Seed Plumpness of Rice with Inhibition Expression of Starch Branching Enzymes and Starch Properties, Grain Position on Panicle



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
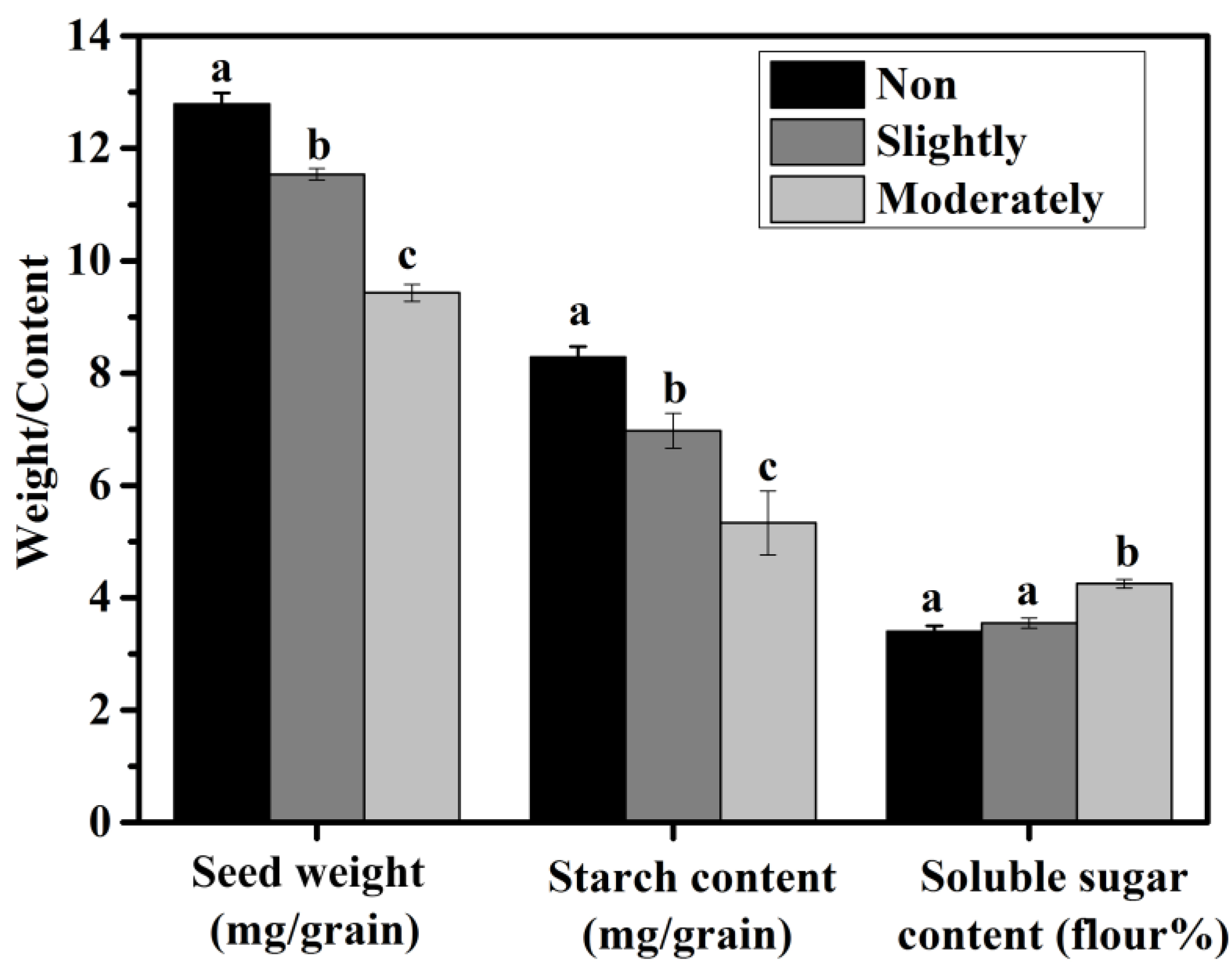
2.2. Determination of Seed Weight and Its Starch and Soluble Sugar Contents
2.3. Seed Sections for Light Microscopy
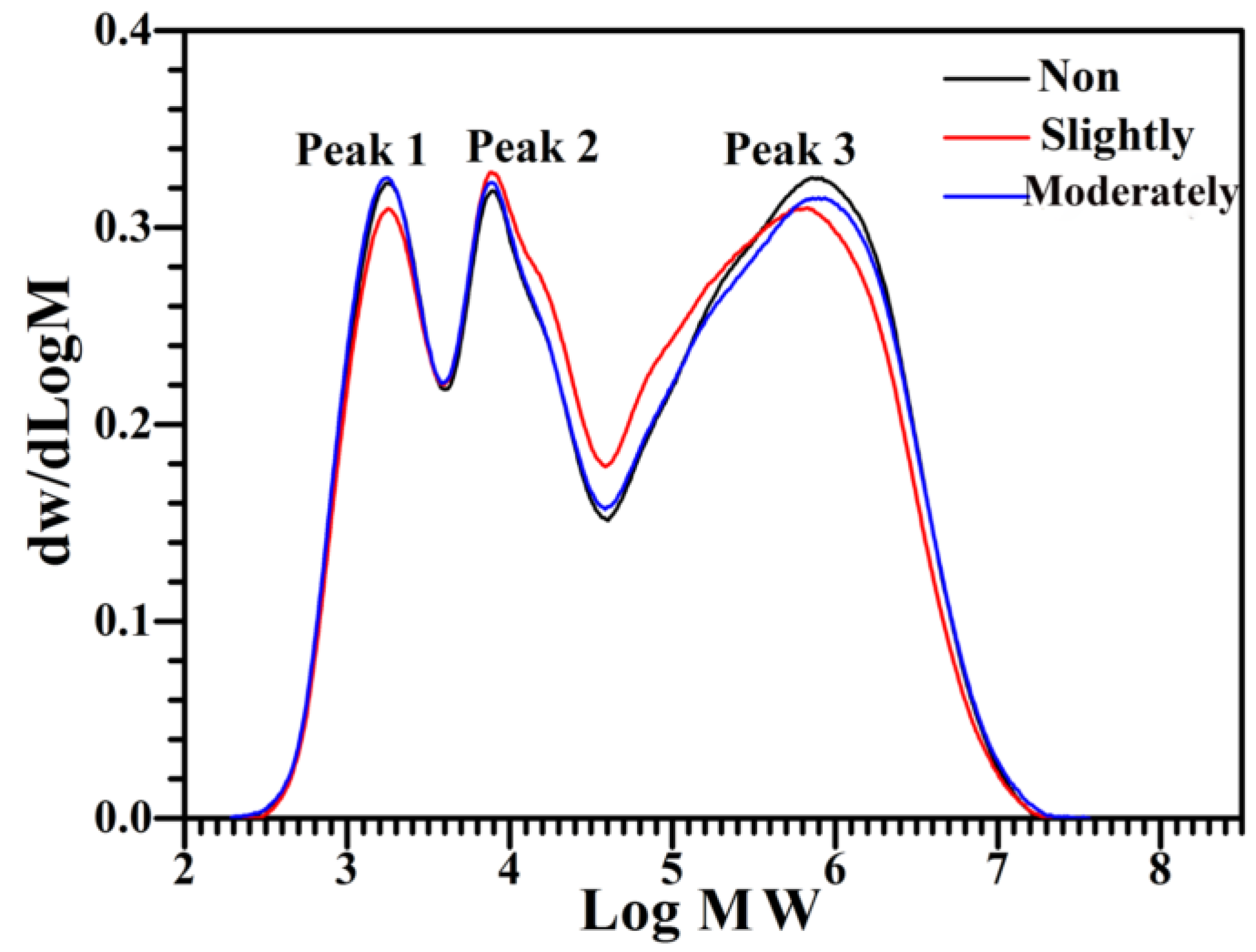
2.4. Molecular Weight Distribution of Starch
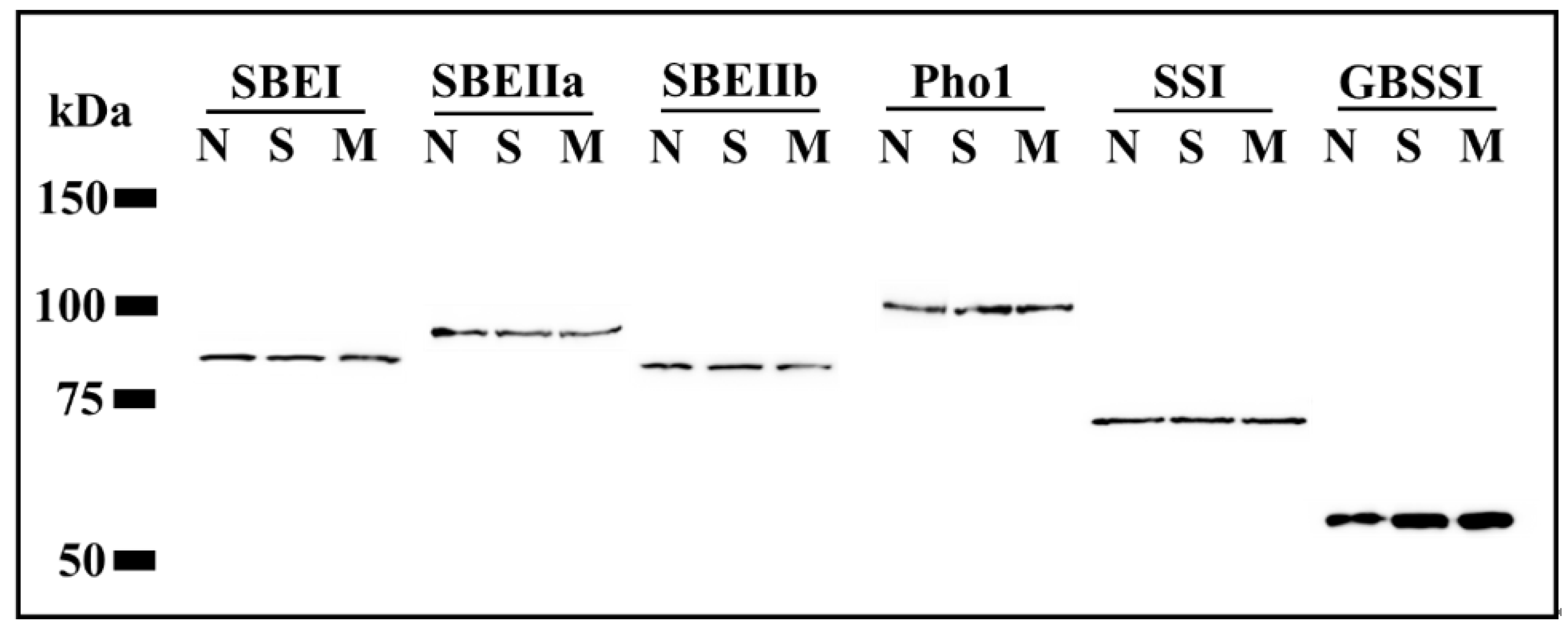
2.5. Western Blot
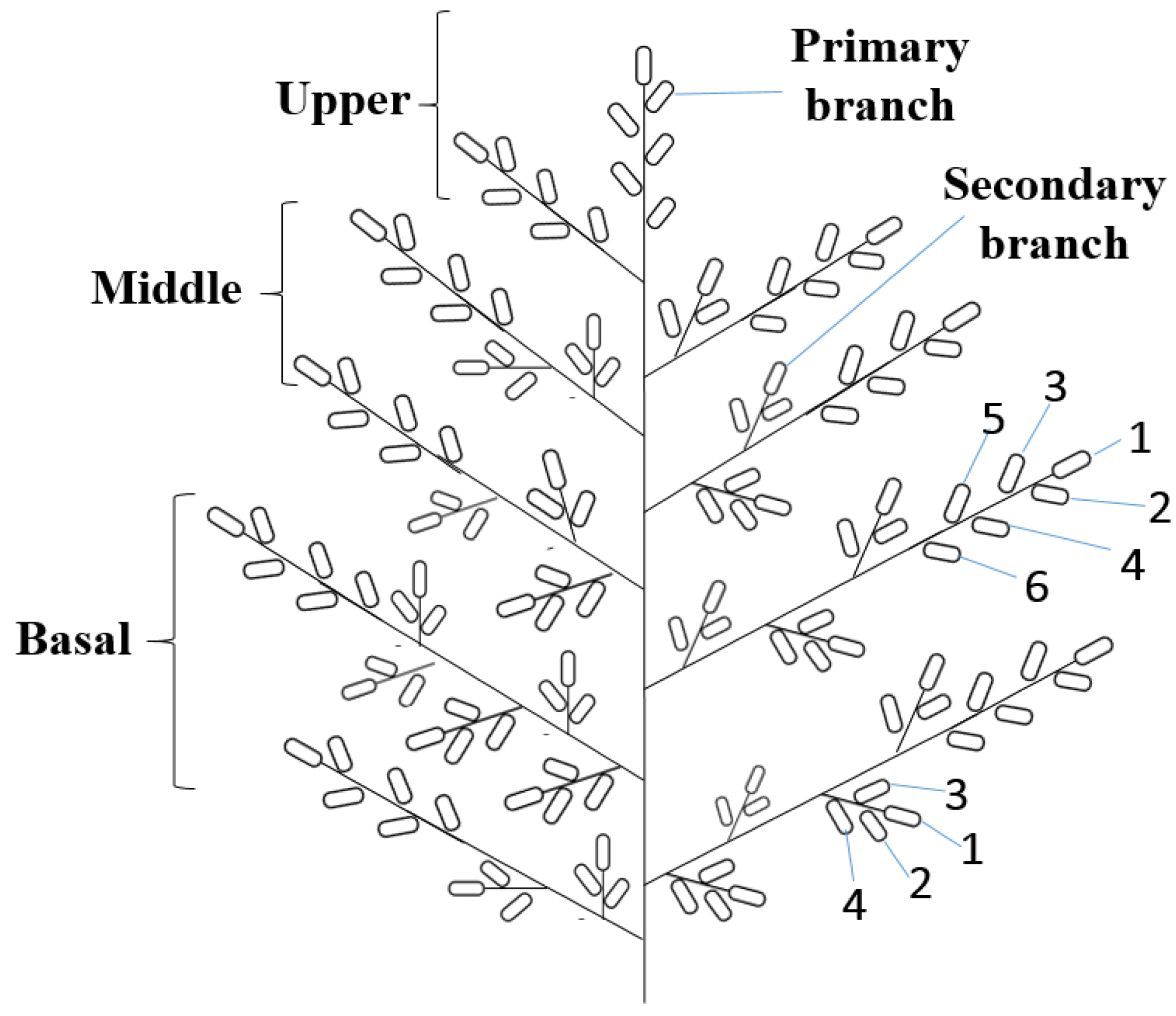
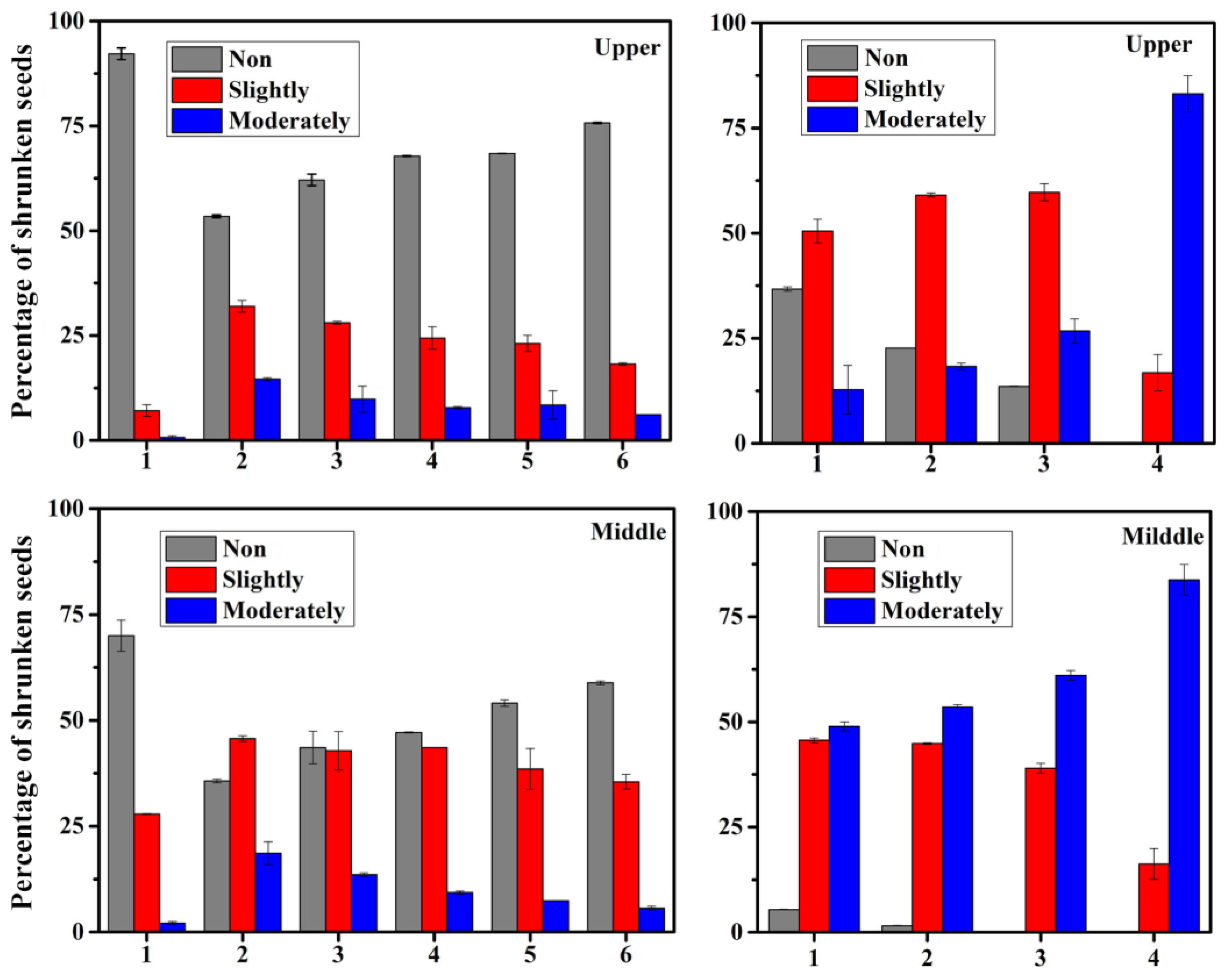
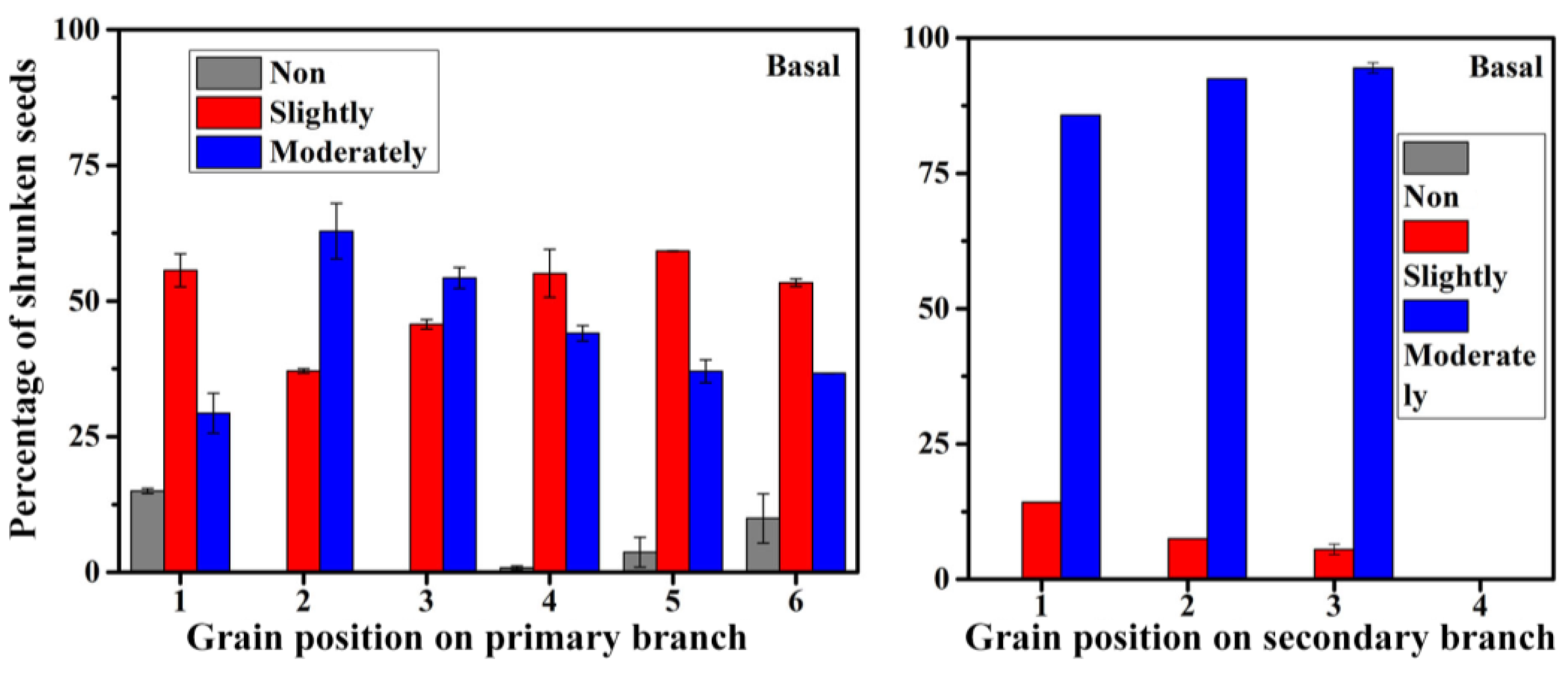
2.6. Analysis of Grain Position on Panicle
2.7. Statistical Analysis
3. Results
3.1. Characteristics of Differently Shrunken Seeds
3.2. Starch Morphology in Differently Shrunken Seeds
3.3. Molecular Weight Distribution of Starch from Differently Shrunken Seeds
3.4. Protein Amount of Amylose/Amylopectin Synthesis Key Enzymes in Differently Shrunken Seeds
3.5. Growing Position on Panicle of Differently Shrunken Seeds
4. Discussion
4.1. Morphology and Structure of Starch and Seed Plumpness
4.2. Grain Position on Panicle and Seed Plumpness
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Englyst, H.N.; Kingman, S.M.; Cummings, J.H. Classification and measurement of nutritionally important starch fractions. Eur. J. Clin. Nutr. 1992, 45, 33–50. [Google Scholar]
- Nugent, A.P. Health properties of resistant starch. Nutr. Bull. 2005, 30, 27–54. [Google Scholar] [CrossRef] [Green Version]
