Effects of Dark Treatment on Lignin and Cellulose Synthesis in Celery


 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Celery Material
2.2. Determination of Lignin and Cellulose Content
2.3. Preparation and Histochemical Staining of Paraffin Sections
2.4. Total RNA Isolation and Real-Time Fluorescence Quantitative PCR Analysis
2.5. Statistical Analysis
3. Results
3.1. Morphological Characteristics of Celery during Dark Treatment and Conventional Cultivation
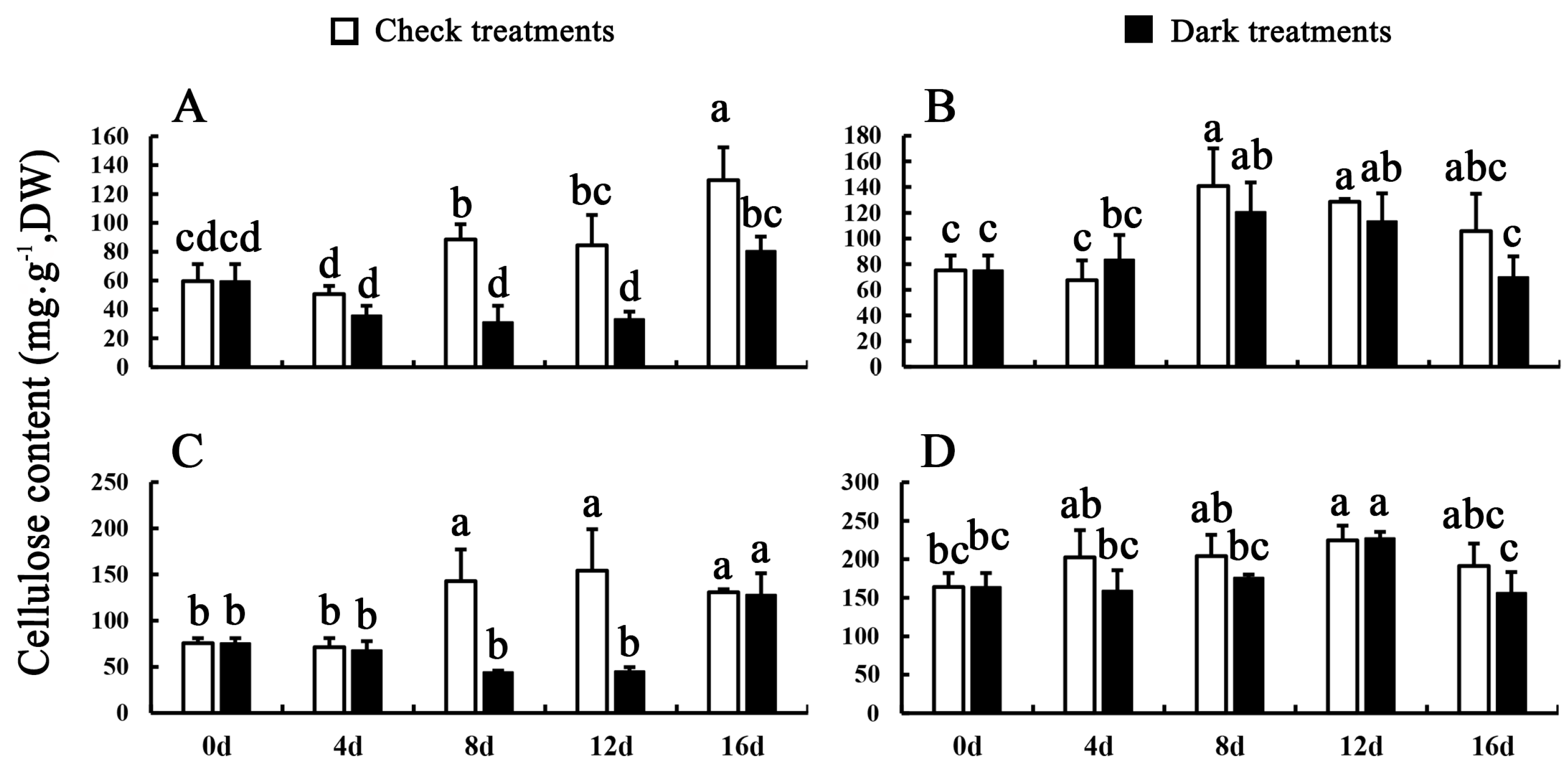
3.2. Lignin and Cellulose Contents of Celery during Dark Treatments and Control Conditions
3.3. Histochemical Analysis of Lignin Distribution in Celery Petioles
3.4. Expression Profiles of Lignin and Cellulose Metabolism-Related Genes in Celery during Dark Treatment and Control Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, M.Y.; Li, J.; Xie, F.J.; Zhou, J.; Sun, Y.; Luo, Y.; Zhang, Y.; Chen, Q.; Wang, Y.; Lin, Y.X.; et al. Combined evaluation of agronomic and quality traits to explore heat germplasm in celery (Apium graveolens L.). Sci. Hortic. 2023, 317, 112039. [Google Scholar] [CrossRef]
- Li, M.Y.; Hou, X.L.; Wang, F.; Tan, G.F.; Xu, Z.S.; Xiong, A.S. Advances in the research of celery, an important Apiaceae vegetable crop. Crit. Rev. Biotechnol. 2018, 38, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Sowbhagya, H.B. Chemistry, technology, and nutraceutical functions of celery (Apium graveolens L.): An overview. Crit. Rev. Food Sci. Nutr. 2014, 54, 389–398. [Google Scholar] [CrossRef]
- Patil, R.H.; Babu, R.L.; Naveen Kumar, M.; Kiran Kumar, K.M.; Hegde, S.M.; Ramesh, G.T.; Chidananda Sharma, S. Apigenin inhibits PMA-induced expression of pro-inflammatory cytokines and AP-1 factors in A549 cells. Mol. Cell Biochem. 2015, 403, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Khairullah, A.R.; Solikhah, T.I.; Ansori, A.N.; Hidayatullah, A.R.; Hartadi, E.B.; Ramandinianto, S.C.; Fadholly, A. Review on the Pharmacological and Health Aspects of Apium Graveolens or Celery: An Update. Syst. Rev. Pharm. 2021, 12, 606–612. [Google Scholar]
- Soliman, G.A. Dietary Fiber, Atherosclerosis, and Cardiovascular Disease. Nutrients 2019, 11, 1155. [Google Scholar] [CrossRef]
- Ioniță-Mîndrican, C.B.; Ziani, K.; Mititelu, M.; Oprea, E.; Neacșu, S.M.; Moroșan, E.; Dumitrescu, D.E.; Roșca, A.C.; Drăgănescu, D.; Negrei, C. Therapeutic Benefits and Dietary Restrictions of Fiber Intake: A State of the Art Review. Nutrients 2022, 14, 2641. [Google Scholar] [CrossRef]
- Larson, R.T.; McFarlane, H.E. Small but Mighty: An Update on Small Molecule Plant Cellulose Biosynthesis Inhibitors. Plant Cell Physiol. 2021, 62, 1828–1838. [Google Scholar] [CrossRef] [PubMed]
- Peter, G.; Neale, D. Molecular basis for the evolution of xylem lignification. Curr. Opin. Plant Biol. 2004, 7, 737–742. [Google Scholar] [CrossRef]
