

Structural and Functional Analysis of the Bacterial Community in the Soil of Continuously Cultivated Lonicera japonica Thunb. and Screening Antagonistic Bacteria for Plant Pathogens
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of the Soil Collection
2.2. Soil Physicochemical Property Analysis
2.3. DNA Extraction, PCR Amplification and Sequencing
2.4. Identification of Potential Isolates
2.5. Validation of the Antagonistic Bacteria Function
2.6. Characterization of Antagonistic Bacteria
2.7. Greenhouse Pot Experiments
2.8. Data Analysis
3. Results
3.1. Physicochemical Properties of Honeysuckle Soil
3.2. Alpha Diversity of Honeysuckle Root Soil Bacterial Community
3.3. Composition and Structure of the Bacterial Community
3.4. Correlation between Soil Physicochemical Properties and Bacterial Community
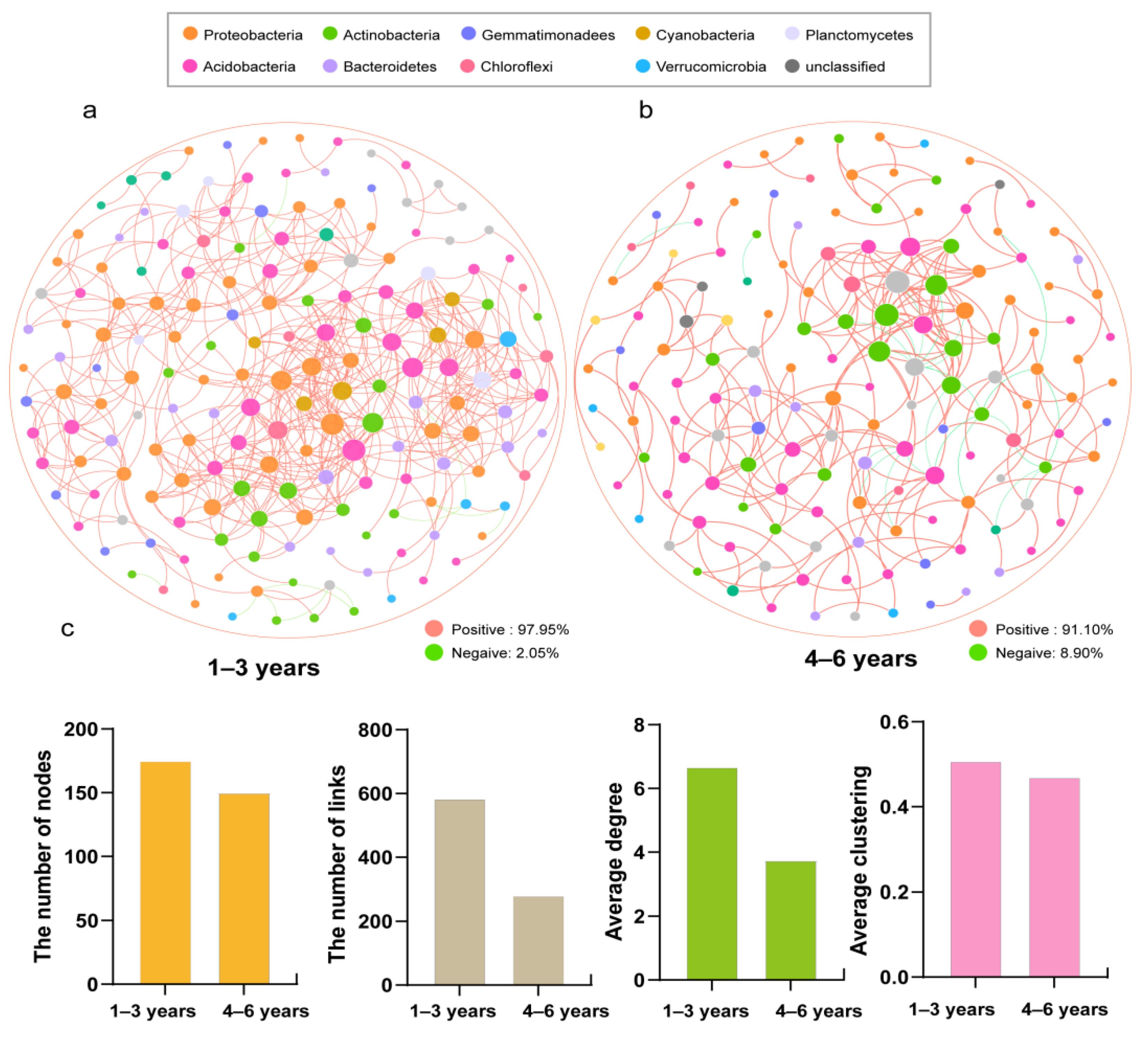
3.5. Network Analysis of the Bacterial Community
3.6. Functional Prediction Analysis of the Bacteria Community
3.7. Characteristics of the Antagonistic Bacteria
3.8. Biocontrol Potential of the Antagonistic Bacteria for Pathogens Fusarium oxysporum
4. Discussion
4.1. Continuous Cropping Led to Changes in Soil Physical and Chemical Properties
4.2. Continuous Cropping of Honeysuckle Led to Changes in Diversity of Bacterial Community
4.3. Continuous Cropping of Honeysuckle Led to Changes in Soil Bacteria Community Structure
4.4. Soil Physical and Chemical Properties Are Closely Related to Bacterial Community Structure
4.5. Continuous Cropping Reduced the Complexity of Bacterial Community Networks
4.6. Continuous Cropping Changed the Function of Bacteria Community
4.7. Antagonistic Bacteria Improve Honeysuckle Growth
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Shang, X.; Pan, H.; Li, M.; Miao, X.; Ding, H. Lonicera Japonica Thunb.: Ethnopharmacology, Phytochemistry and Pharmacology of an Important Traditional Chinese Medicine. J. Ethnopharmacol. 2011, 138, 1–21. [Google Scholar] [CrossRef]
- Zeng, H.; Li, Y.; Chen, J.; Wang, X.; Qiao, Z.; Li, Y.; Cai, N.; Liu, S. Lonicera Japonica ‘Fenglei’. HortScience Horts 2017, 52, 789–791. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, M.; He, L.; Li, Y. Chinese Herbs Combined with Western Medicine for Severe Acute Respiratory Syndrome (SARS). Cochrane Database Syst. Rev. 2012, 10, CD004882. [Google Scholar] [CrossRef]
- Ko, H.-C.; Wei, B.-L.; Chiou, W.-F. The Effect of Medicinal Plants Used in Chinese Folk Medicine on RANTES Secretion by Virus-Infected Human Epithelial Cells. J. Ethnopharmacol. 2006, 107, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Huang, Y.; Sun, M.; Ji, H.; Dou, H.; Hu, J.; Yan, Y.; Wang, X.; Chen, L. Honeysuckle-Encoded MicroRNA2911 Inhibits Enterovirus 71 Replication via Targeting VP1 Gene. Antivir. Res. 2018, 152, 117–123. [Google Scholar] [CrossRef]
- Chen, Y.; Du, J.; Li, Y.; Tang, H.; Yin, Z.; Yang, L.; Ding, X. Evolutions and Managements of Soil Microbial Community Structure Drove by Continuous Cropping. Front. Microbiol. 2022, 13, 839494. [Google Scholar] [CrossRef]
- Chen, P.; Wang, Y.; Liu, Q.; Zhang, Y.; Li, X.; Li, H.; Li, W. Phase Changes of Continuous Cropping Obstacles in Strawberry (Fragaria × Ananassa Duch.) Production. Appl. Soil. Ecol. 2020, 155, 103626. [Google Scholar] [CrossRef]