- Regina, A.; Bird, A.; Topping, D.; Bowden, S.; Freeman, J.; Barsby, T.; Kosar-Hashemi, B.; Li, Z.; Rahman, S.; Morell, M. High-amylose wheat generated by RNA interference improves indices of large-bowel health in rats. Proc. Natl. Acad. Sci. USA 2006, 10, 3546–3551. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Gu, M.; Meng, X.; Cheung, S.C.K.; Yu, H.; Huang, J.; Sun, Y.; Shi, Y.; Liu, Q. High-amylose rice improves indices of animal health in normal and diabetic rats. Plant Biotech. J. 2012, 10, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Jiang, H.; Campbell, M.; Blanco, M.; Jane, J.L. Characterization of maize amylose-extender (ae) mutant starches. Part I: relationship between resistant starch contents and molecular structures. Carbohyd. Polym. 2008, 74, 396–404. [Google Scholar] [CrossRef]
- Carciofi, M.; Blennow, A.; Jensen, S.L.; Shaik, S.S.; Henriksen, A.; Buléon, A.; Holm, P.B.; Hebelstrup, K.H. Concerted suppression of all starch branching enzyme genes in barley produces amylose-only starch granules. BMC Plant Biol. 2012, 12, 223. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Qian, Q.; Liu, Q.; Yan, M.; Liu, X.; Yan, C.; Liu, G.; Gao, Z.; Tang, S.; Zeng, D.; Wang, Y.; Yu, J.; Gu, M.; Li, J. Allelic diversities in rice starch biosynthesis lead to a diverse array of rice eating and cooking qualities. Proc. Natl. Acad. Sci. USA 2009, 106, 21760–21765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh, H.; Shibahara, K.; Tokunaga, T.; Nishi, A.; Tasaki, M.; Hwang, S.K.; Okita, T.W.; Kaneko, N.; Fujita, N.; Yoshida, M. Mutation of the plastidial α-glucan phosphorylase gene in rice affects the synthesis and structure of starch in the endosperm. Plant Cell 2008, 20, 1833–1849. [Google Scholar] [CrossRef] [PubMed]