- Zhu, N.; Zhao, C.; Wei, Y.; Sun, C.; Wu, D.; Chen, K.S. Biosynthetic labeling with 3-O-propargylcaffeyl alcohol reveals in vivo cell-specific patterned lignification in loquat fruits during development and postharvest storage. Hortic. Res. 2021, 8, e61. [Google Scholar] [CrossRef]
- Mélida, H.; Largo-Gosens, A.; Novo-Uzal, E.; Santiago, R.; Pomar, F.; García, P.; García-Angulo, P.; Acebes, J.L.; Álvarez, J.; Encina, A. Ectopic lignification in primary cellulose-deficient cell walls of maize cell suspension cultures. J. Integr. Plant Biol. 2015, 57, 357–372. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, J.; Zou, S.; Liu, Z.; Huang, H.; Feng, C. Genome-wide analysis of the cellulose toolbox of Primulina eburnea, a calcium-rich vegetable. BMC Plant Biol. 2023, 23, e259. [Google Scholar] [CrossRef] [PubMed]
- Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin biosynthesis and structure. Plant Physiol. 2010, 153, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Barros, J. Lignin biosynthesis: Old roads revisited and new roads explored. Open Biol. 2019, 9, e190215. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Growth of the plant cell wall. Nat. Rev. Mol. Cell Biol. 2005, 6, 850–861. [Google Scholar] [CrossRef] [PubMed]
- Daras, G.; Templalexis, D.; Avgeri, F.; Tsitsekian, D.; Karamanou, K.; Rigas, S. Updating Insights into the Catalytic Domain Properties of Plant Cellulose synthase (CesA) and Cellulose synthase-like (Csl) Proteins. Molecules 2021, 26, 4335. [Google Scholar] [CrossRef] [PubMed]
- Schwerdt, J.G.; MacKenzie, K.; Wright, F.; Oehme, D.; Wagner, J.M.; Harvey, A.J.; Shirley, N.J.; Burton, R.A.; Schreiber, M.; Halpin, C.; et al. Evolutionary dynamics of the cellulose synthase gene superfamily in grasses. Plant Physiol. 2015, 168, 968–983. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Bak, G.; Burgin, T.; Barnes, W.J.; Mayes, H.B.; Peña, M.J.; Urbanowicz, B.R.; Nielsen, E. Biochemical and Genetic Analysis Identify CSLD3 as a beta-1,4-Glucan Synthase That Functions during Plant Cell Wall Synthesis. Plant Cell 2020, 32, 1749–1767. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Li, T.; Tan, G.F.; Huang, Y.; Wang, F.; Xiong, A.S. Efects of dark treatment and regular light recovery on the growth characteristics and regulation of chlorophyll in water dropwort. Plant Growth Regul. 2018, 85, 293–303. [Google Scholar] [CrossRef]
- Barry, C.S. The stay-green revolution: Recent progress in deciphering the mechanisms of chlorophyll degradation in higher plants. Plant Sci. 2009, 176, 325–333. [Google Scholar] [CrossRef]