- Li, P.; Liu, J.; Saleem, M.; Li, G.; Luan, L.; Wu, M.; Li, Z. Reduced Chemodiversity Suppresses Rhizosphere Microbiome Functioning in the Mono-Cropped Agroecosystems. Microbiome 2022, 10, 108. [Google Scholar] [CrossRef]
- Zeeshan Ul Haq, M.; Yu, J.; Yao, G.; Yang, H.; Iqbal, H.A.; Tahir, H.; Cui, H.; Liu, Y.; Wu, Y. A Systematic Review on the Continuous Cropping Obstacles and Control Strategies in Medicinal Plants. Int. J. Mol. Sci. 2023, 24, 12470. [Google Scholar] [CrossRef]
- Hartmann, M.; Six, J. Soil Structure and Microbiome Functions in Agroecosystems. Nat. Rev. Earth Environ. 2023, 4, 4–18. [Google Scholar] [CrossRef]
- Carrión, V.J.; Perez-Jaramillo, J.; Cordovez, V.; Tracanna, V.; de Hollander, M.; Ruiz-Buck, D.; Mendes, L.W.; van Ijcken, W.F.J.; Gomez-Exposito, R.; Elsayed, S.S.; et al. Pathogen-Induced Activation of Disease-Suppressive Functions in the Endophytic Root Microbiome. Science 2019, 366, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, Y.; Wu, G.; Zhang, N.; Shen, Q.; Zhang, R. Beneficial Rhizobacterium Bacillus Amyloliquefaciens SQR9 Induces Plant Salt Tolerance through Spermidine Production. Mol. Plant. Microbe. Interact. 2017, 30, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Pervaiz, Z.H.; Iqbal, J.; Zhang, Q.; Chen, D.; Wei, H.; Saleem, M. Continuous Cropping Alters Multiple Biotic and Abiotic Indicators of Soil Health. Soil. Syst. 2020, 4, 59. [Google Scholar] [CrossRef]
- Wang, G.; Ren, Y.; Bai, X.; Su, Y.; Han, J. Contributions of Beneficial Microorganisms in Soil Remediation and Quality Improvement of Medicinal Plants. Plants 2022, 11, 3200. [Google Scholar] [CrossRef] [PubMed]
- Todorović, I.; Moënne-Loccoz, Y.; Raičević, V.; Jovičić-Petrović, J.; Muller, D. Microbial Diversity in Soils Suppressive to Fusarium Diseases. Front. Plant Sci. 2023, 14, 1228749. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Liu, D.; Liu, L.; Wang, Y.; Zhang, Y. Screening and Identification of Antagonistic Bacteria from Vermicompost against Fusarium Oxysporum f. Sp. Cucumerinum. Acta Agric. Scand. Sect. B Soil. Plant Sci. 2021, 71, 266–272. [Google Scholar] [CrossRef]
- Zhang, Q.; Sun, Z.; Wang, X.S. Occurrence pattern and pathogen identification of root rot of Lonicera japonica Thunb. J. Anhui Agric. Sci. 2019, 47, 129–131. (In Chinese) [Google Scholar]
- Liu, M.T.; Sun, H.T.; Zhang, D.F. A distinguish on a new root rot of Lonicera japonica and study on biological characteristic of the pathogen. Acta Agric. Boreali-Sin. 2004, 19, 109–111. (In Chinese) [Google Scholar]
- Zhang, S.; Jiang, Q.; Liu, X.; Liu, L.; Ding, W. Plant Growth Promoting Rhizobacteria Alleviate Aluminum Toxicity and Ginger Bacterial Wilt in Acidic Continuous Cropping Soil. Front. Microbiol. 2020, 11, 569512. [Google Scholar] [CrossRef]
- Hu, Y.; Li, Y.; Yang, X.; Li, C.; Wang, L.; Feng, J.; Chen, S.; Li, X.; Yang, Y. Effects of Integrated Biocontrol on Bacterial Wilt and Rhizosphere Bacterial Community of Tobacco. Sci. Rep. 2021, 11, 2653. [Google Scholar] [CrossRef]
