
Effect of Nitrogen Addition on Tiger Nut (Cyperus esculentus L.) Rhizosphere Microbial Diversity and Drive Factions of Rhizosphere Soil Multifunctionality in Sandy Farmland


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Study Site
2.2. Experimental Design
2.3. Soil Property Analysis and Function Assessment
2.4. DNA Extraction and High-Throughput Sequencing
2.5. Sequence Preprocessing
2.6. Statistical Analysis
3. Results
3.1. Response of Rhizosphere Soil Nutrients and Soil Extracellular Enzyme Activities to N Addition
3.2. Responses of Rhizosphere Microbial Community Composition and Diversity to N Addition
3.3. Influences of N Addition on the Complexity of the Microbial Co-Occurrence Network in Rhizosphere Soil
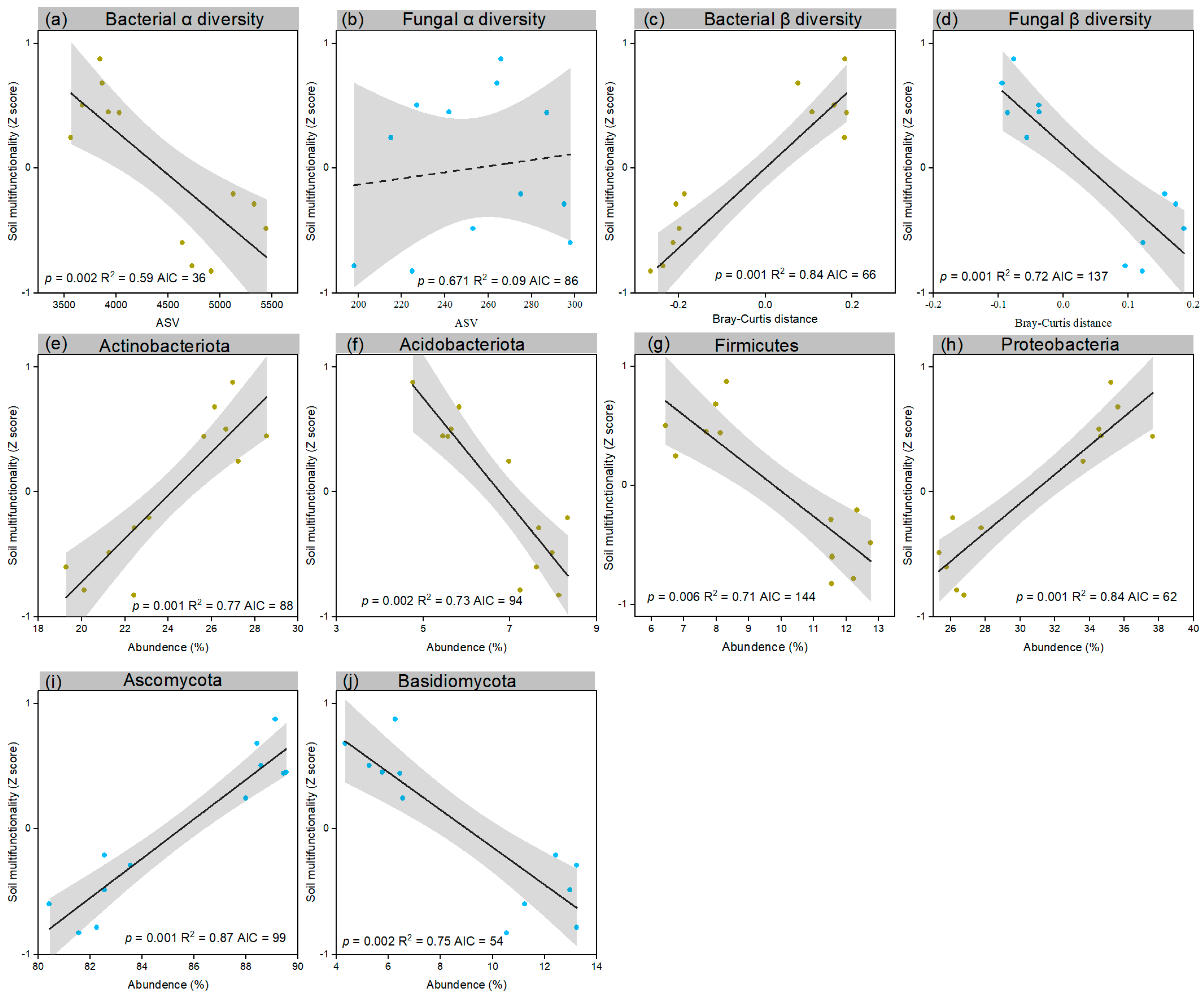
3.4. Rhizosphere Microbial Diversity–Soil Multifunctionality Relationships
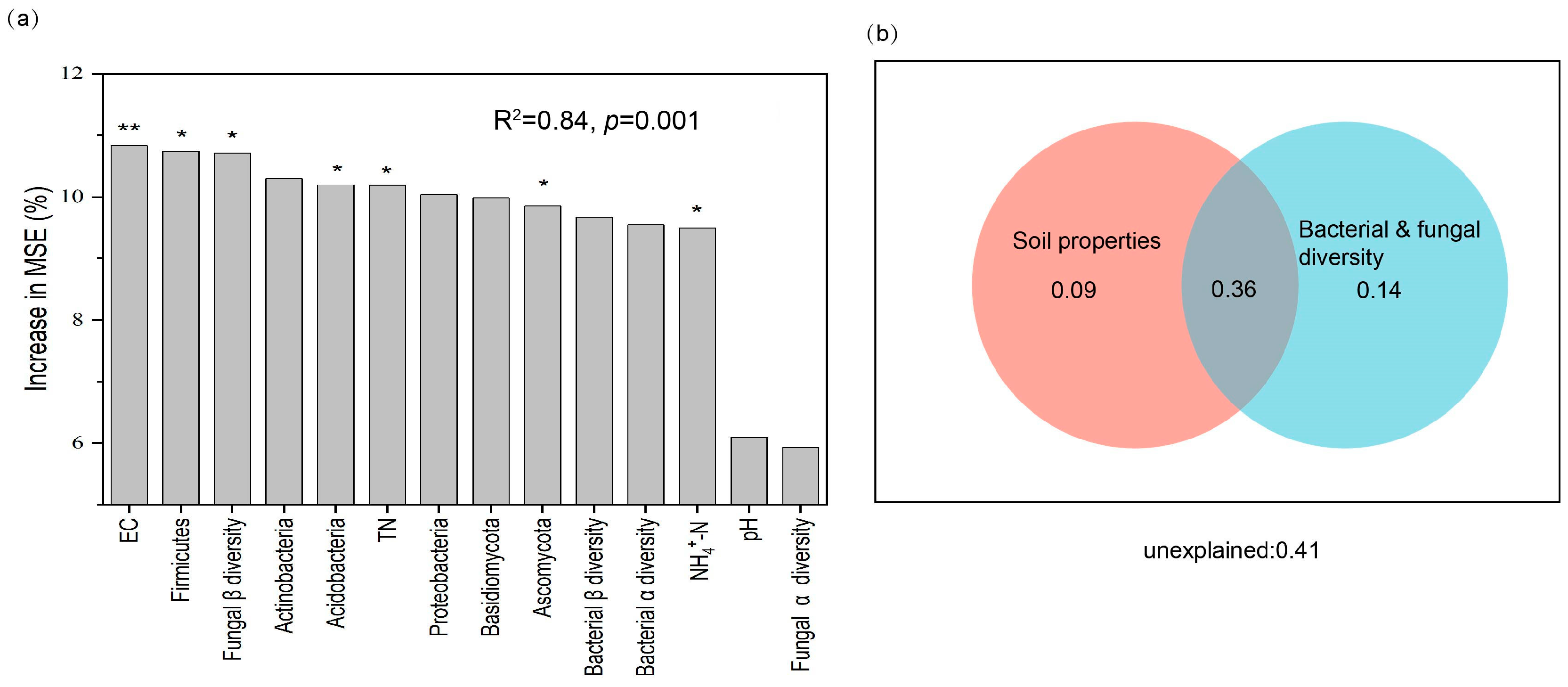
3.5. Potential Drivers of Root Zone Soil Multifunctionality
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Snapp, S.; Sapkota, T.B.; Chamberlin, J.; Cox, C.M.; Gameda, S.; Jat, M.L.; Marenya, P.; Mottaleb, K.A.; Negra, C.; Senthilkumar, K. Spatially differentiated nitrogen supply is key in a global food–fertilizer price crisis. Nat. Sustain. 2023, 6, 1268–1278. [Google Scholar] [CrossRef]
- Shahzad, A.N.; Qureshi, M.K.; Wakeel, A.; Misselbrook, T. Crop production in Pakistan and low nitrogen use efficiencies. Nat. Sustain. 2019, 2, 1106–1114. [Google Scholar] [CrossRef]
- Akhtar, K.; Wang, W.; Ren, G.; Khan, A.; Feng, Y.; Yang, G.; Wang, H. Integrated use of straw mulch with nitrogen fertilizer improves soil functionality and soybean production. Environ. Int. 2019, 132, 105092. [Google Scholar] [CrossRef] [PubMed]
- Hui, K.; Xi, B.; Tan, W.; Song, Q. Long-term application of nitrogen fertilizer alters the properties of dissolved soil organic matter and increases the accumulation of polycyclic aromatic hydrocarbons. Environ. Res. 2022, 215, 114267. [Google Scholar] [CrossRef] [PubMed]
- Ulm, F.; Gouveia, C.; Dias, T.; Cruz, C. N fertilization in a Mediterranean ecosystem alters N and P turnover in soil, roots and the ectomycorrhizal community. Soil Biol. Biochem. 2017, 113, 60–70. [Google Scholar] [CrossRef]
