Variation of Fine Roots Distribution in Apple (Malus pumila M.)–Crop Intercropping Systems on the Loess Plateau of China

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
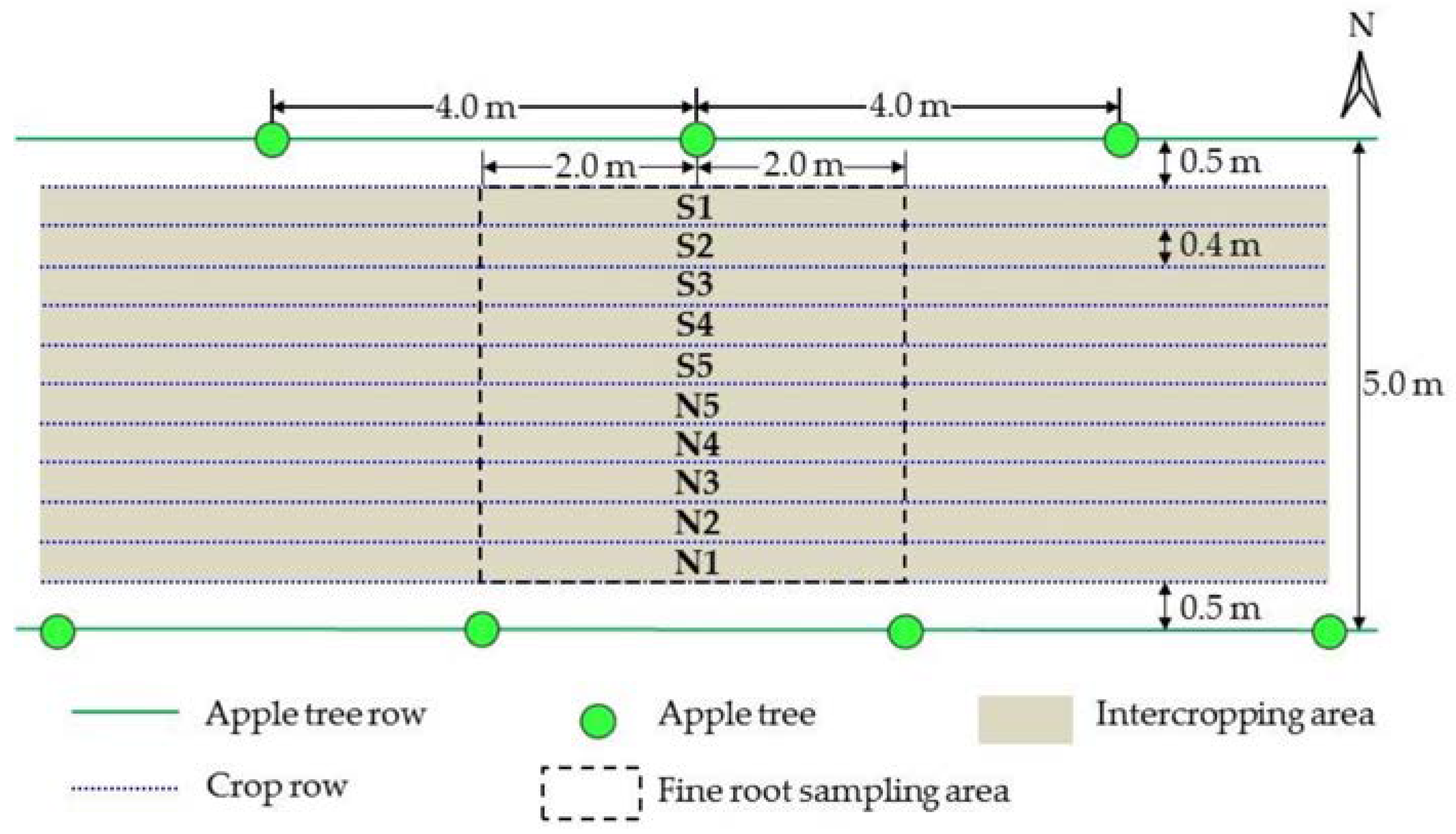
2.2. Materials and Experimental Design
2.3. Collection and Measurement of Root Samples
2.4. Fine-Root Biomass Density
2.5. Fine-Root Vertical Distribution
2.6. Fine-Root Horizontal Distribution
2.7. Statistical Analyses
3. Results
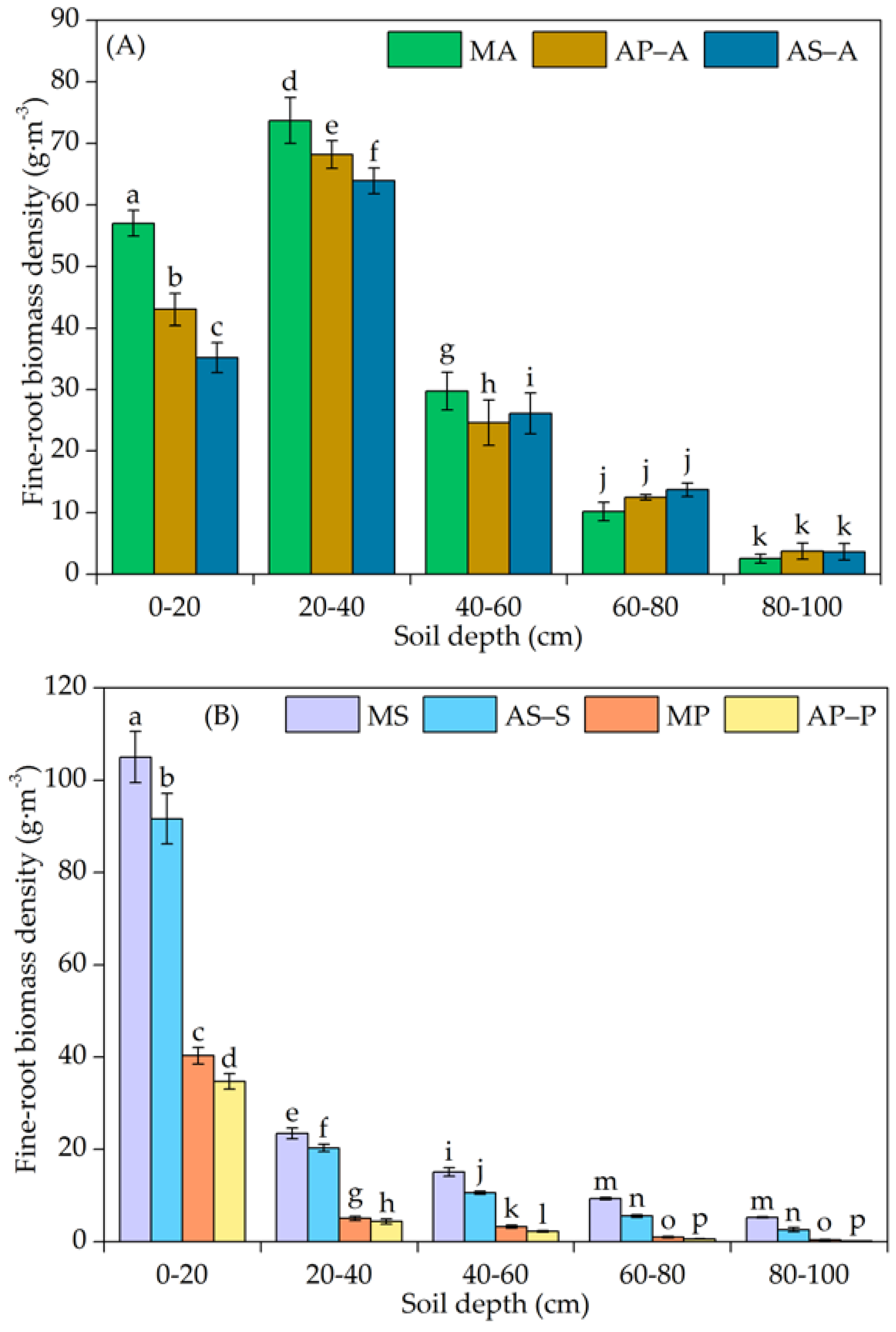
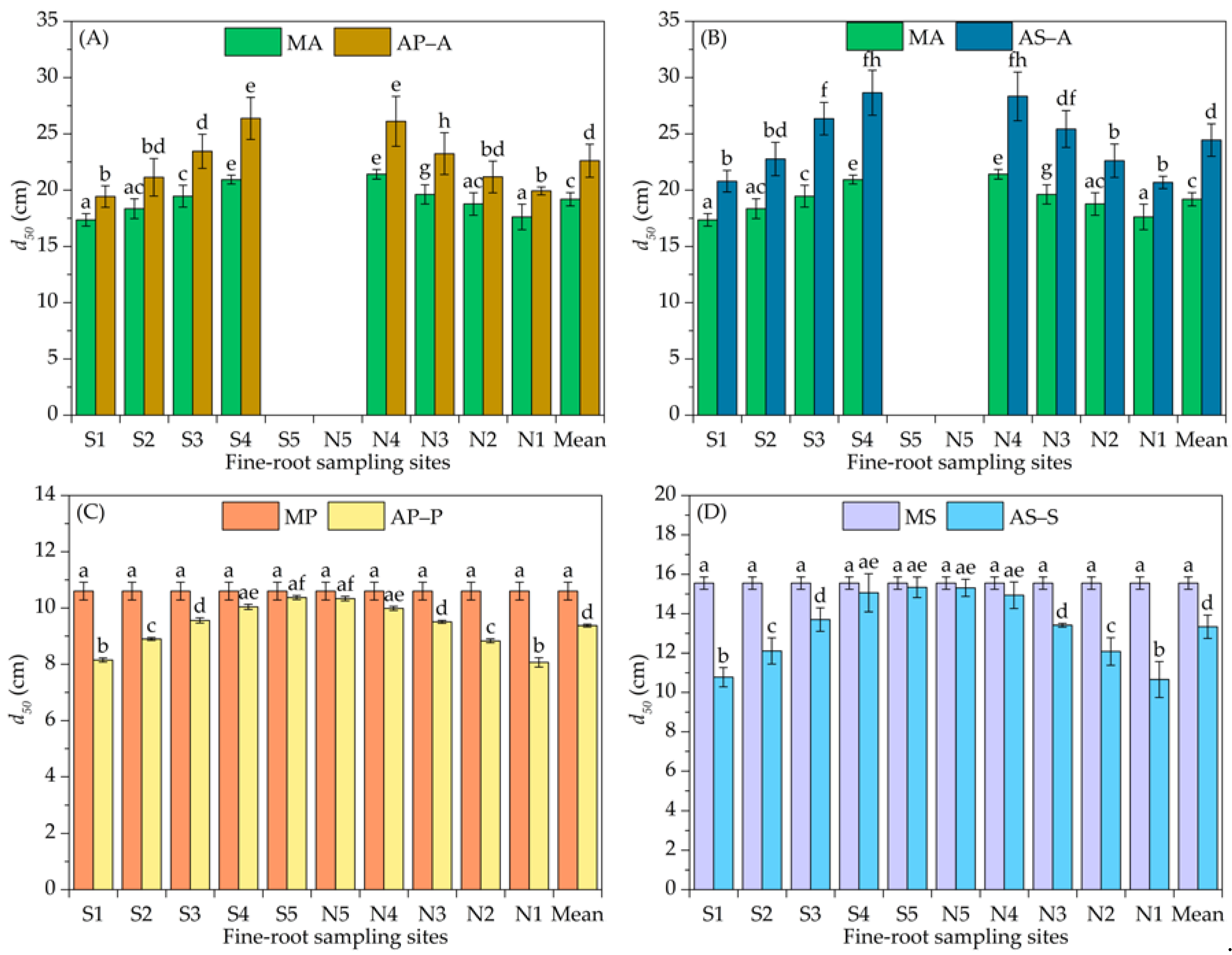
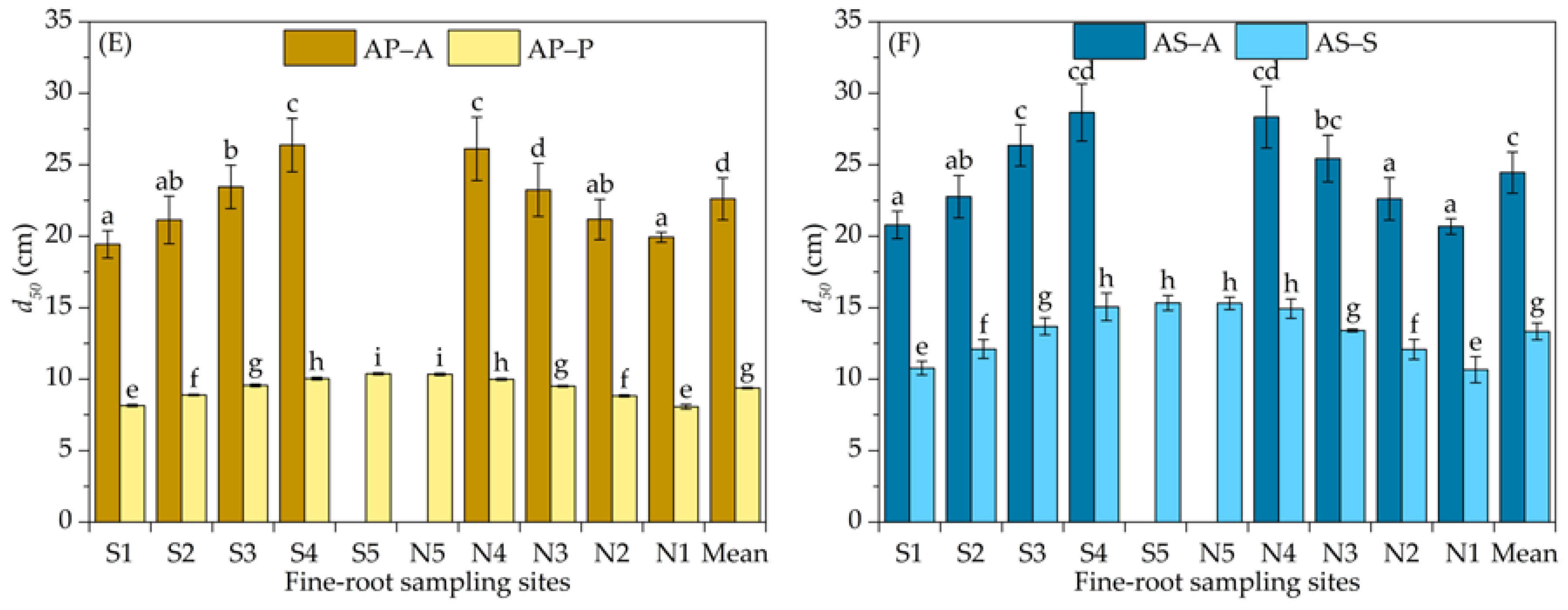
3.1. Vertical Distribution of the Fine-Root Biomass Density
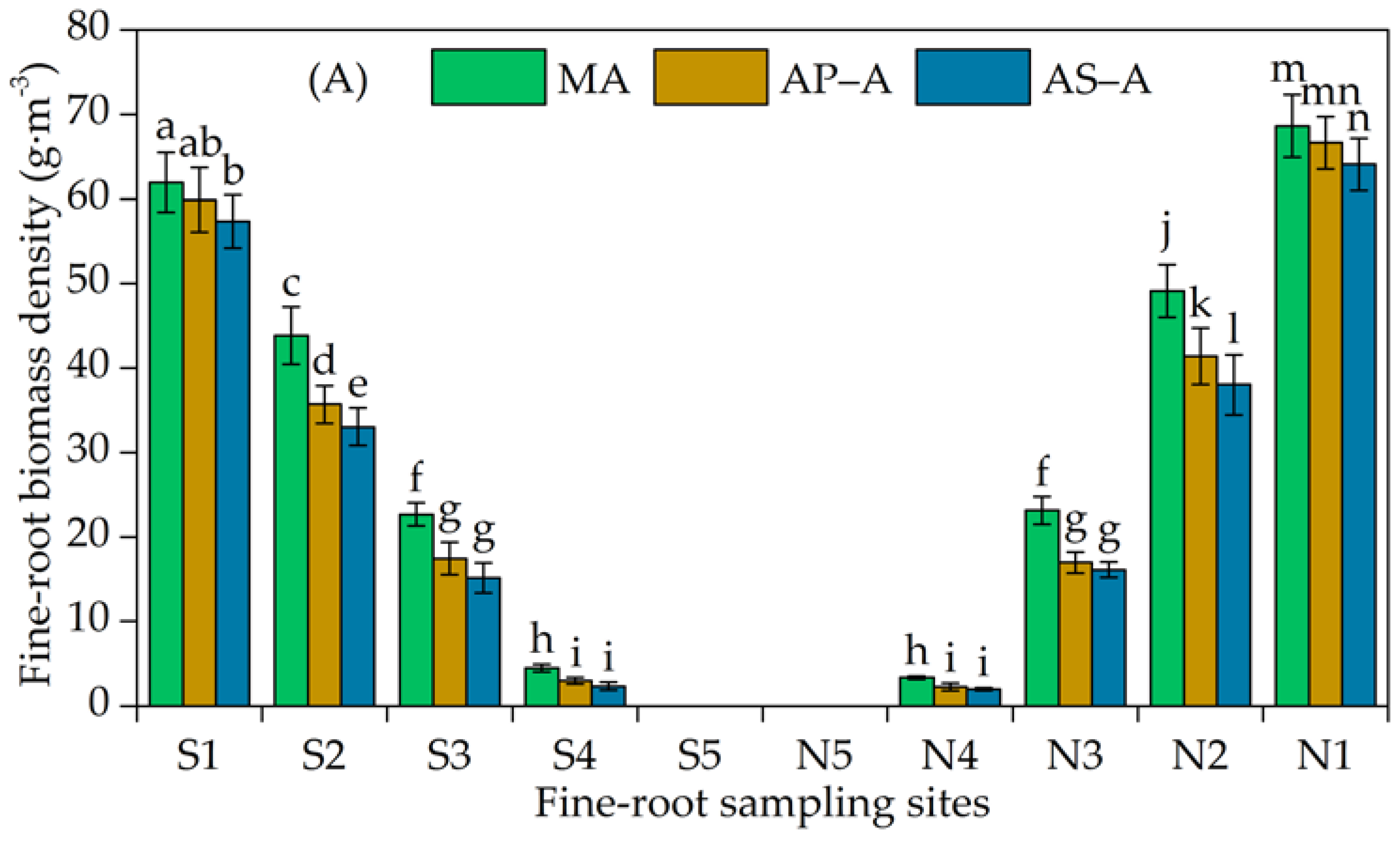
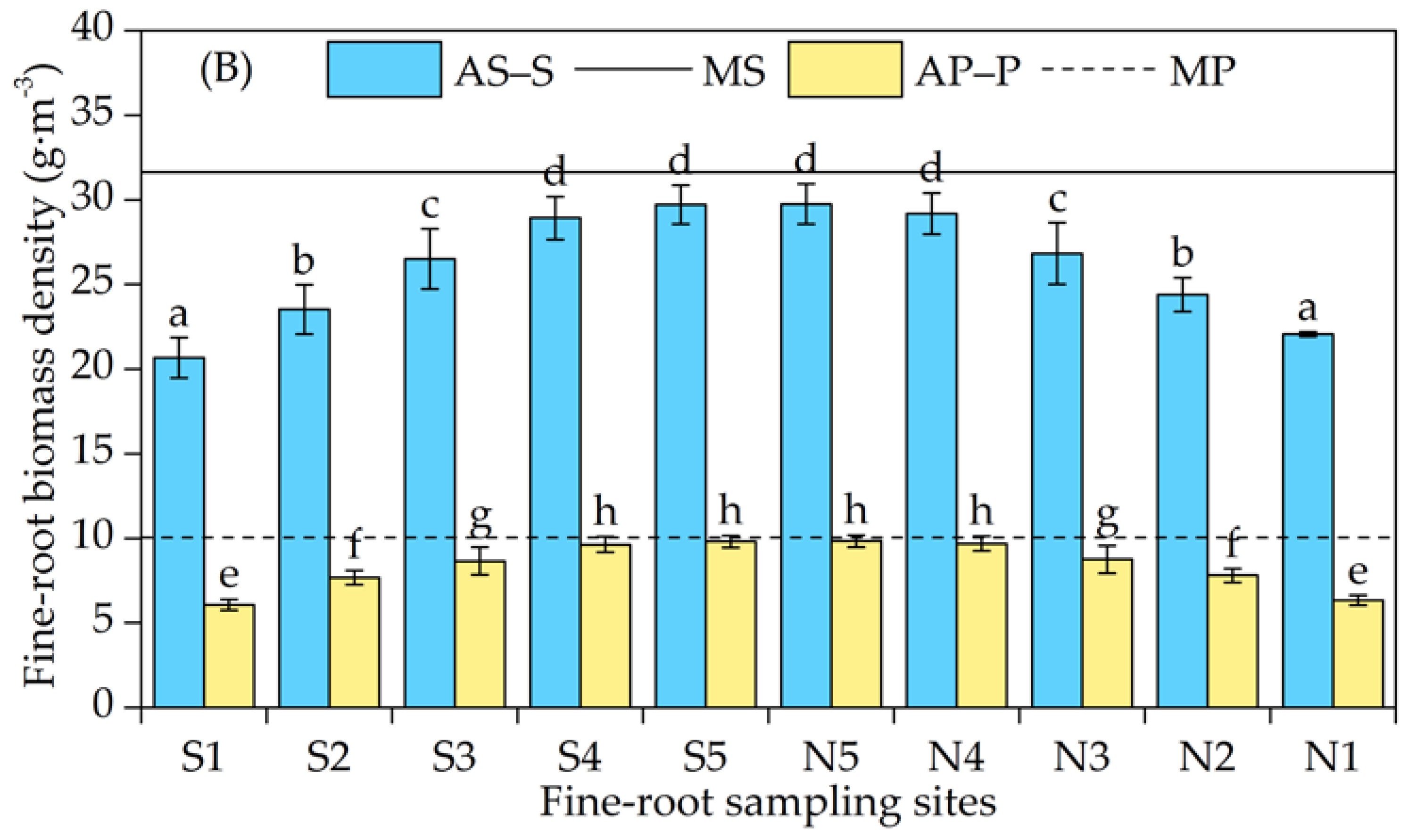
3.2. Horizontal Distribution of the Fine-Root Biomass Density
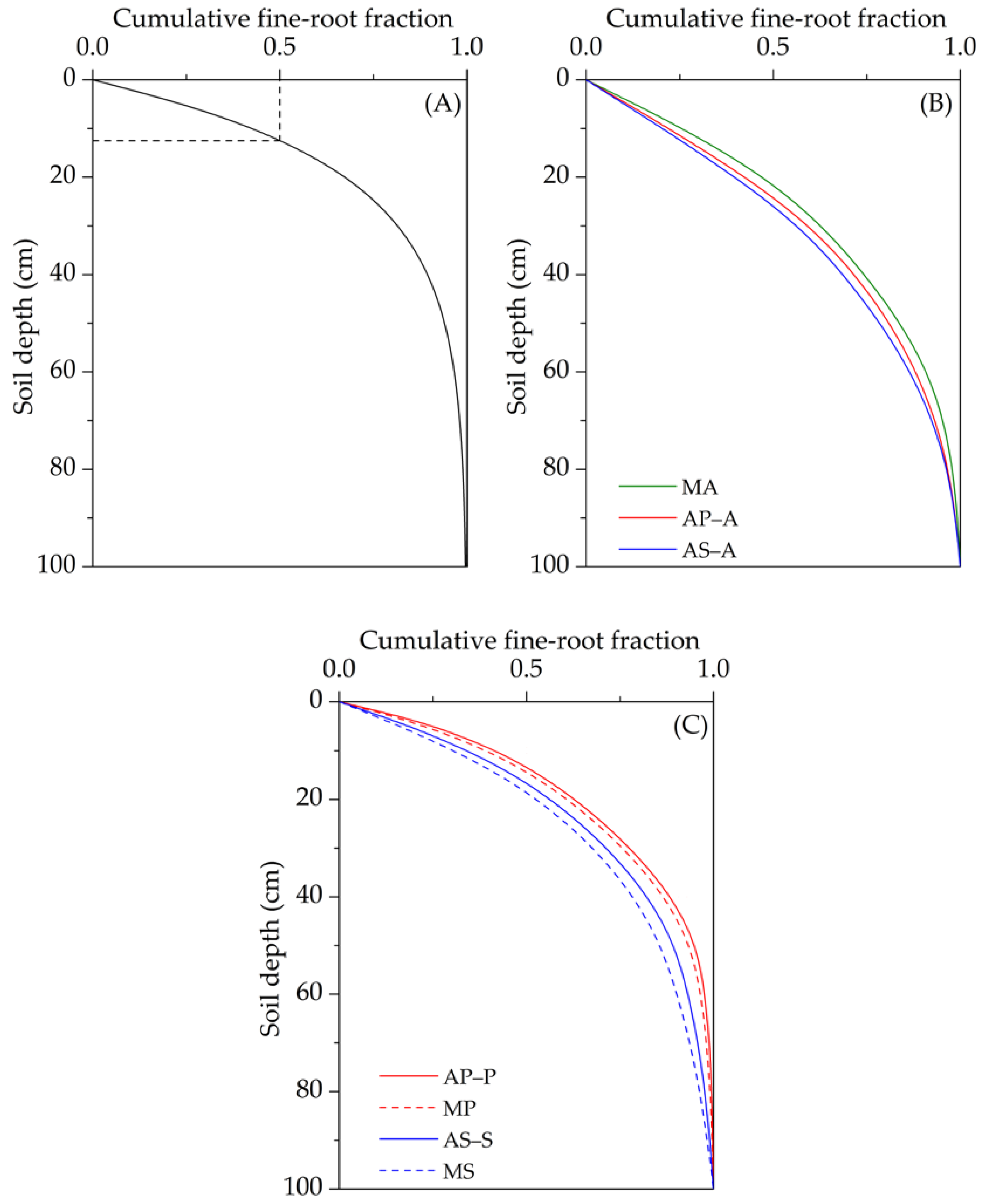
3.3. Vertical Distribution of Fine Roots
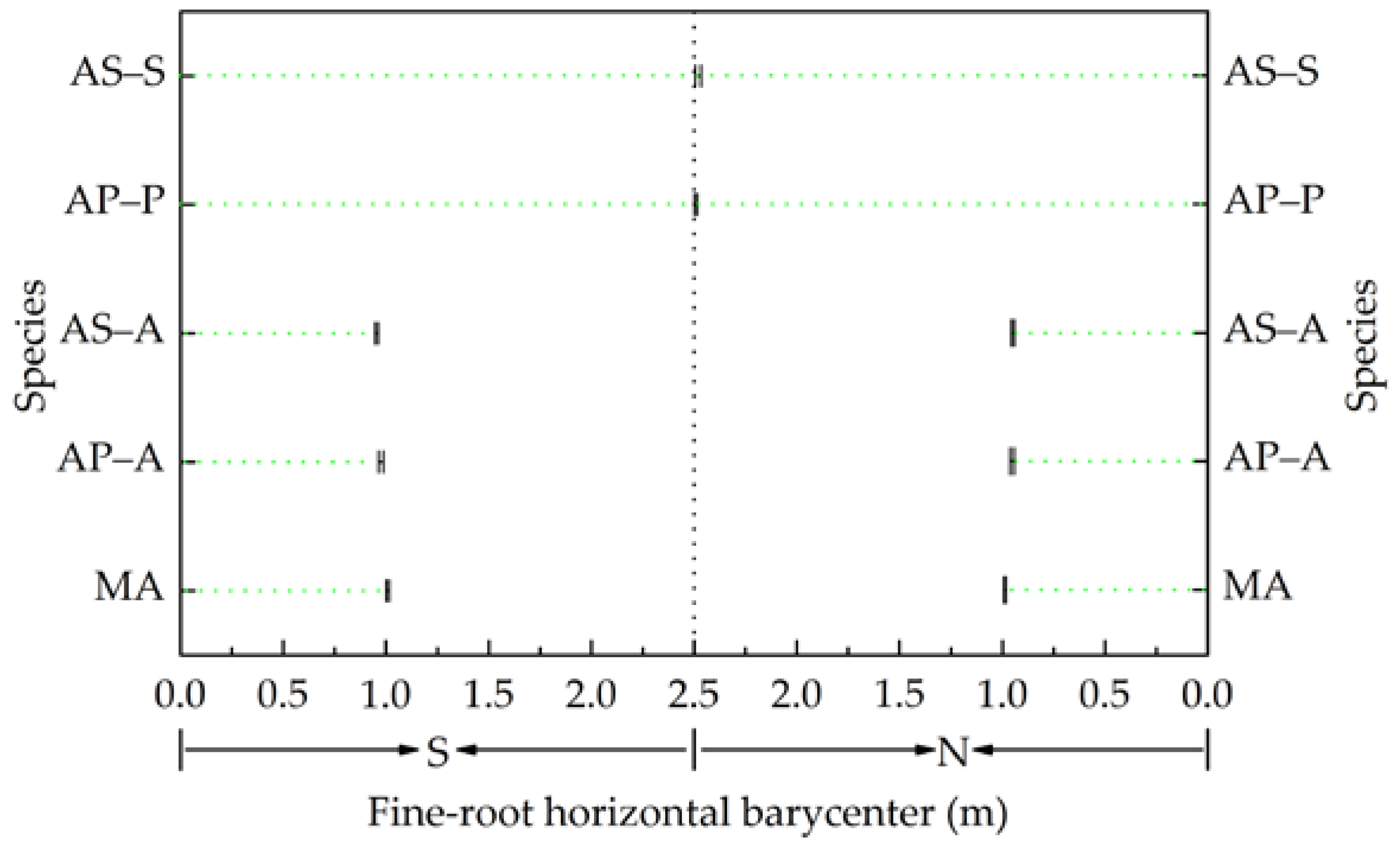
3.4. Horizontal Distribution of Fine Roots
4. Discussion
4.1. Variation in the Vertical Distribution of Fine Roots
4.2. Variation in the Horizontal Distribution of Fine Roots
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Meng, P.; Zhang, J.S.; Fan, W. Research on Agroforestry in China; Chinese Forestry Press: Beijing, China, 2003; pp. 1–4. ISBN 7503835842. [Google Scholar]
- Rao, M.R.; Nair, P.K.R.; Ong, C.K. Biophysical interactions in tropical agroforestry systems. Agrofor. Syst. 1998, 38, 3–50. [Google Scholar] [CrossRef]
- Cai, C.F.; Wang, F.; Ding, S.W.; Huang, L.; Shi, Z.H. Nutrients competition and its action mechanism between component parts in inter-cropping systems and agroforestry. Res. Soil Water Conserv. 2000, 7, 219–221. [Google Scholar]
- Thevathasan, N.V.; Gordon, A.M. Ecology of tree intercropping systems in the North temperate region: Experiences from southern Ontario, Canada. Agrofor. Syst. 2004, 61, 257–268. [Google Scholar] [CrossRef]
- Wu, G.; Li, J.; Deng, H.B. Primary research on interface ecology in agroforestrial ecosystems. Chin. J. Appl. Ecol. 2000, 11, 459–460. [Google Scholar]
- Gao, L.; Xu, H.; Bi, H.; Xi, W.; Bao, B.; Wang, X.; Bi, C.; Chang, Y. Intercropping competition between apple trees and crops in agroforestry systems on the Loess Plateau of China. PLoS ONE 2013, 8, e70739. [Google Scholar] [CrossRef] [PubMed]
- Monteith, J.L.; Ong, C.K.; Corlett, J.E. Microclimatic interactions in agroforestry systems. For. Ecol. Manag. 1991, 45, 31–44. [Google Scholar] [CrossRef]
- Ong, C.K.; Corlett, J.E.; Singh, R.P. Above and below ground interaction in agroforestry systems. For. Ecol. Manag. 1991, 45, 45–57. [Google Scholar] [CrossRef]
- Kowalchuk, T.E.; de Jong, E. Shelterbelts and their effect on crop yield. Can. J. Soil Sci. 1995, 75, 543–550. [Google Scholar] [Green Version]
- McIntyre, B.D.; Riha, S.J.; Ong, C.K. Competition for water in a hedge-intercrop system. Field Crop. Res. 1997, 52, 151–160. [Google Scholar] [CrossRef]
- Smith, D.M.; Jackson, N.A.; Roberts, J.M.; Ong, C.K. Root distributions in a Grevillea robusta-maize agroforestry system in semi-arid Kenya. Plant Soil 1999, 211, 191–205. [Google Scholar] [CrossRef]
- Jose, S.; Gillespie, A.R.; Pallardy, S.G. Interspecific interactions in temperate agroforestry. Agrofor. Syst. 2004, 61, 237–255. [Google Scholar] [CrossRef]
- Jose, S.; Williams, R.A.; Zamora, D.S. Belowground ecological interactions in mixed-species forest plantations. For Ecol. Manag. 2006, 233, 231–239. [Google Scholar] [CrossRef]
- Smucker, A.J.M.; Aiken, R.M. Dynamic Root Responses to Water Deficits. Soil Sci. 1992, 154, 281–289. [Google Scholar] [CrossRef]
- De Kroon, H.; Mommer, L.; Nishiwaki, A. Root competition: Towards a mechanistic understanding. In Root Ecology; de Kroon, H., Visser, E.J.W., Eds.; Springer: Berlin, Germany, 2003; pp. 215–234. ISBN 9783642055201. [Google Scholar]
- Day, K.J.; John, E.A.; Hutchings, M.J. The effects of spatially heterogeneous nutrient supply on yield, intensity of competition and root placement patterns in Briza media and Festuca ovina. Funct. Ecol. 2003, 17, 454–463. [Google Scholar] [CrossRef]
- Tufekcioglu, A.; Raich, J.W.; Isenhart, T.M.; Schultz, R.C. Fine root dynamics, coarse root biomass, root distribution, and soil respiration in a multispecies riparian buffer in Central Iowa, USA. Agrofor. Syst. 1999, 44, 163–174. [Google Scholar] [CrossRef]
- Schenk, H.J.; Jackson, R.B. The global biogeography of roots. Ecol. Monogr. 2002, 72, 311–328. [Google Scholar] [CrossRef]
- Zhang, Y.Q.; Zhu, Q.K.; Qi, S.; Zhang, Y.; Wang, D.M. Root system distribution characteristics of plants on the terrace banks and their impact on soil moisture. Acta Ecol. Sin. 2005, 25, 500–506. [Google Scholar]
- Kumar, S.S.; Kumar, B.M.; Wahid, P.A.; Kamalam, N.V.; Fisher, R.F. Root competition for phosphorus between coconut, multipurpose trees and kacholam (Kaempferia galanga L.) in Kerala, India. Agrofor. Syst. 1999, 46, 131–146. [Google Scholar] [CrossRef]
- Hinsinger, P.; Bengough, A.G.; Vetterlein, D.; Young, I.M. Rhizosphere: Biophysics, biogeochemistry and ecological relevance. Plant Soil 2009, 321, 117–152. [Google Scholar] [CrossRef]
- Cahill, J.F.; McNickle, G.G.; