
Detection of NAM-A1 Natural Variants in Bread Wheat Reveals Differences in Haplotype Distribution between a Worldwide Core Collection and European Elite Germplasm

Abstract
:1. Introduction
2. Results and Discussion
2.1. SNP Detection on NAM-A1, Genotyping and Mapping

{kind=link}
| Genotype | Frequency | |||
|---|---|---|---|---|
| Haplotype | SNP1 | SNP2 | Core Collection | Elite |
| NAM-A1a | C | A | 0.232 (85) | 0.083 (28) |
| NAM-A1b | C | Del | 0.003 (1) | 0.000 (0) |
| NAM-A1c | T | A | 0.477 (175) | 0.189 (63) |
| NAM-A1d | T | Del | 0.215 (79) | 0.716 (239) |
| Undefined | 0.074 (27) | 0.012 (4) | ||
2.2. Effect of NAM-A1 Haplotypes
| SNP1 | SNP2 | Haplotype | GY | TKW | SA | KS | GPC | GPD | NHI | %N_S |
|---|---|---|---|---|---|---|---|---|---|---|
| C | A | NAM-A1a | 6976c | 41.3b | 421a | 40.4b | 10.46a | 0.20a | 81.17ab | 0.41a |
| T | A | NAM-A1c | 7241b | 41.6b | 413a | 42.5a | 10.15b | 0.04ab | 81.47a | 0.41a |
| T | Del | NAM-A1d | 7799a | 42.7a | 411a | 43.0a | 9.79c | -0.09b | 80.98b | 0.42b |
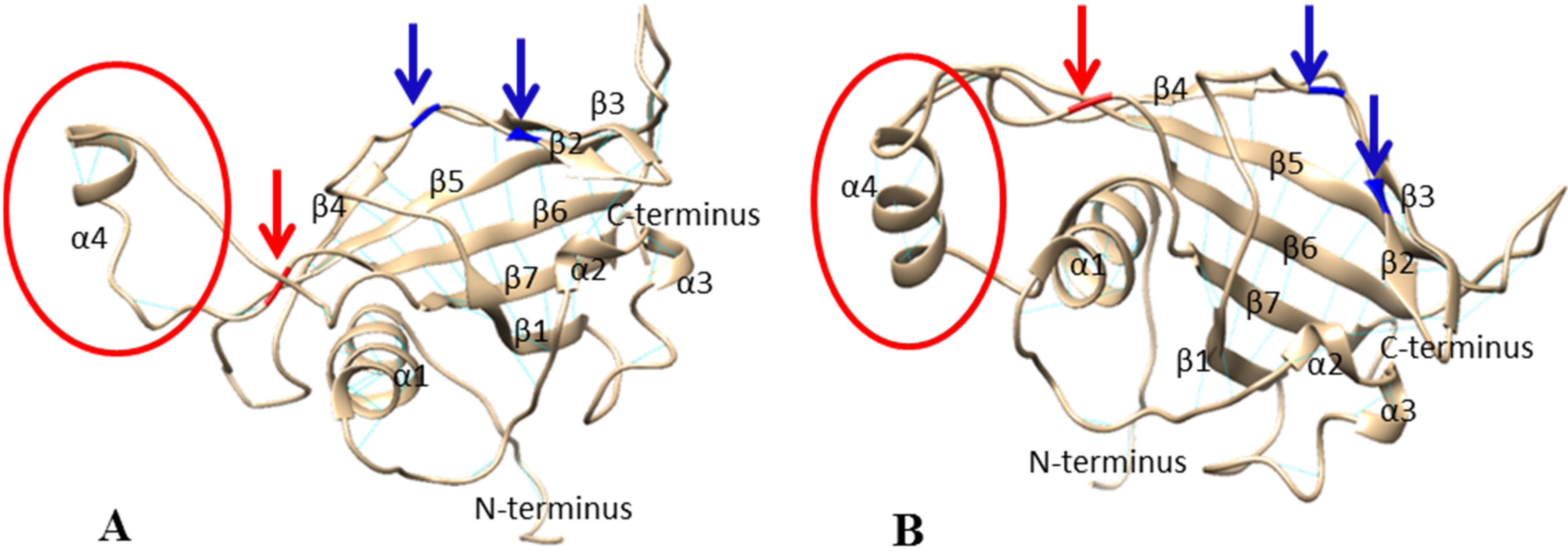
2.3. 3D Conformation of NAM-A1 NAC Domain

3. Experimental Section
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sanford, D.A.; MacKown, C.T. Variation in nitrogen use efficiency among soft red winter wheat genotypes. Theor. Appl. 1986, 72, 158–163. [Google Scholar] [CrossRef]
- Uauy, C.; Brevis, J.C.; Dubcovsky, J. The high grain protein content gene Gpc-B1 accelerates senescence and has pleiotropic effects on protein content in wheat. J. Exp. Bot. 2006, 57, 2785–2794. [Google Scholar] [CrossRef] [PubMed]
- Uauy, C.; Distelfeld, A.; Fahima, T.; Blechl, A.; Dubcovsky, J. A NAC gene regulating senescence improves grain protein, zinc, and iron content in wheat. Science 2006, 314, 1298–1301. [Google Scholar] [CrossRef] [PubMed]
- Distelfeld, A.; Pearce, S.P.; Avni, R.; Scherer, B.; Uauy, C.; Piston, F.; Slade, A.; Zhao, R.; Dubcovsky, J. Divergent functions of orthologous NAC transcription factors in wheat and rice. Plant Mol. Biol. 2012, 78, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Gaju, O.; Allard, V.; Martre, P.; Snape, J.W.; Heumez, E.; Le Gouis, J.; Moreau, D.; Bogard, M.; Griffiths, S.; Orford, S.; et al. Identification of traits to improve the nitrogen-use efficiency of wheat genotypes. Field Crops Res. 2011, 123, 139–152. [Google Scholar] [CrossRef]
- Hagenblad, J.; Asplund, L.; Balfourier, F.; Ravel, C.; Leino, M. Strong presence of the high grain protein content allele of NAM-B1 in Fennoscandian wheat. Theor. Appl. Genet. 2012, 125, 1677–1686. [Google Scholar] [CrossRef] [PubMed]
- Distefeld, A.; Uauy, C.; Fahima, T.; Dubcosky, J. Physical map of the wheat high-grain protein content gene Gpc-B1 and development of a high-throughput molecular marker. New Physiol. 2006, 169, 753–763. [Google Scholar] [CrossRef]
- Avni, R.; Zhao, R.; Pearce, S.; Jun, Y.; Uauy, C.; Tabbita, F.; Fahima, T.; Slade, A.; Dubcovsky, J.; Distelfeld, A. Functional characterization of GPC-1 genes in hexaploid wheat. Planta 2014, 239, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Brevis, J.C.; Dubcovsky, J. Effects of the Chromosome Region Including the Gpc-B1 Locus on Wheat Grain and Protein Yield. Crop Sci. 2010, 50, 93–104. [Google Scholar] [CrossRef]