- Niu, D.; Liu, H.; Jiang, C.; Wang, Y.; Wang, Q.; Jin, H.; Guo, J. The Plant Growth-Promoting Rhizobacterium Bacillus Cereus AR156 Induces Systemic Resistance in Arabidopsis Thaliana by Simultaneously Activating Salicylate- and Jasmonate/Ethylene-Dependent Signaling Pathways. Mol. Plant. Microbe. Interact. 2011, 24, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Guo, W.; Chen, X. Exogenous Addition of Alkanoic Acids Enhanced Production of Antifungal Lipopeptides in Bacillus Amyloliquefaciens Pc3. Appl. Microbiol. Biotechnol. 2019, 103, 5367–5377. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, Y.; Fu, X.; Li, Y.; Wang, Q. Isolation and Characterization of Bacillus Amyloliquefaciens PG12 for the Biological Control of Apple Ring Rot. Postharvest Biol. Technol. 2016, 115, 113–121. [Google Scholar] [CrossRef]
- Fira, D.; Dimkić, I.; Berić, T.; Lozo, J.; Stanković, S. Biological Control of Plant Pathogens by Bacillus Species. J. Biotechnol. 2018, 285, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Wang, S.; Diao, M.; Fu, J.; Borthwick, A.G.L. Microbial Community Responses to Vanadium Distributions in Mining Geological Environments and Bioremediation Assessment. J. Geophys. Res. Biogeosci. 2019, 124, 601–615. [Google Scholar] [CrossRef]
- Parkinson, J.A.; Allen, S.E. A Wet Oxidation Procedure Suitable for the Determination of Nitrogen and Mineral Nutrients in Biological Material. Commun. Soil. Sci. Plant Anal. 1975, 6, 1–11. [Google Scholar] [CrossRef]
- Walters, W.; Hyde, E.R.; Berg-Lyons, D.; Ackermann, G.; Humphrey, G.; Parada, A.; Gilbert, J.A.; Jansson, J.K.; Caporaso, J.G.; Fuhrman, J.A.; et al. Improved Bacterial 16S rRNA Gene (V4 and V4-5) and Fungal Internal Transcribed Spacer Marker Gene Primers for Microbial Community Surveys. mSystems 2016, 1, e00009-15. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly Accurate OTU Sequences from Microbial Amplicon Reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Ding, Y.; Wang, J.; Liu, Y.; Chen, S. Isolation and Identification of Nitrogen-Fixing Bacilli from Plant Rhizospheres in Beijing Region. J. Appl. Microbiol. 2005, 99, 1271–1281. [Google Scholar] [CrossRef]
- Gothwal, R.; Nigam, V.; Medicherla, K.; Sasmal, D.; Ghosh, P. Phosphate Solubilization by Rhizospheric Bacterial Isolates from Economically Important Desert Plants. Indian J. Microbiol. 2006, 46, 355–361. [Google Scholar]
- MAYER, A.M. Determination of Indole Acetic Acid by the Salkowsky Reaction. Nature 1958, 182, 1670–1671. [Google Scholar] [CrossRef]
- Kumar, A.; Kumar, A.; Devi, S.; Patil, S.; Payal, C.; Negi, S. Isolation, Screening and Characterization of Bacteria from Rhizospheric Soils for Different Plant Growth Promotion (PGP) Activities: An in Vitro Study. Recent Res. Sci. Technol. 2012, 4, 1–5. [Google Scholar]
- Chang, W.T.; Hsieh, C.H.; Hsieh, H.S.; Chen, C. Conversion of Crude Chitosan to an Anti-Fungal Protease by Bacillus Cereus. World J. Microbiol. Biotechnol. 2009, 25, 375–382. [Google Scholar] [CrossRef]
- Azizoglu, Z.B.; Yilmaz, S.; Azizoglu, U.; Karabörklü, S.; Temizgul, R.; Ayvaz, A. Molecular Characterization of the Chitinase Genes of Native Bacillus Thuringiensis Isolates and Their Antagonistic Activity against Three Important Phytopathogenic Fungi. Biologia 2021, 76, 2745–2755. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K. QIIME Allows Analysis of High-Throughput Community Sequencing Data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and Clustering Orders of Magnitude Faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Kemp, P.F.; Aller, J.Y. Bacterial Diversity in Aquatic and Other Environments: What 16S rDNA Libraries Can Tell Us. FEMS Microbiol. Ecol. 2004, 47, 161–177. [Google Scholar] [CrossRef] [PubMed]