- Nishi, A.; Nakamura, Y.; Tanaka, N.; Satoh, H. Biochemical and genetic analysis of the effects of amylose-extender mutation in rice endosperm. Plant Physiol. 2001, 127, 459–472. [Google Scholar] [CrossRef] [PubMed]
- Regina, A.; Kosar-Hashemi, B.; Ling, S.; Li, Z.; Rahman, S.; Morell, M. Control of starch branching in barley defined through differential RNAi suppression of starch branching enzyme IIa and IIb. J. Exp. Bot. 2010, 61, 1469–1482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butardo, V.M.; Fitzgerald, M.A.; Bird, A.R.; Gidley, M.J.; Flanagan, B.M.; Larroque, O.; Resurreccion, A.P.; Laidlaw, H.K.C.; Jobling, S.A.; Morell, M.K.; Rahman, S. Impact of down-regulation of starch branching enzyme IIb in rice by artificial microRNA- and hairpin RNA-mediated RNA silencing. J. Exp. Bot. 2011, 62, 4927–4941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slade, A.J.; McGuire, C.; Loeffl, D.; Mullenberg, J.; Skinner, W.; Fazio, G.; Holm, A.; Brandt, K.M.; Steine, M.N.; Goodstal, J.F.; Knauf, V.C. Development of high amylose wheat through TILLING. BMC Plant Biol. 2012, 12, 69. [Google Scholar] [CrossRef] [PubMed]