- Kimmel, K.; Furey, G.N.; Hobbie, S.E.; Isbell, F.; Tilman, D.; Reich, P.B. Diversity-dependent soil acidification under nitrogen enrichment constrains biomass productivity. Glob. Change Biol. 2020, 26, 6594–6603. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zheng, T.; Li, Y.; Zheng, X. A critical review of the central role of microbial regulation in the nitrogen biogeochemical process: New insights for controlling groundwater nitrogen contamination. J. Environ. Manag. 2023, 328, 116959. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chi, J.; Lu, X.; Cai, Y.; Jiang, H.; Zhang, Q.; Zhang, K. Fungal-bacterial composition and network complexity determine soil multifunctionality during ecological restoration. Catena 2023, 230, 107251. [Google Scholar] [CrossRef]
- Xue, R.; Wang, C.; Zhao, L.; Cao, J.; Liu, M.; Zhang, D. Agricultural intensification weakens soil multifunctionality by reducing fungal diversity. Appl. Soil Ecol. 2023, 189, 104900. [Google Scholar] [CrossRef]
- Zheng, Q.; Hu, Y.; Kosina, S.M.; Van Goethem, M.W.; Tringe, S.G.; Bowen, B.P.; Northen, T.R. Conservation of beneficial microbes between the rhizosphere and the cyanosphere. New Phytol. 2023, 240, 1246–1258. [Google Scholar] [CrossRef]
- Afridi, M.S.; Fakhar, A.; Kumar, A.; Ali, S.; Medeiros, F.H.; Muneer, M.A.; Ali, H.; Saleem, M. Harnessing microbial multitrophic interactions for rhizosphere microbiome engineering. Microbiol. Res. 2022, 265, 127199. [Google Scholar] [CrossRef] [PubMed]
- Dai, T.; Wen, D.; Bates, C.T.; Wu, L.; Guo, X.; Liu, S.; Su, Y.; Lei, J.; Zhou, J.; Yang, Y. Nutrient supply controls the linkage between species abundance and ecological interactions in marine bacterial communities. Nat. Commun. 2022, 13, 175. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Wu, X.; He, Y.; Li, Y.; Li, X.; Yu, X.; Shi, J. Mutual feedback mechanisms between functional traits and soil nutrients drive adaptive potential of tiger nuts (Cyperus esculentus L.) in marginal land. Plant Soil 2023, 495, 177–194. [Google Scholar] [CrossRef]
- Pascual, B.; Maroto, J.V.; López-Galarza, S.; Sanbautista, A.; Alagarda, J. Chufa (Cyperus esculentus L. var. sativus Boeck.): An unconventional crop. Studies related to applications and cultivation. Econ. Bot. 2000, 54, 439–448. [Google Scholar] [CrossRef]
- Wetters, S.; Häser, A.; Ehrlich, T.; Scheitle, C.; Nick, P. Tracing tiger nut (C. esculentus L.): Functional food from the colossal Cyperus genus. Eur. Food Res. Technol. 2023, 250, 225–238. [Google Scholar] [CrossRef]
