Linking Soil Microbial Properties with Plant Performance in Acidic Tropical Soil Amended with Biochar

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Design
2.2. Soil Sampling and Physico-Chemical Analysis
2.3. Microbial Biomass Analysis
2.4. Soil Enzyme Assay
2.5. DNA Extraction and Real-Time Quantitative Polymerase Chain Reaction (qPCR)
2.6. Plant Sampling and Performance Analysis
2.7. Statistical Analysis
3. Results
3.1. Effect of Biochar on Soil Properties.
3.2. Effect of Biochar on Soil Microbial Properties
3.3. Effect of Biochar on Maize Growth Performance
3.4. Relationship of Microbial Properties with Maize Biomass
4. Discussion
4.1. Effect of Biochar on Soil Properties
4.2. Effect of Biochar on Soil Microbial Properties
4.3. Effect of Biochar on Maize Biomass Yield and its Relationship with Microbial Properties
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lehmann, J.; Gaunt, J.; Rondon, M. Bio-char sequestration in terrestrial ecosystems—A review. Mitig. Adapt. Strateg. Glob. Chang. 2006, 11, 403–427. [Google Scholar] [CrossRef]
- Novotny, E.H.; Hayes, M.H.; Madari, B.E.; Bonagamba, T.J.; Azevedo, E.R.; Souza, A.A.; Song, G.; Nogueira, C.M.; Mangrich, A.S. Lessons from the Terra Preta de Índios of the Amazon region for the utilisation of charcoal for soil amendment. J. Braz. Chem. Soc. 2009, 20, 1003–1010. [Google Scholar] [CrossRef]
- Major, J.; Rondon, M.; Molina, D.; Riha, S.J.; Lehmann, J. Maize yield and nutrition during 4 years after biochar application to a Colombian savanna oxisol. Plant Soil 2010, 333, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Kloss, S.; Zehetner, F.; Wimmer, B.; Buecker, J.; Rempt, F.; Soja, G. Biochar application to temperate soils: Effects on soil fertility and crop growth under greenhouse conditions. J. Plant Nutr. Soil Sci. 2014, 177, 3–15. [Google Scholar] [CrossRef]
- Martinsen, V.; Alling, V.; Nurida, N.L.; Mulder, J.; Hale, S.E.; Ritz, C.; Rutherford, D.W.; Heikens, A.; Breedveld, G.D.; Cornelissen, G. pH of the addition of three biochars to acidic Indonesian mineral soils. Soil Sci. Plant Nutr. 2015, 61, 821–834. [Google Scholar] [CrossRef]
- Berihun, T.; Tadele, M.; Kebede, F. The application of biochar on soil acidity and other physico-chemical properties of soils in southern Ethiopia. J. Plant Nutr. Soil Sci. 2017, 180, 381–388. [Google Scholar] [CrossRef]
- Manickam, T.; Cornelissen, G.; Bachmann, R.T.; Ibrahim, I.Z.; Mulder, J.; Hale, S.E. Biochar application in Malaysian sandy and acid sulfate soils: Soil amelioration effects and improved crop production over two cropping seasons. Sustainability 2015, 7, 16756–16770. [Google Scholar] [CrossRef] [Green Version]
