic. 2020, 261, 109029. [Google Scholar] [CrossRef]
- Zahedifar, M.; Najafian, S. Evaluation of essential oil composition and antioxidant activity of yarrow as influenced by foliar application of humic substance-based products: Using multivariate exploratory method. Sci. Hortic. 2023, 308, 111557. [Google Scholar] [CrossRef]
- Savarese, C.; Cozzolino, V.; Verrillo, M.; Vinci, G.; De Martino, A.; Scopa, A.; Piccolo, A. Combination of humic biostimulants with a microbial inoculum improves lettuce productivity, nutrient uptake, and primary and secondary metabolism. Plant Soil 2022, 481, 285–314. [Google Scholar] [CrossRef]
- Conselvan, G.B.; Pizzeghello, D.; Francioso, O.; Di Foggia, M.; Nardi, S.; Carletti, P. Biostimulant activity of humic substances extracted from leonardites. Plant Soil 2017, 420, 119–134. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Shoot FW (g−1 plant) | Leaf Area (cm2) | Shoot DW (g−1 plant) | |
|---|---|---|---|
| Control | 49.65 ± 1.30 c | 479.38 ± 13.67 b | 2.23 ± 0.22 bcd |
| R1 | 57.35 ± 2.76 a | 529.96 ± 15.81 a | 2.16 ± 0.14 bcd |
| R2 | 58.14 ± 2.07 a | 526.89 ± 13.41 a | 2.65 ± 0.01 bc |
| R3 | 61.17 ± 1.90 a | 532.05 ± 12.38 a | 3.95 ± 0.48 a |
| R4 | 45.90 ± 2.19 c | 477.38 ± 12.15 b | 1.99 ± 0.15 d |
| F1 | 50.83 ± 1.80 bc | 483.72 ± 9.49 b | 2.78 ± 0.04 b |
| F2 | 56.07 ± 0.66 ab | 528.17 ± 13.08 a | 2.55 ± 0.12 bcd |
| F3 | 55.91 ± 2.39 ab | 528.90 ± 14.17 a | 2.72 ± 0.22 bc |
| F4 | 48.36 ± 1.35 c | 498.98 ± 9.72 ab | 2.12 ± 0.11 cd |
| p-value | *** | * | *** |
| LSD0.05 | 5.43 | 37.65 | 0.63 |
| Total AsA (µg gFW−1) | Total GSH (µg gFW−1) | Flavonoids (µg gFW−1) | Total Phenols (µg gFW−1) | Lycopene (µg gFW−1) | β-Carotene (µg gFW−1) | Anthocyanins (mg gFW−1) | |
|---|---|---|---|---|---|---|---|
| Control | 8.51 ± 0.23 d | 69.33 ± 1.31 d | 353.85 ± 5.57 e | 693.18 ± 7.84 f | 1.37 ± 0.01 cd | 1.24 ± 0.05 cd | 1.67 ± 0.03 de |
| R1 | 16.83 ± 2.40 bc | 105.29 ± 3.73 a | 709.11 ± 8.30 c | 855.09 ± 4.94 c | 1.63 ± 0.01 a | 0.47 ± 0.03 f | 1.55 ± 0.05 e |
| R2 | 15.96 ± 1.80 bc | 67.45 ± 1.63 d | 260.31 ± 9.86 g | 599.98 ± 1.70 g | 1.46 ± 0.02 b | 0.94 ± 0.08 e | 1.70 ± 0.06 d |
| R3 | 15.36 ± 0.54 c | 73.04 ± 4.57 cd | 299.46 ± 20.07 f | 613.26 ± 4.25 g | 1.59 ± 0.03 a | 1.46 ± 0.17 bc | 1.63 ± 0.06 de |
| R4 | 19.75 ± 1.34 ab | 115.03 ± 3.44 a | 398.52 ± 7.75 d | 716.58 ± 4.51 ef | 1.44 ± 0.02 bc | 1.24 ± 0.02 cd | 1.56 ± 0.03 de |
| F1 | 16.14 ± 0.45 bc | 64.54 ± 2.76 d | 705.83 ± 10.10 c | 750.33 ± 1.90 de | 1.38 ± 0.03 cd | 1.70 ± 0.09 a | 1.92 ± 0.07 c |
| F2 | 21.25 ± 1.08 a | 84.33 ± 9.12 bc | 731.88 ± 14.15 bc | 759.60 ± 4.17 d | 1.36 ± 0.02 d | 1.37 ± 0.04 bcd | 2.33 ± 0.03 a |
| F3 | 23.24 ± 0.96 a | 86.05 ± 3.69 b | 761.15 ± 6.94 b | 1084.48 ± 39.50 a | 1.36 ± 0.03 d | 1.51 ± 0.08 ab | 2.24 ± 0.06 ab |
| F4 | 15.29 ± 1.97 c | 68.59 ± 4.24 d | 818.65 ± 19.25 a | 997.24 ± 8.24 b | 1.33 ± 0.02 d | 1.16 ± 0.03 de | 2.12 ± 0.05 b |
| p-value | *** | *** | *** | *** | *** | *** | *** |
| LSD0.05 | 3.91 | 12.37 | 34.92 | 39.61 | 0.07 | 0.23 | 0.14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atero-Calvo, S.; Magro, F.; Masetti, G.; Navarro-León, E.; Rios, J.J.; Ruiz, J.M. Assaying the Use of a Leonardite-Suspension Concentrate-Based Product as a Potential Biostimulant to Enhance Growth, NPK Use Efficiency, and Antioxidant Capacity in Lactuca sativa L. Agronomy 2024, 14, 64. https://doi.org/10.3390/agronomy14010064
Atero-Calvo S, Magro F, Masetti G, Navarro-León E, Rios JJ, Ruiz JM. Assaying the Use of a Leonardite-Suspension Concentrate-Based Product as a Potential Biostimulant to Enhance Growth, NPK Use Efficiency, and Antioxidant Capacity in Lactuca sativa L. Agronomy. 2024; 14(1):64. https://doi.org/10.3390/agronomy14010064
Chicago/Turabian StyleAtero-Calvo, Santiago, Francesco Magro, Giacomo Masetti, Eloy Navarro-León, Juan Jose Rios, and Juan Manuel Ruiz. 2024. "Assaying the Use of a Leonardite-Suspension Concentrate-Based Product as a Potential Biostimulant to Enhance Growth, NPK Use Efficiency, and Antioxidant Capacity in Lactuca sativa L." Agronomy 14, no. 1: 64. https://doi.org/10.3390/agronomy14010064
APA StyleAtero-Calvo, S., Magro, F., Masetti, G., Navarro-León, E., Rios, J. J., & Ruiz, J. M. (2024). Assaying the Use of a Leonardite-Suspension Concentrate-Based Product as a Potential Biostimulant to Enhance Growth, NPK Use Efficiency, and Antioxidant Capacity in Lactuca sativa L. Agronomy, 14(1), 64. https://doi.org/10.3390/agronomy14010064