- Pascual-Seva, N.; Pascual, B. Determination of crop coefficient for chufa crop (Cyperus esculentus L. var. sativus Boeck.) for sustainable irrigation scheduling. Sci. Total Environ. 2021, 768, 144975. [Google Scholar] [CrossRef]
- Scarabel, L.; Farinati, S.; Sattin, M. Occurrence of resistance to ALS inhibitors in European Cyperus esculentus L.: Characterisation and implications for management. Agronomy 2020, 10, 1133. [Google Scholar] [CrossRef]
- Ayeni, A.O. Hoop house and field evaluation of tigernut (Cyperus esculentus l. var. sativus boeck) selections in New Jersey, USA. Plants 2022, 11, 897. [Google Scholar] [CrossRef]
- Venturi, V.; Keel, C. Signaling in the rhizosphere. Trends Plant Sci. 2016, 21, 187–198. [Google Scholar] [CrossRef]
- Rüger, L.; Feng, K.; Dumack, K.; Freudenthal, J.; Chen, Y.; Sun, R.; Wilson, M.; Yu, P.; Sun, B.; Deng, Y. Assembly patterns of the rhizosphere microbiome along the longitudinal root axis of maize (Zea mays L.). Front. Microbiol. 2021, 12, 614501. [Google Scholar] [CrossRef]
- Qu, Q.; Wang, Z.; Gan, Q.; Liu, R.T.; Xu, H.W. Impact of drought on soil microbial biomass and extracellular enzyme activity. Front. Plant Sci. 2023, 14, 1221288. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.L.; Chen, Y.M.; Xu, W.J.; Liu, M. Effects of warming on the stoichiometry of soil microbial biomass and extracellular enzymes in an alpine shrubland. Geoderma 2023, 430, 116329. [Google Scholar] [CrossRef]
- Ke, W.S.; Li, C.X.; Zhu, F.; Luo, X.H.; Feng, J.P.; Li, X.; Jiang, Y.F.; Wu, C.; Hartley, W.; Xue, S.G. Effect of potentially toxic elements on soil multifunctionality at a lead smelting site. J. Hazard. Mater. 2023, 454, atr. [Google Scholar] [CrossRef] [PubMed]
- Sauvadet, M.; Saj, S.; Freschet, G.T.; Essobo, J.D.; Enock, S.; Becquer, T.; Tixier, P.; Harmand, J.M. Cocoa agroforest multifunctionality and soil fertility explained by shade tree litter traits. J. Appl. Ecol. 2020, 57, 476–487. [Google Scholar] [CrossRef]
- Byrnes, J.E.K.; Gamfeldt, L.; Isbell, F.; Lefcheck, J.S.; Griffin, J.N.; Hector, A.; Cardinale, B.J.; Hooper, D.U.; Dee, L.E.; Duffy, J.E. Investigating the relationship between biodiversity and ecosystem multifunctionality: Challenges and solutions. Methods Ecol. Evol. 2014, 5, 111–124. [Google Scholar] [CrossRef]
- Velmurugan, A.; Swarnam, T.P.; Jaisankar, I.; Swain, S.; Subramani, T. Vegetation-soil-microbial diversity influences ecosystem multifunctionality across different tropical coastal ecosystem types. Trop. Ecol. 2022, 63, 273–285. [Google Scholar] [CrossRef]