- Asai, H.; Abe, N.; Matsushima, R.; Crofts, N.; Oitome, NF.; Nakamura, Y.; Fujita, N. Deficiencies in both starch synthase IIIa and branching enzyme IIb lead to a significant increase in amylose in SSIIa inactive japonica rice seeds. J. Exp. Bot. 2014, 65, 5497–5507. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Thompson, D.B.; Guiltinan, M.J. Maize starch-branching enzyme isoforms and amylopectin structure. In the absence of starch branching enzyme IIb, the further absence of starch branching enzyme Ia leads to increased branching. Plant Physiol. 2004, 136, 3515–3523. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hu, P.; Lin, L.; Chen, Z.; Liu, Q.; Wei, C. Gradually decreasing starch branching enzyme expression is responsible for the formation of heterogeneous starch granules. Plant Physiol. 2018, 176, 582–595. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Qin, F.; Zhu, L.; Zhou, W.; Chen, Y.; Wang, Y.; Gu, M.; Liu, Q. Microstructure and ultrastructure of high-amylose rice resistant starch granules modified by antisense RNA inhibition of starch branching enzyme. J. Agric. Food Chem. 2010, 58, 1224–1232. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; Huang, J.; Zhao, L.; Liu, Q.; Zhang, C.; Wei, C. Heterogeneous structure and spatial distribution in endosperm of high-amylose rice starch granules with different morphologies. J. Agric. Food Chem. 2014, 62, 10143–10152. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Cai, J.; Han, W.; Huai, H.; Chen, Y.; Wei, C. Comparison of starches isolated from three different Trapa species. Food Hydrocolloid. 2014, 37, 174–181. [Google Scholar] [CrossRef]