- Sun, X.; Zhu, S.; Li, N.; Cheng, Y.; Zhao, J.; Qiao, X.; Lu, L.; Liu, S.; Wang, Y.; Liu, C.; et al. A Chromosome-Level Genome Assembly of Garlic (Allium sativum) Provides Insights into Genome Evolution and Allicin Biosynthesis. Mol. Plant. 2020, 13, 1328–1339. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, G.; Kong, Y.; Xu, H.; Xiao, B.; Liu, Y.; Zhou, H. A comparative analysis of the essential oils from two species of garlic seedlings cultivated in China: Chemical profile and anticoagulant potential. Food Funct. 2020, 11, 6020–6027. [Google Scholar] [CrossRef] [PubMed]
- Ai, P.; Xue, J.; Zhu, Y.; Tan, W.; Wu, Y.; Wang, Y.; Li, Z.; Shi, Z.; Kang, D.; Zhang, H.; et al. Comparative analysis of two kinds of garlic seedings: Qualities and transcriptional landscape. BMC Genom. 2023, 24, 87. [Google Scholar] [CrossRef]
- Jia, X.L.; Wang, G.L.; Xiong, F.; Yu, X.R.; Xu, Z.S.; Wang, F.; Xiong, A.S. De novo assembly, transcriptome characterization, lignin accumulation, and anatomic characteristics: Novel insights into lignin biosynthesis during celery leaf development. Sci. Rep. 2015, 5, e8259. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zhang, H.; Zhang, H.; Deng, X.W.; Wei, N. HY5 regulates nitrite reductase 1 (NIR1) and ammonium transporter1; 2 (AMT1; 2) in Arabidopsis seedlings. Plant Sci. 2015, 238, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Guo, Z.; Jiang, X.; Ahammed, G.J.; Zhou, Y. Light regulation of horticultural crop nutrient uptake and utilization. Hortic. Plant J. 2021, 7, 367–379. [Google Scholar] [CrossRef]
- Liu, H.; Yang, C.; Li, L. Shade-induced stem elongation in rice seedlings: Implication of tissue-specific phytohormone regulation. J. Integr. Plant Biol. 2016, 58, 614–617. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.J.; Chen, J.X.; Wang, Z.L.; Fan, Y.F.; Chen, S.Y.; Li, Z.L.; Liu, Q.L.; Li, Z.C.; Yang, F.; Yang, W.Y. Effects of light intensity and light quality on morphological and photosynthetic characteristics of soybean seedlings. Sci. Agric. Sin. 2018, 51, 2655–2663. [Google Scholar]
- Liu, W.G.; Ren, M.L.; Liu, T.; Du, Y.L.; Zhou, T.; Liu, X.M.; Liu, J.; Hussain, S.; Yang, W.Y. Effect of shade stress on lignin biosynthesis in soybean stems. J. Integr. Agric. 2018, 17, 1594–1604. [Google Scholar] [CrossRef]
- Liu, Y.; Jafari, F.; Wang, H.Y. Integration of light and hormone signaling pathways in the regulation of plant shade avoidance syndrome. aBIOTECH 2021, 2, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.C.; Wang, B.; Xie, G.Q.; Liu, Z.L.; Zhang, M.J.; Zhang, S.Q.; Cheng, X.G. Enrichment profile of GA4 is an important regulatory factor triggering rice dwarf. Sci. Agric. Sin. 2019, 52, 786–800. [Google Scholar]
- Tang, T.; Botwright Acuña, T.; Spielmeyer, W.; Richards, R.A. Effect of gibberellin-sensitive Rht18 and gibberellin-insensitive Rht-D1b dwarfing genes on vegetative and reproductive growth in bread wheat. J. Exp. Bot. 2021, 72, 445–458. [Google Scholar] [CrossRef] [PubMed]
- Li, C.H.; Luo, Y.L.; Jin, M.; Sun, S.F.; Wang, Z.L.; Li, Y. Response of Lignin Metabolism to Light Quality in Wheat Population. Front. Plant Sci. 2021, 12, e729647. [Google Scholar] [CrossRef] [PubMed]