- Luo, J.; Liao, G.; Banerjee, S.; Gu, S.; Liang, J.; Guo, X.; Zhao, H.; Liang, Y.; Li, T. Long-term organic fertilization promotes the resilience of soil multifunctionality driven by bacterial communities. Soil Biol. 2023, 177, 108922. [Google Scholar] [CrossRef]
- Liu, Y.X.; Qin, Y.; Chen, T.; Lu, M.P.; Qian, X.B.; Guo, X.X.; Bai, Y. A practical guide to amplicon and metagenomic analysis of microbiome data. Protein Cell 2021, 12, 315–330. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, 2584. [Google Scholar] [CrossRef]
- Edgar, R.C. UNOISE2: Improved error-correction for Illumina 16S and ITS amplicon sequencing. bioRxiv 2016. [Google Scholar] [CrossRef]
- Cole, J.R.; Wang, Q.; Cardenas, E.; Fish, J.; Chai, B.; Farris, R.J.; Kulam-Syed-Mohideen, A.S.; McGarrell, D.M.; Marsh, T.; Garrity, G.M.; et al. The Ribosomal Database Project: Improved alignments and new tools for rRNA analysis. Nucleic Acids Res. 2009, 37, D141–D145. [Google Scholar] [CrossRef]
- Koljalg, U.; Larsson, K.H.; Abarenkov, K.; Nilsson, R.H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Hoiland, K.; Kjoller, R.; Larsson, E.; et al. UNITE: A database providing web-based methods for the molecular identification of ectomycorrhizal fungi. New Phytol. 2005, 166, 1063–1068. [Google Scholar] [CrossRef]
- Sun, Y.; Du, X.; Li, Y.; Han, X.; Fang, S.; Geisen, S.; Li, Q. Database and primer selections affect nematode community composition under different vegetations of Changbai Mountain. Soil Ecol. Lett. 2023, 5, 142–150. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef]
- Edgar, R. SINTAX: A simple non-Bayesian taxonomy classifier for 16S and ITS sequences. bioRxiv 2016. [Google Scholar] [CrossRef]
- Xu, S.D.; Liu, Y.X.; Cernava, T.; Wang, H.K.; Zhou, Y.Q.; Xia, T.; Cao, S.G.; Berg, G.; Shen, X.X.; Wen, Z.Y.; et al. Fusarium fruiting body microbiome member Pantoea agglomerans inhibits fungal pathogenesis by targeting lipid rafts. Nat. Microbiol. 2022, 7, 831–843. [Google Scholar] [CrossRef] [PubMed]
- Oren, A.; Garrity, G.M. Valid publication of the names of forty-two phyla of prokaryotes. J Int. J. Syst. Evol. Microbiol. 2021, 71, 005056. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.S.; Zou, Y.; Zhang, J.L.; Peres-Neto, P.R. Generalizing hierarchical and variation partitioning in multiple regression and canonical analyses using the rdacca.hp R package. Methods Ecol. Evol. 2022, 13, 782–788. [Google Scholar] [CrossRef]