- Zhao, L.; Pan, T.; Cai, C.; Wang, J.; Wei, C. Application of whole sections of mature cereal seeds to visualize the morphology of endosperm cell and starch and the distribution of storage protein. J. Cereal Sci. 2016, 71, 19–27. [Google Scholar] [CrossRef]
- Man, J.; Lin, L.; Wang, Z.; Wang, Y.; Liu, Q.; Wei, C. Different structures of heterogeneous starch granules from high-amylose rice. J. Agric. Food Chem. 2014, 62, 11254–11263. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Cai, C.; Gilber, R.G.; Li, E.; Wang, J.; Wei, C. Relationships between amylopectin molecular structures and functional properties of different-sized fractions of normal and high-amylose maize starches. Food Hydrocolloid. 2016, 52, 359–368. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Wang, W.; Cai, X. Modulation of amylose content by structure-based modification of OsGBSS1 activity in rice (Oryza sativa L.). Plant Biotech. J. 2014, 12, 1297–1307. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.H.; Sang, D.Z.; Wang, P.; Zhang, W.J.; Yang, J.C. Difference in chalky characters of the grains at different positions within a rice panicle. Acta Agron. Sin. 2006, 32, 103–111. [Google Scholar]
- Song, Y.; Jane, J. Characterization of barley starches of waxy, normal, and high amylose varieties. Carbohydr. Polym. 2000, 41, 365–377. [Google Scholar] [CrossRef]