- Stilianos, L.; Wegener, P.L.; Michael, D. Decoupling Function and Taxonomy in the Global Ocean Microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef]
- Bai, Y.; Wang, G.; Cheng, Y.; Shi, P.; Yang, C.; Yang, H.; Xu, Z. Soil Acidification in Continuously Cropped Tobacco Alters Bacterial Community Structure and Diversity via the Accumulation of Phenolic Acids. Sci. Rep. 2019, 9, 12499. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Fan, S.; Qin, J.; Dai, J.; Zhao, F.; Gao, L.; Lian, X.; Shang, W.; Xu, X.; Hu, X. Changes in the Microbiome in the Soil of an American Ginseng Continuous Plantation. Front. Plant Sci. 2020, 11, 572199. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Hu, Y.; Han, M.; Xu, J.; Wang, X.; Liu, L.; Tang, Z.; Jiao, W.; Jin, R.; Liu, M.; et al. Effects of Continuous Cropping of Sweet Potatoes on the Bacterial Community Structure in Rhizospheric Soil. BMC Microbiol. 2021, 21, 102. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, Q.; Zhou, J.; Wei, Q. Illumina Amplicon Sequencing of 16S rRNA Tag Reveals Bacterial Community Development in the Rhizosphere of Apple Nurseries at a Replant Disease Site and a New Planting Site. PLoS ONE 2014, 9, e111744. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Wu, J.; Ji, Q.; Wu, W.; Dong, S.; Yu, J.; Zhang, Q.; Qin, L. Diversity of Rhizosphere and Endophytic Fungi in Atractylodes Macrocephala during Continuous Cropping. PeerJ 2020, 8, e8905. [Google Scholar] [CrossRef]
- Yao, Y.; Yao, X.; An, L.; Bai, Y.; Xie, D.; Wu, K. Rhizosphere Bacterial Community Response to Continuous Cropping of Tibetan Barley. Front. Microbiol. 2020, 11, 551444. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wang, Z.; Niu, J.; Dang, K.; Zhang, S.; Wang, S.; Wang, Z. Changes in Physicochemical Properties, Enzymatic Activities, and the Microbial Community of Soil Significantly Influence the Continuous Cropping of Panax Quinquefolius L. (American Ginseng). Plant Soil 2021, 463, 427–446. [Google Scholar] [CrossRef]
- Wu, L.; Chen, J.; Xiao, Z.; Zhu, X.; Wang, J.; Wu, H.; Wu, Y.; Zhang, Z.; Lin, W. Barcoded Pyrosequencing Reveals a Shift in the Bacterial Community in the Rhizosphere and Rhizoplane of Rehmannia Glutinosa under Consecutive Monoculture. Int. J. Mol. Sci. 2018, 19, 850. [Google Scholar] [CrossRef] [PubMed]
- Lei, H.; Liu, A.; Hou, Q.; Zhao, Q.; Guo, J.; Wang, Z. Diversity Patterns of Soil Microbial Communities in the Sophora Flavescens Rhizosphere in Response to Continuous Monocropping. BMC Microbiol. 2020, 20, 272. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, D.; Yang, Y.; Pan, Y.; Zhao, D.; Zhu, J.; Zhang, L.; Yang, Z. Dissecting the Effect of Continuous Cropping of Potato on Soil Bacterial Communities as Revealed by High-Throughput Sequencing. PLoS ONE 2020, 15, e0233356. [Google Scholar] [CrossRef]
- Li, J.; Chen, X.; Li, S.; Zuo, Z.; Zhan, R.; He, R. Variations of Rhizospheric Soil Microbial Communities in Response to Continuous Andrographis Paniculata Cropping Practices. Bot. Stud. 2020, 61, 18. [Google Scholar] [CrossRef]