- Dong, Y.; Gao, M.; Qiu, W.; Song, Z. Effect of microplastics and arsenic on nutrients and microorganisms in rice rhizosphere soil. Ecotoxicol. Environ. Saf. 2021, 211, 111899. [Google Scholar] [CrossRef] [PubMed]
- Preece, C.; Penuelas, J. A return to the wild: Root exudates and food security. Trends Plant Sci. 2020, 25, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yang, S.; Semenov, M.V.; Yao, F.; Ye, J.; Bu, R.; Ma, R.; Lin, J.; Kurganova, I.; Wang, X. Temperature sensitivity of SOM decomposition is linked with a K-selected microbial community. Glob. Change Biol. 2021, 27, 2763–2779. [Google Scholar] [CrossRef]
- Compant, S.; Samad, A.; Faist, H.; Sessitsch, A. A review on the plant microbiome: Ecology, functions, and emerging trends in microbial application. J. Adv. Res. 2019, 19, 29–37. [Google Scholar] [CrossRef]
- Wilhelm, R.C.; Muñoz-Ucros, J.; Weikl, F.; Pritsch, K.; Goebel, M.; Buckley, D.H.; Bauerle, T.L. The effects of mixed-species root zones on the resistance of soil bacteria and fungi to long-term experimental and natural reductions in soil moisture. Sci. Total Environ. 2023, 873, 162266. [Google Scholar] [CrossRef]
- Li, C.; Li, X.; Min, K.; Liu, T.; Li, D.; Xu, J.; Zhao, Y.; Li, H.; Chen, H.; Hu, F. Copiotrophic taxa in pig manure mitigate nitrogen limitation of soil microbial communities. Chemosphere 2022, 301, 134812. [Google Scholar] [CrossRef]
- Duan, Y.; Wang, X.; Wang, L.; Lian, J.; Wang, W.; Wu, F.; Li, Y.; Li, Y. Biogeographic patterns of soil microbe communities in the deserts of the Hexi Corridor, northern China. Catena 2022, 211, 106026. [Google Scholar] [CrossRef]
- Horton, D.J.; Theis, K.R.; Uzarski, D.G.; Learman, D.R. Microbial community structure and microbial networks correspond to nutrient gradients within coastal wetlands of the Laurentian Great Lakes. FEMS Microbiol. Ecol. 2019, 95, fiz033. [Google Scholar] [CrossRef]
- Wang, J.; Wang, X.; Liu, G.; Zhang, C.; Wang, G. Bacterial richness is negatively related to potential soil multifunctionality in a degraded alpine meadow. Ecol. Indic. 2021, 121, 106996. [Google Scholar] [CrossRef]
- Li, J.; Delgado-Baquerizo, M.; Wang, J.-T.; Hu, H.-W.; Cai, Z.-J.; Zhu, Y.-N.; Singh, B.K. Fungal richness contributes to multifunctionality in boreal forest soil. Soil Biol. Biochem. 2019, 136, 107526. [Google Scholar] [CrossRef]