- Liu, F.; Romanova, N.; Lee, E.A.; Ahmed, R.; Evans, M.; Gilbert, E.P.; Morell, M.K.; Emes, M.J.; Tetlow, I.J. Glucan affinity of starch synthase IIa determines binding of starch synthase I and starch-branching enzyme IIb to starch granules. Biochem. J. 2012, 448, 373–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.; Ahmed, R.; Kosar-Hashemi, B.; Larroque, O.; Butardo, V.M.; Tanner, G.J.; Colgrave, M.L.; Upadhyaya, N.M.; Tetlow, I.J.; Emes, M.J.; et al. The different effects of starch synthase IIa mutations or variation on endosperm amylose content of barley, wheat and rice are determined by the distribution of starch synthase I and starch branching enzyme IIb between the starch granule and amyloplast stroma. Theor. Appl. Genet. 2015, 128, 1407–1419. [Google Scholar] [PubMed]
- Itoh, Y.; Crofts, N.; Abe, M.; Hosaka, Y.; Fujita, N. Characterization of the endosperm starch and the pleiotropic effects of biosynthetic enzymes on their properties in novel mutant rice lines with high resistant starch and amylose content. Plant Sci. 2017, 258, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Ishimaru, T.; Hirose, T.; Matsuda, T.; Goto, A.; Takahashi, K.; Sasaki, H.; Terao, T.; Ishii, R.; Ohsugi, R.; Yamagishi, T. Expression patterns of genes encoding carbohydrate- metabolizing enzymes and their relationship to grain filling in rice (Oryza sativa L.): Comparison of caryopses located at different positions in a panicle. Plant Cell Physiol. 2005, 46, 620–628. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, J.; Wang, Z.; Liu, K.; Wang, P. Post-anthesis development of inferior and superior spikelets in rice in relation to abscisic acid and ethylene. J. Exp. Bot. 2006, 57, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Ye, N.; Yang, J.; Peng, X.; Zhang, J. Regulation of expression of starch synthesis genes by ethylene and ABA in relation to the development of rice inferior and superior spikelets. J. Exp. Bot. 2011, 62, 3907–3916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, K.; Panda, B.B.; Sekhar, S.; Kariali, E.; Mohapatra, K.P.; Shaw, P.B. Comparative proteomics of the superior and inferior nspikelets at the early grain filling stage in rice cultivars contrast for panicle compactness and ethylene evolution. J. Plant Physiol. 2016, 202, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, N.; Fujita, N.; Nishi, A.; Satoh, H.; Hosaka, Y.; Ugaki, M.; Kawasaki, S.; Nakamura, Y. The structure of starch can be manipulated by changing the expression levels of starch branching enzyme IIb in rice endosperm. Plant Biotech. J. 2004, 2, 507–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.X.; Xiong, H.; Zhu, Y.C.; Wang, G.X. Effect of source-sink ratio on grain filling and the source-sink characteristics of high yield varieties of mid-season hybrid rice. Sci. Agric. Sin. 2005, 38, 265–271. [Google Scholar]
- You, C.; Chen, L.; He, H.; Wu, L.; Wang, S.; Ding, Y.; Ma, C. iTRAQ-based proteome profile analysis of superior and inferior spikelets at early grain filling stage in japonica rice. BMC Plant Biol. 2017, 17, 100. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Zhang, J.; Cao, X. Grain sink strength may be related to the poor grain filling of indica-japonica (Oryza sativa) hybrids. Physiol. Plant. 2001, 112, 470–477. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak Area | Area Ratio of Peak 1 to Peak 2 | |||