- Brevis, J.C.; Morris, C.F.; Manthey, F.; Dubcovsky, J. Effect of the grain protein content locus Gpc-B1 on bread and pasta quality. J. Cereal Sci. 2010, 51, 357–365. [Google Scholar] [CrossRef]
- Eagles, H.A.; McLean, R.; Eastwood, R.F.; Appelbee, M.J.; Cane, K.; Martin, P.J.; Wallwork, H. High-yielding lines of wheat carrying Gpc-B1 adapted to Mediterranean-type environments of the south and west of Australia. Crop Pasture Sci. 2014, 65, 854–861. [Google Scholar] [CrossRef]
- Tabbita, F.; Lewis, S.; Vouilloz, J.P.; Ortega, M.A.; Kade, M.; Abbate, P.E.; Barneix, A.J. Effects of the Gpc-B1 locus on high grain protein content introgressed into Argentinean wheat germplasm. Plant Breed. 2012, 132, 48–52. [Google Scholar] [CrossRef]
- Asplund, L.; Bergkvist, G.; Leino, M.W.; Westerbergh, A.; Weih, M. Swedish spring varieties with the rare high grain protein allele of NAM-B1 differ leaf senescence and grain mineral content. PLoS ONE 2013, 8, e59704. [Google Scholar] [CrossRef]
- Balfourier, F.; Roussel, V.; Strelchenko, P.; Exbrayat-Vinson, F.; Sourdille, P.; Boutet, G.; Koenig, J.; Ravel, C.; Mitrofanova, O.; Beckert, M.; et al. A worldwide bread wheat core collection arrayed in a 384-well plate. Theor. Appl. Genet. 2007, 114, 1265–1275. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Debbie, W.; Forrest, K.; Allen, A.; Chao, S.; Huang, B.E.; Maccaferri, M.; Salvi, S.; Milner, S.G.; Cattivelli, L.; et al. Characterization of polyploidy wheat genomic diversity using a high-density 90,000 single nucleotide polymorphism array. Plant Biotech. J. 2014, 12, 787–796. [Google Scholar] [CrossRef] [Green Version]
- Flint-Garcia, S.; Thornberry, J.M.; Buckler, E.S. Structure of linkage disequilibrium in plants. Annu. Rev. Plant Biol. 2003, 54, 357–374. [Google Scholar] [CrossRef] [PubMed]
- Cormier, F.; Faure, S.; Dubreuil, P.; Heumez, E.; Beauchêne, K.; Lafarge, S.; Praud, S.; Le Gouis, J. A multi-environmental study of recent breeding progress on nitrogen use efficiency in wheat (Triticum aestivum L.). Theor. Appl. Genet. 2013, 126, 3035–3048. [Google Scholar] [CrossRef] [PubMed]
- Feil, B. The inverse yield-protein relationships in cereals: Possibilities and limitations for genetically improving the grain protein yield. Trends Agron. 1997, 1, 103–119. [Google Scholar]
- Monaghan, J.M.; Snape, J.W.; Chojecki, A.J.S.; Kettlewell, P.S. The use of grain protein deviation for identifying wheat cultivars with high protein concentration and yield. Euphytica 2001, 122, 309–317. [Google Scholar] [CrossRef]