- Saleem, M.; Hu, J.; Jousset, A. More Than the Sum of Its Parts: Microbiome Biodiversity as a Driver of Plant Growth and Soil Health. Annu. Rev. Ecol. Evol. Syst. 2019, 50, 145–168. [Google Scholar] [CrossRef]
- Chaer, G.; Fernandes, M.; Myrold, D.; Bottomley, P. Comparative Resistance and Resilience of Soil Microbial Communities and Enzyme Activities in Adjacent Native Forest and Agricultural Soils. Microb. Ecol. 2009, 58, 414–424. [Google Scholar] [CrossRef]
- Cheng, Z.; Lei, S.; Li, Y.; Huang, W.; Ma, R.; Xiong, J.; Zhang, T.; Jin, L.; Haq, H.U.; Xu, X.; et al. Revealing the Variation and Stability of Bacterial Communities in Tomato Rhizosphere Microbiota. Microorganisms 2020, 8, 170. [Google Scholar] [CrossRef]
- Singh, J.S.; Kumar, A.; Rai, A.N.; Singh, D.P. Cyanobacteria: A Precious Bio-Resource in Agriculture, Ecosystem, and Environmental Sustainability. Front. Microbiol. 2016, 7, 529. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Li, H.; Wang, J.; Gao, W.; Shu, X.; Sun, X.; Wang, K.; Duan, Y.; Liu, Y.; Kuramae, E.E.; et al. Composition, Function and Succession of Bacterial Communities in the Tomato Rhizosphere during Continuous Cropping. Biol. Fertil. Soils 2023, 59, 723–732. [Google Scholar] [CrossRef]
- Lazcano, C.; Boyd, E.; Holmes, G.; Hewavitharana, S.; Pasulka, A.; Ivors, K. The Rhizosphere Microbiome Plays a Role in the Resistance to Soil-Borne Pathogens and Nutrient Uptake of Strawberry Cultivars under Field Conditions. Sci. Rep. 2021, 11, 3188. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Bai, J.L.; Yang, H.T.; Zhang, W.D.; Xiong, Y.W.; Ding, P.; Qin, S. Phylogenetic Diversity and Investigation of Plant Growth-Promoting Traits of Actinobacteria in Coastal Salt Marsh Plant Rhizospheres from Jiangsu, China. Syst. Appl. Microbiol. 2018, 41, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Daims, H.; Lücker, S.; Wagner, M. A New Perspective on Microbes Formerly Known as Nitrite-Oxidizing Bacteria. Trends Microbiol. 2016, 24, 699–712. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Xiao, Y.; Lv, F.; Hu, L.; Wei, L.; Yuan, Z.; Lin, H. Bacterial Community Structure and Functional Potential of Rhizosphere Soils as Influenced by Nitrogen Addition and Bacterial Wilt Disease under Continuous Sesame Cropping. Appl. Soil Ecol. 2018, 125, 117–127. [Google Scholar] [CrossRef]
- Wang, T.; Yang, K.; Ma, Q.; Jiang, X.; Zhou, Y.; Kong, D.; Wang, Z.; Parales, R.E.; Li, L.; Zhao, X.; et al. Rhizosphere Microbial Community Diversity and Function Analysis of Cut Chrysanthemum During Continuous Monocropping. Front. Microbiol. 2022, 13, 801546. [Google Scholar] [CrossRef]
- Huang, W.; Sun, D.; Fu, J.; Zhao, H.; Wang, R.; An, Y. Effects of Continuous Sugar Beet Cropping on Rhizospheric Microbial Communities. Genes 2019, 11, 13. [Google Scholar] [CrossRef]