- Hu, W.; Ran, J.; Dong, L.; Du, Q.; Ji, M.; Yao, S.; Sun, Y.; Gong, C.; Hou, Q.; Gong, H. Aridity-driven shift in biodiversity–soil multifunctionality relationships. Nat. Commun. 2021, 12, 5350. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-F.; Chen, P.; Wang, F.-H.; Han, W.-X.; Qiao, M.; Dong, W.-X.; Hu, C.-S.; Zhu, D.; Chu, H.-Y.; Zhu, Y.-G. The ecological clusters of soil organisms drive the ecosystem multifunctionality under long-term fertilization. Environ. Int. 2022, 161, 107133. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Xu, X.; Zhang, F.; Si, H.; Li, L.; Mao, G. The Preliminary Research on Shifts in Maize Rhizosphere Soil Microbial Communities and Symbiotic Networks under Different Fertilizer Sources. Agronomy 2023, 13, 2111. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Property | Bulk Density (g cm−3) | pH | Soil Organic Carbon (g kg−1) | Soil Total N (g kg−1) | Available N (mg kg−1) |
|---|---|---|---|---|---|
| Value | 1.48 | 8.97 | 1.22 | 0.46 | 26.32 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, X.; Chen, J.; Liu, J.; Cheng, Z.; Li, L.; Wang, J. Effect of Nitrogen Addition on Tiger Nut (Cyperus esculentus L.) Rhizosphere Microbial Diversity and Drive Factions of Rhizosphere Soil Multifunctionality in Sandy Farmland. Agronomy 2024, 14, 368. https://doi.org/10.3390/agronomy14020368
Zheng X, Chen J, Liu J, Cheng Z, Li L, Wang J. Effect of Nitrogen Addition on Tiger Nut (Cyperus esculentus L.) Rhizosphere Microbial Diversity and Drive Factions of Rhizosphere Soil Multifunctionality in Sandy Farmland. Agronomy. 2024; 14(2):368. https://doi.org/10.3390/agronomy14020368
Chicago/Turabian StyleZheng, Xu, Jun Chen, Jianguo Liu, Zhibo Cheng, Luhua Li, and Jiaping Wang. 2024. "Effect of Nitrogen Addition on Tiger Nut (Cyperus esculentus L.) Rhizosphere Microbial Diversity and Drive Factions of Rhizosphere Soil Multifunctionality in Sandy Farmland" Agronomy 14, no. 2: 368. https://doi.org/10.3390/agronomy14020368
APA StyleZheng, X., Chen, J., Liu, J., Cheng, Z., Li, L., & Wang, J. (2024). Effect of Nitrogen Addition on Tiger Nut (Cyperus esculentus L.) Rhizosphere Microbial Diversity and Drive Factions of Rhizosphere Soil Multifunctionality in Sandy Farmland. Agronomy, 14(2), 368. https://doi.org/10.3390/agronomy14020368