|---|---|---|---|---|
| Peak 1 (%) | Peak 2 (%) | Peak 3 (%) | ||
| Non-shrunken seed | 20.9 ± 0.2 a | 23.8 ± 0.8 a | 55.3 ± 0.6 a | 0.88 ± 0.04 a |
| Slightly shrunken seed | 20.9 ± 1.8 a | 24.9 ± 1.7 a | 54.1 ± 0.2 a | 0.84 ± 0.13 a |
| Moderately shrunken seed | 21.4 ± 0.9 a | 24.7 ± 0.1 a | 53.9 ± 0.8 a | 0.87 ± 0.04 a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Chen, Z.; Hu, P.; Zhao, L.; Lin, L.; Wei, C. Seed Plumpness of Rice with Inhibition Expression of Starch Branching Enzymes and Starch Properties, Grain Position on Panicle. Agronomy 2018, 8, 252. https://doi.org/10.3390/agronomy8110252
Wang J, Chen Z, Hu P, Zhao L, Lin L, Wei C. Seed Plumpness of Rice with Inhibition Expression of Starch Branching Enzymes and Starch Properties, Grain Position on Panicle. Agronomy. 2018; 8(11):252. https://doi.org/10.3390/agronomy8110252
Chicago/Turabian StyleWang, Juan, Zichun Chen, Pan Hu, Lingxiao Zhao, Lingshang Lin, and Cunxu Wei. 2018. "Seed Plumpness of Rice with Inhibition Expression of Starch Branching Enzymes and Starch Properties, Grain Position on Panicle" Agronomy 8, no. 11: 252. https://doi.org/10.3390/agronomy8110252
APA StyleWang, J., Chen, Z., Hu, P., Zhao, L., Lin, L., & Wei, C. (2018). Seed Plumpness of Rice with Inhibition Expression of Starch Branching Enzymes and Starch Properties, Grain Position on Panicle. Agronomy, 8(11), 252. https://doi.org/10.3390/agronomy8110252