- Syuhada, A.B.; Shamshuddin, J.; Fauziah, C.I.; Rosenani, A.B.; Arifin, A. Biochar as soil amendment: Impact on chemical properties and corn nutrient uptake in a Podzol. Can. J. Soil Sci. 2016, 96, 400–412. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef] [Green Version]
- Brookes, P. The soil microbial biomass: Concept, measurement and applications in soil ecosystem research. Microbes Environ. 2001, 16, 131–140. [Google Scholar] [CrossRef]
- Lentz, R.D.; Ippolito, J.A. Biochar and manure affect calcareous soil and corn silage nutrient concentrations and uptake. J. Environ. Qual. 2012, 41, 1033–1043. [Google Scholar] [CrossRef] [PubMed]
- Kelly, N.; Calderón, F.C.; Acosta-martinez, V.; Mikha, M.M.; Benjamin, J.; Rutherford, D.W.; Rostad, C.E. Switchgrass biochar effects on plant biomass and microbial dynamics in two soils from different regions. Pedosphere 2015, 25, 329–342. [Google Scholar] [CrossRef]
- Lehmann, J.; Rillig, M.C.; Thies, J.; Masiello, C.A.; Hockaday, W.C.; Crowley, D. Biochar effects on soil biota—A review. Soil Biol. Biochem. 2011, 43, 1812–1836. [Google Scholar] [CrossRef]
- Simarani, K.; Halmi, M.F.A.; Abdullah, R. Short-term effects of biochar amendment on soil microbial community in humid tropics. Arch. Agron. Soil Sci. 2018, 64, 1847–1860. [Google Scholar] [CrossRef]
- Gillman, G.P.; Sumpter, E.A. Modification to the compulsive exchange method for measuring exchange characteristics of soils. Soil Res. 1986, 24, 61–66. [Google Scholar] [CrossRef]
- Walinga, I.; Kithome, M.; Novozamsky, I.; Houba, V.J.G.; Van der Lee, J.J. Spectrophotometric determination of organic carbon in soil. Commun. Soil Sci. Plant Anal. 1992, 23, 1935–1944. [Google Scholar] [CrossRef]
- Baethgen, W.E.; Alley, M.M. A manual colorimetric procedure for measuring ammonium nitrogen in soil and plant kjeldahl digests. Commun. Soil Sci. Plant Anal. 1989, 20, 961–969. [Google Scholar] [CrossRef]
- Cataldo, D.A.; Maroon, M.; Schrader, L.E.; Youngs, V.L. Rapid colorimetric determination of nitrate in plant tissue by nitration of salicylic acid. Commun. Soil Sci. Plant Anal. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Bray, R.H.; Kurtz, L.T. Determination of total, organic, and available forms of phosphorus in soils. Soil Sci. 1945, 59, 39–46. [Google Scholar] [CrossRef]
- Vance, E.D.; Brokes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Cai, Y.; Peng, C.; Qiu, S.; Li, Y.; Gao, Y. Dichromate digestion–spectrophotometric procedure for determination of soil microbial biomass carbon in association with fumigation–extraction. Commun. Soil Sci. Plant Anal. 2011, 42, 2824–2834. [Google Scholar] [CrossRef]
- Doyle, A.; Weintraub, M.N.; Schimel, J.P. Persulfate digestion and simultaneous colorimetric analysis of carbon and nitrogen in soil extracts. Soil Sci. Soc. Am. J. 2004, 68, 669–676. [Google Scholar] [CrossRef]
- Brookes, P.C.; Powlson, D.S.; Jenkinson, D.S. Phosphorus in the soil microbial biomass. Soil Biol. Biochem. 1984, 16, 169–175. [Google Scholar] [CrossRef]
- Wu, J.; He, Z.L.; Wei, W.X.; O’donnell, A.G.; Syers, J.K. Quantifying microbial biomass phosphorus in acid soils. Biol. Fertil. Soils 2000, 32, 500–507. [Google Scholar] [CrossRef]
- Eivazi, F.; Tabatabai, M.A. Glucosidases and galactosidases in soils. Soil Biol. Biochem. 1988, 20, 601–606. [Google Scholar] [CrossRef]
- Kandeler, E.; Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertil. Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Eivazi, F.; Tabatabai, M.A. Phosphatases in soils. Soil Biol. Biochem. 1977, 9, 167–172. [Google Scholar] [CrossRef]
- Fierer, N.; Jackson, J.A.; Vilgalys, R.; Jackson, R.B. Assessment of soil microbial community structure by use of taxon-specific quantitative PCR assays. Appl. Environ. Microbiol. 2005, 71, 4117–4120. [Google Scholar] [CrossRef] [PubMed]
- Ch’ng, H.Y.; Ahmed, O.H.; Nik, A.M.N.M. Biochar and compost influence the phosphorus availability, nutrients uptake, and growth of maize (Zea mays L.) in tropical acid soil. Pak. J. Agric. Sci. 2014, 51, 797–806. [Google Scholar]
- Alling, V.; Hale, S.E.; Martinsen, V.; Mulder, J.; Smebye, A.; Breedveld, G.D.; Cornelissen, G. The role of biochar in retaining nutrients in amended tropical soils. J. Plant Nutr. Soil Sci. 2004, 177, 671–680. [Google Scholar] [CrossRef]
- Hernandez-Soriano, M.C.; Kerré, B.; Goos, P.; Hardy, B.; Dufey, J.; Smolders, E. Long-term effect of biochar on the stabilization of recent carbon: Soils with historical inputs of charcoal. GCB Bioenergy 2016, 8, 371–381. [Google Scholar] [CrossRef] [Green Version]
- Burger, M.; Jackson, L.E. Microbial immobilization of ammonium and nitrate in relation to ammonification and nitrification rates in organic and conventional cropping systems. Soil Biol. Biochem. 2003, 35, 29–36. [Google Scholar] [CrossRef]
- Hale, S.E.; Alling, V.; Martinsen, V.; Mulder, J.; Breedveld, G.D.; Cornelissen, G. The sorption and desorption of phosphate-P, ammonium-N and nitrate-N in cacao shell and corn cob biochars. Chemosphere 2013, 91, 1612–1619. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Ni, T.; Li, J.; Lu, Q.; Fang, Z.; Huang, Q.; Zhang, R.; Li, R.; Shen, B.; Shen, Q. Effects of organic–inorganic compound fertilizer with reduced chemical fertilizer application on crop yields, soil biological activity and bacterial community structure in a rice–wheat cropping system. Appl. Soil Ecol. 2016, 99, 1–12. [Google Scholar] [CrossRef]
- Zhao, S.; Wang, B.; Gao, Q.; Gao, Y.; Liu, S. Adsorption of phosphorus by different biochars. Spectr. Lett. 2017, 50, 73–80. [Google Scholar] [CrossRef]
- Farrell, M.; Kuhn, T.K.; Macdonald, L.M.; Maddern, T.M.; Murphy, D.V.; Hall, P.A.; Singh, B.P.; Baumann, K.; Krull, E.S.; Baldock, J.A. Microbial utilisation of biochar-derived carbon. Sci. Total Environ. 2013, 465, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Spohn, M.; Kuzyakov, Y. Distribution of microbial-and root-derived phosphatase activities in the rhizosphere depending on P availability and C allocation–Coupling soil zymography with 14C imaging. Soil Biol. Biochem. 2013, 67, 106–113. [Google Scholar] [CrossRef]
- Chen, C.R.; Condron, L.M.; Davis, M.R.; Sherlock, R.R. Effects of afforestation on phosphorus dynamics and biological properties in a New Zealand grassland soil. Plant Soil. 2000, 220, 151–163. [Google Scholar] [CrossRef]
- Achat, D.L.; Morel, C.; Bakker, M.R.; Augusto, L.; Pellerin, S.; Gallet-Budynek, A.; Gonzalez, M. Assessing turnover of microbial biomass phosphorus: Combination of an isotopic dilution method with a mass balance model. Soil Biol. Biochem. 2010, 42, 2231–2240. [Google Scholar] [CrossRef]
- Latati, M.; Aouiche, A.; Tellah, S.; Laribi, A.; Benlahrech, S.; Kaci, G.; Ouarem, F.; Ounane, S.M. Intercropping maize and common bean enhances microbial carbon and nitrogen availability in low phosphorus soil under Mediterranean conditions. Eur. J. Soil Biol. 2017, 80, 9–18. [Google Scholar] [CrossRef]
- Li, Y.; Wu, J.; Shen, J.; Liu, S.; Wang, C.; Chen, D.; Huang, T.; Zhang, J. Soil microbial C:N ratio is a robust indicator of soil productivity for paddy fields. Sci. Rep. 2016, 6, 35266. [Google Scholar] [CrossRef] [PubMed]
- Zechmeister-Boltenstern, S.; Keiblinger, K.M.; Mooshammer, M.; Peñuelas, J.; Richter, A.; Sardans, J.; Wanek, W. The application of ecological stoichiometry to plant–microbial–soil organic matter transformations. Ecol. Monogr. 2015, 85, 133–155. [Google Scholar] [CrossRef]
- Adetunji, A.T.; Lewu, F.B.; Mulidzi, R.; Ncube, B. The biological activities of β-glucosidase, phosphatase and urease as soil quality indicators: A review. J. Soil Sci. Plant Nutr. 2017, 17, 794–807. [Google Scholar] [CrossRef]
- Abujabhah, I.S.; Bound, S.A.; Doyle, R.; Bowman, J.P. Effects of biochar and compost amendments on soil physico-chemical properties and the total community within a temperate agricultural soil. Appl. Soil Ecol. 2016, 98, 243–253. [Google Scholar] [CrossRef]
- Huang, D.; Liu, L.; Zeng, G.; Xu, P.; Huang, C.; Deng, L.; Wang, R.; Wan, J. The effects of rice straw biochar on indigenous microbial community and enzymes activity in heavy metal-contaminated sediment. Chemosphere 2017, 174, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Gascó, G.; Paz-Ferreiro, J.; Cely, P.; Plaza, C.; Méndez, A. Influence of pig manure and its biochar on soil CO2 emissions and soil enzymes. Ecol. Eng. 2016, 95, 19–24. [Google Scholar] [CrossRef]
- Jaafar, N.M.; Clode, P.L.; Abbott, L.K. Microscopy observations of habitable space in biochar for colonization by fungal hyphae from soil. J. Integr. Agric. 2014, 13, 483–490. [Google Scholar] [CrossRef]
- Cornelissen, G.; Gustafsson, Ö.; Bucheli, T.D.; Jonker, M.T.; Koelmans, A.A.; van Noort, P.C. Extensive sorption of organic compounds to black carbon, coal, and kerogen in sediments and soils: Mechanisms and consequences for distribution, bioaccumulation, and biodegradation. Environ. Sci. Technol. 2005, 39, 6881–6895. [Google Scholar] [CrossRef] [PubMed]
- Warnock, D.D.; Lehman, J.; Kuype, T.W.; Rillig, M.C. Mycorrhizal responses to biochar in soil-concepts and mechanisms. Plant Soil. 2007, 300, 9–20. [Google Scholar] [CrossRef]
- Lu, H.; Lashari, M.S.; Liu, X.; Ji, H.; Li, L.; Zheng, J.; Kibue, G.W.; Joseph, S.; Pan, G. Changes in soil microbial community structure and enzyme activity with amendment of biochar-manure compost and pyroligneous solution in a saline soil from Central China. Eur. J. Soil Biol. 2015, 70, 67–76. [Google Scholar] [CrossRef]
- Noyce, G.L.; Basiliko, N.; Fulthorpe, R.; Sackett, T.E.; Thomas, S.C. Soil microbial responses over 2 years following biochar addition to a north temperate forest. Biol. Fertil. Soils 2015, 51, 649–659. [Google Scholar] [CrossRef]
- Jeffery, S.; Abalos, D.; Prodana, M.; Bastos, A.C.; Van Groenigen, J.W.; Hungate, B.A.; Verheijen, F. Biochar boosts tropical but not temperate crop yields. Environ. Res. Lett. 2017, 12, 053001. [Google Scholar] [CrossRef] [Green Version]
- Singh, H. Nitrogen mineralization, microbial biomass and crop yield as affected by wheat residue placement and fertilizer in a semi-arid tropical soil with minimum tillage. J. Appl. Ecol. 1995, 32, 588–595. [Google Scholar] [CrossRef]
- Olsen, S.R. The role of organic matter and ammonium in producing high corn yields. In The Role of Organic Matter in Modern Agriculture; Chen, Y., Avnimelech, Y., Eds.; Martinus Nijhoff: Dordrecht, The Netherlands, 1986; pp. 29–54. [Google Scholar]
- Yusuf, A.A.; Abaidoo, R.C.; Iwuafor, E.N.O.; Olufajo, O.O.; Sanginga, N. Rotation effects of grain legumes and fallow on maize yield, microbial biomass and chemical properties of an Alfisol in the Nigerian savanna. Agric. Ecosyst. Environ. 2009, 129, 325–331. [Google Scholar] [CrossRef]


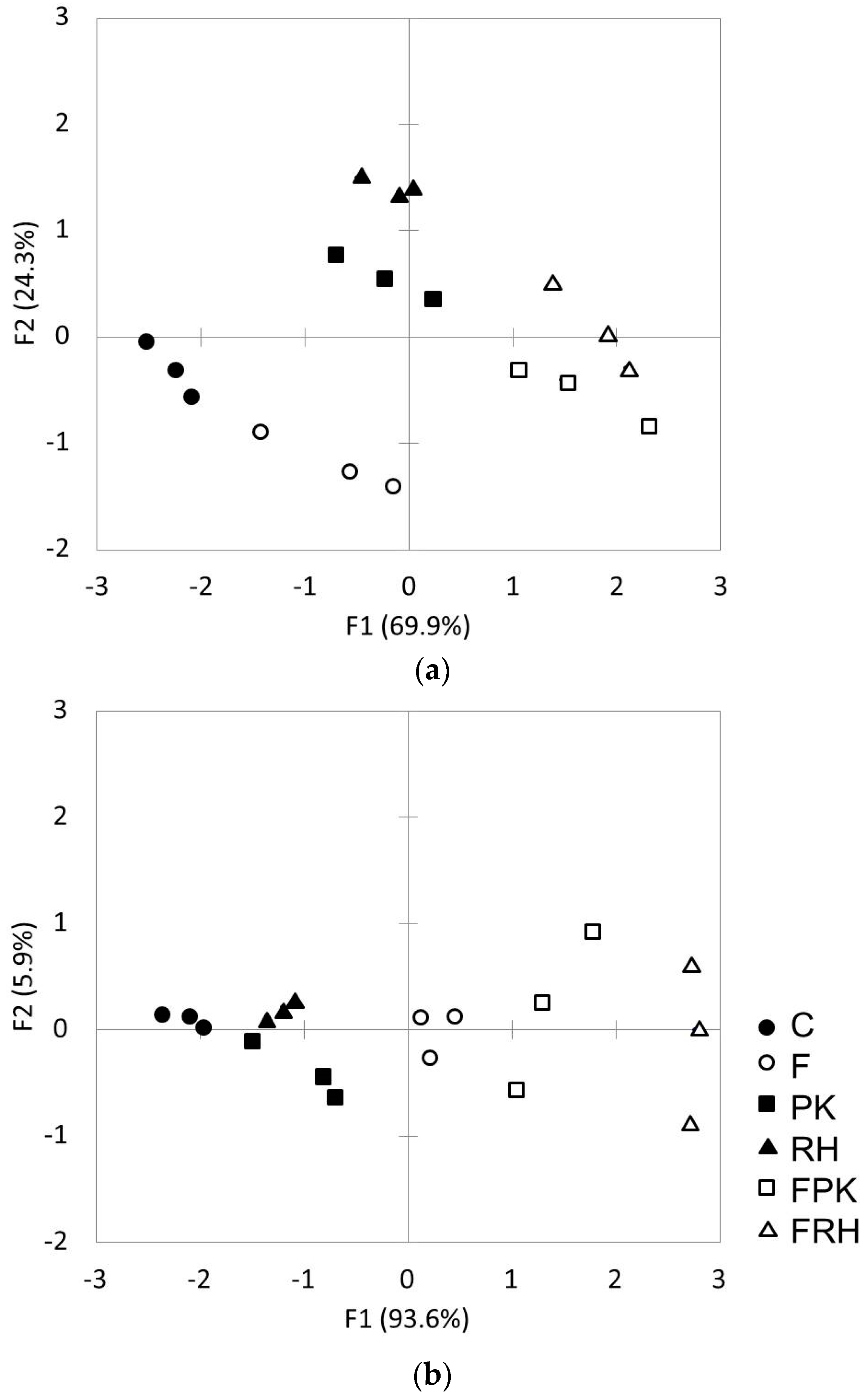{kind=link}
| Properties | Palm Kernel Shell Biochar | Rice Husk Biochar |
|---|---|---|
| Production Process | Slow Pyrolysis at 400 °C | Gasification at 800 °C |
| pH | 8.67 | 10.24 |
| Total C (g kg−1) | 434.1 | 101.7 |
| Total N (g kg−1) | 5.07 | 2.52 |
| Total P (g kg−1) | 1.56 | 2.11 |
| C:N | 85.62 | 40.37 |
| NH4+—N (mg kg−1) | 21.05 | 25.48 |
| NO3−—N (mg kg−1) | 40.88 | 92.17 |
| Ash (g kg−1) | 180 | 750 |
| BET surface area (m2 g−1) | 184.2 | 1.6 |
| Particle size range (mm) | 5–10 | 1–3 |
| Treatment | pH | Cation Exchange Capacity (CEC) (cmolc kg−1 soil) | Total | Available | ||||
|---|---|---|---|---|---|---|---|---|
| Organic C (g kg−1 soil) | N (g kg−1 soil) | P (mg kg−1 soil) | NH4+—N (mg kg−1 soil) | NO3—N (mg kg−1 soil) | P (mg kg−1 soil) | |||
| C | 4.97 b | 4.11 b | 5.96 b | 0.61 b | 189.0 ab | 9.86 b | 20.93 b | 37.33 b |
| F | 5.03 b | 4.21 b | 5.83 b | 0.83 a | 147.6 ab | 10.52 b | 21.74 b | 46.33 b |
| PK | 6.31 a | 4.94 a | 8.39 a | 0.77 ab | 130.9 b | 10.03 b | 21.04 b | 42.64 b |
| RH | 6.21 a | 4.88 a | 9.41 a | 0.93 a | 134.4 ab | 10.15 b | 35.11 a | 40.73 b |
| FPK | 6.18 a | 4.93 a | 8.85 a | 0.91 a | 238.2 a | 12.31 a | 22.33 b | 65.26 a |
| FRH | 6.14 a | 4.89 a | 9.19 a | 0.87 a | 145.7 ab | 12.88 a | 38.42 a | 42.53 b |
| Treatment | Microbial Biomass | Enzyme Activity | Microbial Gene Abundance | |||||
|---|---|---|---|---|---|---|---|---|
| C (mg kg−1 Soil) | N (mg kg−1 Soil) | P (mg kg−1 Soil) | β-glucosidase (μmol pNP g −1 Soil h−1) | Urease (μmol NH3 g−1 Soil h−1) | Phosphodiesterase (μmol pNP −1g Soil h−1) | Bacterial 16S rRNA (log10 Gene Copies g−1 Soil) | Fungal ITS (log10 Gene Copies g−1 Soil) | |
| C | 97.1 c | 8.33 c | 0.79 b | 228.3 b | 2.46 c | 95.1 b | 6.35 b | 5.46 b |
| F | 93.9 c | 26.67 b | 0.85 b | 222.2 b | 6.93 a | 167.5 a | 8.23 a | 6.03 b |
| PK | 140.5 b | 23.33 b | 0.82 b | 295.9 a | 5.62 b | 152.9 a | 8.57 a | 7.62 a |
| RH | 160.9 a | 21.68 b | 0.81 b | 287.1 a | 5.24 b | 165.3 a | 9.78 a | 7.54 a |
| FPK | 137.2 b | 38.34 a | 0.91 a | 256.4 a | 7.24 a | 181.7 a | 9.11 a | 7.23 a |
| FRH | 154.8 a | 36.71 a | 0.84 b | 265.1 a | 7.47 a | 175.2 a | 9.02 a | 7.42 a |
| Treatment | C:N | C:P | N:P |
|---|---|---|---|
| Soil | |||
| C | 9.77 b | 31.52 b | 3.22 b |
| F | 7.02 c | 39.49 b | 5.62 a |
| PK | 10.89 a | 64.49 a | 5.92 a |
| RH | 10.12 ab | 70.01 a | 6.91 a |
| FPK | 9.73 b | 37.16 b | 3.82 b |
| FRH | 10.56 a | 63.34 a | 5.99 a |
| Microbial biomass | |||
| C | 11.65 a | 122.92 bc | 10.54 c |
| F | 3.52 c | 110.52 c | 31.37 ab |
| PK | 6.02 b | 171.35 b | 28.45 b |
| RH | 7.43 b | 198.64 a | 26.76 b |
| FPK | 3.58 c | 150.78 bc | 40.13 a |
| FRH | 4.22 c | 173.65 a | 41.05 a |
| Treatment | Uptake | Biomass (t h−1) | |
|---|---|---|---|
| N (t ha−1) | P (kg ha−1) | ||
| C | 8.01 c | 28.87 e | 1.97 d |
| F | 60.34 bc | 102.77 c | 6.56 b |
| PK | 19.36 c | 77.42 cd | 4.41 c |
| RH | 32.39 c | 51.61 ed | 3.84 c |
| FPK | 94.69 ab | 144.71 b | 7.57 b |
| FRH | 115.01 a | 194.62 a | 10.25 a |
| Parameter | Biomass |
|---|---|
| Soil | |
| C | 0.4016 |
| N | 0.5998 |
| P | 0.0705 |
| C:N | −0.0443 |
| C:P | 0.1624 |
| N:P | 0.2204 |
| Microbial biomass | |
| C | 0.3449 |
| N | 0.9186 ** |
| P | 0.8105 |
| C:N | −0.8297 * |
| C:P | 0.2015 |
| N:P | 0.8061 |
| Enzyme activity | |
| β-glucosidase | 0.0131 |
| Urease | 0.8174 |
| Phosphodiesterase | 0.7686 |
| Microbial gene abundance | |
| 16S rRNA | 0.5035 |
| ITS | 0.4075 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azlan Halmi, M.F.; Hasenan, S.N.; Simarani, K.; Abdullah, R. Linking Soil Microbial Properties with Plant Performance in Acidic Tropical Soil Amended with Biochar. Agronomy 2018, 8, 255. https://doi.org/10.3390/agronomy8110255
Azlan Halmi MF, Hasenan SN, Simarani K, Abdullah R. Linking Soil Microbial Properties with Plant Performance in Acidic Tropical Soil Amended with Biochar. Agronomy. 2018; 8(11):255. https://doi.org/10.3390/agronomy8110255
Chicago/Turabian StyleAzlan Halmi, Muhammad Farid, Siti Nahdiyyah Hasenan, Khanom Simarani, and Rosazlin Abdullah. 2018. "Linking Soil Microbial Properties with Plant Performance in Acidic Tropical Soil Amended with Biochar" Agronomy 8, no. 11: 255. https://doi.org/10.3390/agronomy8110255
APA StyleAzlan Halmi, M. F., Hasenan, S. N., Simarani, K., & Abdullah, R. (2018). Linking Soil Microbial Properties with Plant Performance in Acidic Tropical Soil Amended with Biochar. Agronomy, 8(11), 255. https://doi.org/10.3390/agronomy8110255