- Matsumoto, H.; Fan, X.; Wang, Y.; Kusstatscher, P.; Duan, J.; Wu, S.; Chen, S.; Qiao, K.; Wang, Y.; Ma, B.; et al. Bacterial Seed Endophyte Shapes Disease Resistance in Rice. Nat. Plants 2021, 7, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Hu, Y.; Zhang, S.; Noll, L.; Böckle, T.; Dietrich, M.; Herbold, C.W.; Eichorst, S.A.; Woebken, D.; Richter, A.; et al. Soil Multifunctionality Is Affected by the Soil Environment and by Microbial Community Composition and Diversity. Soil Biol. Biochem. 2019, 136, 107521. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Zhou, D.; Wei, H.; Wu, S.; Xie, B. Alleviating Soil Degradation Caused by Watermelon Continuous Cropping Obstacle: Application of Urban Waste Compost. Chemosphere 2021, 262, 128387. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Hu, Y.; Razavi, B.S.; Zhou, J.; Shen, J.; Nannipieri, P.; Wu, J.; Ge, T. Rare Taxa of Alkaline Phosphomonoesterase-Harboring Microorganisms Mediate Soil Phosphorus Mineralization. Soil Biol. Biochem. 2019, 131, 62–70. [Google Scholar] [CrossRef]
- Hontoria, C.; García-González, I.; Quemada, M.; Roldán, A.; Alguacil, M.M. The Cover Crop Determines the AMF Community Composition in Soil and in Roots of Maize after a Ten-Year Continuous Crop Rotation. Sci. Total Environ. 2019, 660, 913–922. [Google Scholar] [CrossRef]
- Wan, W.; Tan, J.; Wang, Y.; Qin, Y.; He, H.; Wu, H.; Zuo, W.; He, D. Responses of the Rhizosphere Bacterial Community in Acidic Crop Soil to pH: Changes in Diversity, Composition, Interaction, and Function. Sci. Total Environ. 2020, 700, 134418. [Google Scholar] [CrossRef]
- Zhu, S.; Wang, Y.; Xu, X.; Liu, T.; Wu, D.; Zheng, X.; Tang, S.; Dai, Q. Potential Use of High-Throughput Sequencing of Soil Microbial Communities for Estimating the Adverse Effects of Continuous Cropping on Ramie (Boehmeria Nivea L. Gaud). PLoS ONE 2018, 13, e0197095. [Google Scholar] [CrossRef]
- Santolini, M.; Barabási, A.L. Predicting Perturbation Patterns from the Topology of Biological Networks. Proc. Natl. Acad. Sci. USA 2018, 115, E6375–E6383. [Google Scholar] [CrossRef]
- Banerjee, S.; Walder, F.; Büchi, L.; Meyer, M.; Held, A.Y.; Gattinger, A.; Keller, T.; Charles, R.; van der Heijden, M.G.A. Agricultural Intensification Reduces Microbial Network Complexity and the Abundance of Keystone Taxa in Roots. ISME J. 2019, 13, 1722–1736. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Schlaeppi, K.; van der Heijden, M.G.A. Keystone Taxa as Drivers of Microbiome Structure and Functioning. Nat. Rev. Microbiol. 2018, 16, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Mendes, R.; Pizzirani-Kleiner, A.A.; Araujo, W.L.; Raaijmakers, J.M. Diversity of Cultivated Endophytic Bacteria from Sugarcane: Genetic and Biochemical Characterization of Burkholderia Cepacia Complex Isolates. Appl. Environ. Microbiol. 2007, 73, 7259–7267. [Google Scholar] [CrossRef]
- Meng, L.; Xu, C.; Wu, F. Huhe Microbial Co-Occurrence Networks Driven by Low-Abundance Microbial Taxa during Composting Dominate Lignocellulose Degradation. Sci. Total Environ. 2022, 845, 157197. [Google Scholar] [CrossRef]
- Chen, X.; Wong, J.T.F.; Chen, Z.; Tang, T.W.L.; Guo, H.; Leung, A.O.W.; Ng, C.W.W.; Wong, M.H. Effects of Biochar on the Ecological Performance of a Subtropical Landfill. Sci. Total Environ. 2018, 644, 963–975. [Google Scholar] [CrossRef]
- Li, Q.; Gao, Y.; Yang, A. Sulfur Homeostasis in Plants. Int. J. Mol. Sci. 2020, 21, 8926. [Google Scholar] [CrossRef]
- Liang, Z.; Yu, Y.; Ye, Z.; Li, G.; Wang, W.; An, T. Pollution Profiles of Antibiotic Resistance Genes Associated with Airborne Opportunistic Pathogens from Typical Area, Pearl River Estuary and Their Exposure Risk to Human. Environ. Int. 2020, 143, 105934. [Google Scholar] [CrossRef] [PubMed]
- Mendes, R.; Kruijt, M.; de Bruijn, I.; Dekkers, E.; van der Voort, M.; Schneider, J.H.M.; Piceno, Y.M.; DeSantis, T.Z.; Andersen, G.L.; Bakker, P.A.H.M.; et al. Deciphering the Rhizosphere Microbiome for Disease-Suppressive Bacteria. Science 2011, 332, 1097–1100. [Google Scholar] [CrossRef]
- Rana, A.; Sahgal, M.; Johri, B.N. Fusarium oxysporum: Genomics, Diversity and Plant–Host Interaction BT. In Developments in Fungal Biology and Applied Mycology; Satyanarayana, T., Deshmukh, S.K., Johri, B.N., Eds.; Springer: Singapore, 2017; pp. 159–199. ISBN 978-981-10-4768-8. [Google Scholar]
- Kashiwa, T.; Kozaki, T.; Ishii, K.; Turgeon, B.G.; Teraoka, T.; Komatsu, K.; Arie, T. Sequencing of Individual Chromosomes of Plant Pathogenic Fusarium oxysporum. Fungal Genet. Biol. 2017, 98, 46–51. [Google Scholar] [CrossRef]
- Rajninec, M.; Fratrikova, M.; Boszoradova, E.; Jopcik, M.; Bauer, M.; Libantova, J. Basic β-1,3-Glucanase from Drosera Binata Exhibits Antifungal Potential in Transgenic Tobacco Plants. Plants 2021, 10, 1747. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, M.; Torres, M.; Blanco, L.; Béjar, V.; Sampedro, I.; Llamas, I. Plant Growth-Promoting Activity and Quorum Quenching-Mediated Biocontrol of Bacterial Phytopathogens by Pseudomonas Segetis Strain P6. Sci. Rep. 2020, 10, 4121. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Xu, Z.; Xie, J.; Hesselberg-Thomsen, V.; Tan, T.; Zheng, D.; Strube, M.L.; Dragoš, A.; Shen, Q.; Zhang, R.; et al. Bacillus Velezensis Stimulates Resident Rhizosphere Pseudomonas Stutzeri for Plant Health through Metabolic Interactions. ISME J. 2022, 16, 774–787. [Google Scholar] [CrossRef]
- Shwaiman, H.A.; Shahid, M.; Elgorban, A.M.; Siddique, K.H.M.; Syed, A. Beijerinckia Fluminensis BFC-33, a Novel Multi-Stress-Tolerant Soil Bacterium: Deciphering the Stress Amelioration, Phytopathogenic Inhibition and Growth Promotion in Triticum Aestivum (L.). Chemosphere 2022, 295, 133843. [Google Scholar] [CrossRef]
- Frey-Klett, P.; Burlinson, P.; Deveau, A.; Barret, M.; Tarkka, M.; Sarniguet, A. Bacterial-Fungal Interactions: Hyphens between Agricultural, Clinical, Environmental, and Food Microbiologists. Microbiol. Mol. Biol. Rev. 2011, 75, 583–609. [Google Scholar] [CrossRef]
- Berg, G. Plant–Microbe Interactions Promoting Plant Growth and Health: Perspectives for Controlled Use of Microorganisms in Agriculture. Appl. Microbiol. Biotechnol. 2009, 84, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Santhanam, R.; Luu, V.T.; Weinhold, A.; Goldberg, J.; Oh, Y.; Baldwin, I.T. Native Root-Associated Bacteria Rescue a Plant from a Sudden-Wilt Disease That Emerged during Continuous Cropping. Proc. Natl. Acad. Sci. USA 2015, 112, E5013–E5020. [Google Scholar] [CrossRef]
- Shen, Z.; Xue, C.; Taylor, P.W.J.; Ou, Y.; Wang, B.; Zhao, Y.; Ruan, Y.; Li, R.; Shen, Q. Soil Pre-Fumigation Could Effectively Improve the Disease Suppressiveness of Biofertilizer to Banana Fusarium Wilt Disease by Reshaping the Soil Microbiome. Biol. Fertil. Soils 2018, 54, 793–806. [Google Scholar] [CrossRef]
- Xun, W.; Shao, J.; Shen, Q.; Zhang, R. Rhizosphere Microbiome: Functional Compensatory Assembly for Plant Fitness. Comput. Struct. Biotechnol. J. 2021, 19, 5487–5493. [Google Scholar] [CrossRef] [PubMed]
- Xun, W.; Ren, Y.; Yan, H.; Ma, A.; Liu, Z.; Wang, L.; Zhang, N.; Xu, Z.; Miao, Y.; Feng, H.; et al. Sustained Inhibition of Maize Seed-Borne Fusarium Using a Bacillus-Dominated Rhizospheric Stable Core Microbiota with Unique Cooperative Patterns. Adv. Sci. 2023, 10, 2205215. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Yuan, X.; Yang, J.; Yang, Y.; Jv, H.; Li, R.; Jia, Z.; Ruan, Y. Selection of Rhizosphere Communities of Diverse Rotation Crops Reveals Unique Core Microbiome Associated with Reduced Banana Fusarium Wilt Disease. New Phytol. 2023, 238, 2194–2209. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Yield (kg/acre) | pH | TN (g/kg) | TP (g/kg) | TK (g/kg) |
|---|---|---|---|---|---|
| CK | - | 6.75 ± 0.23 a | 0.69 ± 0.03 a | 0.80 ± 0.04 a | 17.56 ± 3.21 a |
| HS1 | 102.69 ± 2.34 c | 6.71 ± 0.15 a | 0.65 ± 0.10 a | 0.71 ± 0.06 a | 19.76 ± 2.37 a |
| HS2 | 105.13 ± 2.46 c | 6.51 ± 0.25 ab | 0.64 ± 0.09 a | 0.83 ± 0.10 a | 17.32 ± 2.07 a |
| HS3 | 228.98 ± 10.46 a | 6.21 ± 0.02 b | 0.68 ± 0.05 a | 0.75 ± 0.09 a | 19.20 ± 3.33 a |
| HS4 | 181.85 ± 3.04 b | 5.78 ± 0.14 c | 0.66 ± 0.06 a | 0.58 ± 0.12 a | 16.06 ± 2.21 a |
| HS5 | 169.45 ± 16.33 b | 5.71 ± 0.65 c | 0.57 ± 0.03 a | 0.59 ± 0.12 a | 17.56 ± 0.69 a |
| HS6 | 110.82 ± 12.57 c | 5.32 ± 0.21 d | 0.48 ± 0.04 a | 0.55 ± 0.10 a | 16.67 ± 2.49 a |
| Bacteria Name | Similar Species | GenBank Accession Number | Inhibition Rate for Fusarium oxysporum (%) |
|---|---|---|---|
| 2M311 | Bacillus tequilensis | OR334900 | 48.66 ± 3.05 |
| R38 | Bacillus velezensis | OR334901 | 69.66 ± 4.16 |
| LB6-1 | Bacillus paralicheniformis | OR334899 | 74.66 ± 1.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Q.; Li, L.; Han, X.; Jiang, X.; Zhou, Y.; Wang, T.; Zhuang, Y.; Ruan, Z. Structural and Functional Analysis of the Bacterial Community in the Soil of Continuously Cultivated Lonicera japonica Thunb. and Screening Antagonistic Bacteria for Plant Pathogens. Agronomy 2024, 14, 260. https://doi.org/10.3390/agronomy14020260
Ma Q, Li L, Han X, Jiang X, Zhou Y, Wang T, Zhuang Y, Ruan Z. Structural and Functional Analysis of the Bacterial Community in the Soil of Continuously Cultivated Lonicera japonica Thunb. and Screening Antagonistic Bacteria for Plant Pathogens. Agronomy. 2024; 14(2):260. https://doi.org/10.3390/agronomy14020260
Chicago/Turabian StyleMa, Qingyun, Lin Li, Xiaoyan Han, Xu Jiang, Yiqing Zhou, Tan Wang, Yan Zhuang, and Zhiyong Ruan. 2024. "Structural and Functional Analysis of the Bacterial Community in the Soil of Continuously Cultivated Lonicera japonica Thunb. and Screening Antagonistic Bacteria for Plant Pathogens" Agronomy 14, no. 2: 260. https://doi.org/10.3390/agronomy14020260
APA StyleMa, Q., Li, L., Han, X., Jiang, X., Zhou, Y., Wang, T., Zhuang, Y., & Ruan, Z. (2024). Structural and Functional Analysis of the Bacterial Community in the Soil of Continuously Cultivated Lonicera japonica Thunb. and Screening Antagonistic Bacteria for Plant Pathogens. Agronomy, 14(2), 260. https://doi.org/10.3390/agronomy14020260