- Su, G.X.; An, Z.F.; Zhang, W.H.; Liu, Y.L. Light promotes the synthesis of lignin through the production of H2O2 mediated by diamine oxidases in soybean hypocotyls. J. Plant Physiol. 2005, 162, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Boex-Fontvieille, E.; Davanture, M.; Jossier, M.; Zivy, M.; Hodges, M.; Tcherkez, G. Photosynthetic activity influences cellulose biosynthesis and phosphorylation of proteins involved therein in Arabidopsis leaves blades. J. Exp. Bot. 2014, 65, 4997–5010. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.M.; Zhang, W.J.; Ding, Y.F.; Zhang, J.W.; Cambula, E.D.; Weng, F.; Liu, Z.H.; Ding, C.Q.; Tang, S.; Chen, L.; et al. Shading Contributes to the Reduction of Stem Mechanical Strength by Decreasing Cell Wall Synthesis in Japonica Rice (Oryza sativa L.). Front. Plant Sci. 2017, 8, e881. [Google Scholar] [CrossRef] [PubMed]
- Falcioni, R.; Moriwaki, T.; Oliveira, D.M.; Andreotti, G.C.; De Souza, L.A.; Dos Santos, W.D.; Bonato, C.M.; Antunes, W.C. Increased Gibberellins and Light Levels Promotes Cell Wall Thickness and Enhance Lignin Deposition in Xylem Fibers. Front. Plant Sci. 2018, 9, e1391. [Google Scholar] [CrossRef]
- Teng, R.M.; Wang, Y.X.; Li, H.; Lin, S.J.; Liu, H.; Zhuang, J. Effects of shading on lignin biosynthesis in the leaf of tea plant (Camellia sinensis (L.) O. Kuntze). Mol. Genet. Genom. 2021, 296, 165–177. [Google Scholar] [CrossRef]
- Wen, B.; Zhang, Y.; Hussain, S.; Wang, S.; Zhang, X.; Yang, J.; Xu, M.; Qin, S.; Yang, W.; Liu, W. Slight Shading Stress at Seedling Stage Does not Reduce Lignin Biosynthesis or Affect Lodging Resistance of Soybean Stems. Agronomy 2020, 10, 544. [Google Scholar] [CrossRef]
- Li, X.; Weng, J.K.; Chapple, C. Improvement of biomass through lignin modification. Plant J. 2008, 54, 569–581. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.Z.; Wang, M.; Gyalpo, T.; Basang, Y. Stem lodging resistance in hulless barley: Transcriptome and metabolome analysis of lignin biosynthesis pathways in contrasting genotypes. Genomics 2021, 113 Pt 2, 935–943. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.H.; Shi, W.B.; Xia, X.; Zhao, D.Q.; Wu, Y.Q.; Tao, J. Morphological, microstructural and lignin-related responses of herbaceous peony stem to shading. Sci. Hortic. 2022, 293, e110734. [Google Scholar] [CrossRef]
- Luo, Y.; Chang, Y.; Li, C.; Wang, Y.; Cui, H.; Jin, M.; Wang, Z.; Li, Y. Shading decreases lodging resistance of wheat under different planting densities by altering lignin monomer composition of stems. Front. Plant Sci. 2022, 13, e1056193. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Li, X.; He, M.; Li, Y.; Lu, W.; Li, M.; Sun, B.; Zheng, Y. A Joint Transcriptomic and Metabolomic Analysis Reveals the Regulation of Shading on Lignin Biosynthesis in Asparagus. Int. J. Mol. Sci. 2023, 24, 1539. [Google Scholar] [CrossRef]