- Waters, B.M.; Uauy, C.; Dubcovsky, J.; Grusak, M.A. Wheat (Triticum aestivum) NAM proteins regulate the translocation of iron, zinc, and nitrogen compounds from vegetative tissues to grain. J. Exp. Bot. 2009, 60, 4263–4274. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wang, Q.; Xiong, L.; Lou, Z. A structural view of the conserved domain of rice stress-responsive NAC1. Protein Cell 2011, 2, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Olsen, A.N.; Ernst, H.A.; Lo Leggio, L.; Johansson, E.; Larsen, S.; Skriver, K. Preliminary crystallographic analysis of the NAC domain of ANAC, a member of the plant-specific NAC transcription factor family. Acta Crystallogr. D 2004, 60, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Ernst, H.A.; Olsen, A.N.; Skriver, K.; Larsen, S.; Lo Leggio, L. Structure of the conserved domain of ANAC, a member of the NAC family of transcription factors. EMBO Rep. 2004, 3, 297–303. [Google Scholar] [CrossRef]
- Olsen, A.N.; Ernst, H.A.; Leggio, L.L.; Skriver, K. NAC transcription factors: Structurally distinct, functionally diverse. Trends Plant Sci. 2005, 10, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Müller, C.W. Transcription factors: Global and detailed views. Curr. Opin. Struct. Biol. 2001, 11, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Duval, M.; Hsieh, T.F.; Kim, S.Y.; Thomas, T.L. Molecular characterization of AtNAM: A member of the Arabidopsis NAC domain superfamily. Plant Mol. Biol. 2002, 50, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Arnold, K.; Bordoli, L.; Kopp, J.; Schwede, T. The SWISS-MODEL workspace: A web based environment for protein structure homology modelling. Bioinformatics 2006, 22, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Kumar, J.; Jaiswal, V.; Kumar, A.; Kumar, N.; Mir, R.R.; Kumar, S.; Dhariwal, R.; Tyagi, S.; Khandelwal, M.; Prabhu, K.V.; et al. Introgression of a major gene for high grain protein content in some Indian bread wheat cultivars. Field Crops Res. 2011, 123, 226–233. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cormier, F.; Throude, M.; Ravel, C.; Gouis, J.L.; Leveugle, M.; Lafarge, S.; Exbrayat, F.; Duranton, N.; Praud, S. Detection of NAM-A1 Natural Variants in Bread Wheat Reveals Differences in Haplotype Distribution between a Worldwide Core Collection and European Elite Germplasm. Agronomy 2015, 5, 143-151. https://doi.org/10.3390/agronomy5020143
Cormier F, Throude M, Ravel C, Gouis JL, Leveugle M, Lafarge S, Exbrayat F, Duranton N, Praud S. Detection of NAM-A1 Natural Variants in Bread Wheat Reveals Differences in Haplotype Distribution between a Worldwide Core Collection and European Elite Germplasm. Agronomy. 2015; 5(2):143-151. https://doi.org/10.3390/agronomy5020143
Chicago/Turabian StyleCormier, Fabien, Mickaël Throude, Catherine Ravel, Jacques Le Gouis, Magalie Leveugle, Stéphane Lafarge, Florence Exbrayat, Nadine Duranton, and Sébastien Praud. 2015. "Detection of NAM-A1 Natural Variants in Bread Wheat Reveals Differences in Haplotype Distribution between a Worldwide Core Collection and European Elite Germplasm" Agronomy 5, no. 2: 143-151. https://doi.org/10.3390/agronomy5020143
APA StyleCormier, F., Throude, M., Ravel, C., Gouis, J. L., Leveugle, M., Lafarge, S., Exbrayat, F., Duranton, N., & Praud, S. (2015). Detection of NAM-A1 Natural Variants in Bread Wheat Reveals Differences in Haplotype Distribution between a Worldwide Core Collection and European Elite Germplasm. Agronomy, 5(2), 143-151. https://doi.org/10.3390/agronomy5020143