- Liu, Q.; Luo, L.; Zheng, L. Lignins: Biosynthesis and Biological Functions in Plants. Int. J. Mol. Sci. 2018, 19, 335. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lv, F.; Liu, J.; Ma, Y.; Wang, Y.; Chen, B.; Meng, Y.; Zhou, Z.; Oosterhuis, D.M. Effect of late planting and shading on cellulose synthesis during cotton fiber secondary wall development. PLoS ONE 2014, 9, e105088. [Google Scholar] [CrossRef]
- Huang, L.; Li, X.; Zhang, W.; Ung, N.; Liu, N.; Yin, X.; Li, Y.; Mcewan, R.E.; Dilkes, B.; Dai, M.; et al. Endosidin20 Targets the Cellulose Synthase Catalytic Domain to Inhibit Cellulose Biosynthesis. Plant Cell 2020, 32, 2141–2157. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′→3′) | Forward Primer (5′→3′) | Substance |
|---|---|---|---|
| AgActin | CTTCCTGCCATATATGATTGG | GCCAGCACCTCGATCTTCATG | Actin |
| AgPAL | GGTGGTGAAGTTGGGAGGAGAA | TGTTGCCGAGTGTGGTAATGTG | Lignin |
| AgC4H | TGGTTGTTGTGTCCTCTCCTGAT | GATTCTCCTCATCTTCCTCCAATGC | Lignin |
| Ag4CL | GGGAGATTGTGTAGCACCAGCA | GCCTGTTGAATGCCGAGTTTAT | Lignin |
| AgCCR | CAAGAGCAAAGCCGCTGAAGTT | GTGCAAAGCAGATCTTCAGG | Lignin |
| AgCAD | TGGGGGTTATCAACACTCCTTT | CGTCGTTTTTCTCCAACCTCTC | Lignin |
| AgHCT | TCTATCCGATGGCTGGGAGGTT | ATGGTCAGGTCAAGTCCCCGAG | Lignin |
| AgC3′H | CTCTACAACTTCTATCAACGGCTG | TCAAATCACTCCCACCTCTACT | Lignin |
| AgF5H | GCCAACCGTCCTGCTACCAT | TTCACCATGTCATCAACCTCTTCAC | Lignin |
| AgCCOAOMT | CAAACATTCAGAAGTTGGGCAC | GGCAAGGAGAGAATAACCAGTGTA | Lignin |
| AgCOMT | CTTGACCTTCTTGAGTCCATAGC | GCATCTTGTTGCTGCTGGGTAG | Lignin |
| AgPOD | GGAAGTGCTAGAACATTTGACC | CCTTATCTCTCCAGAAGACCCT | Lignin |
| AgLAC | GCTCTCCTTCAAGCACATTACT | TTTTGTTCCAGTTGTGGTCCCT | Lignin |
| AgCesA6 | CCTCGCTGTAGATTATCCTGTG | GAGATAGTCAACCTTTTCGGCA | cellulose |
| AgCslD3 | ACTTCAACTCCCACCTCCATCT | ATAGCATTTCCGTAGCCATAGG | cellulose |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, S.; Zhong, X.; Zhang, X.; Xiong, A.; Luo, Q.; Wang, K.; Li, M.; Tan, G. Effects of Dark Treatment on Lignin and Cellulose Synthesis in Celery. Agronomy 2024, 14, 896. https://doi.org/10.3390/agronomy14050896
Zhu S, Zhong X, Zhang X, Xiong A, Luo Q, Wang K, Li M, Tan G. Effects of Dark Treatment on Lignin and Cellulose Synthesis in Celery. Agronomy. 2024; 14(5):896. https://doi.org/10.3390/agronomy14050896
Chicago/Turabian StyleZhu, Shunhua, Xiulai Zhong, Xinqi Zhang, Aisheng Xiong, Qing Luo, Kun Wang, Mengyao Li, and Guofei Tan. 2024. "Effects of Dark Treatment on Lignin and Cellulose Synthesis in Celery" Agronomy 14, no. 5: 896. https://doi.org/10.3390/agronomy14050896
APA StyleZhu, S., Zhong, X., Zhang, X., Xiong, A., Luo, Q., Wang, K., Li, M., & Tan, G. (2024). Effects of Dark Treatment on Lignin and Cellulose Synthesis in Celery. Agronomy, 14(5), 896. https://doi.org/10.3390/agronomy14050896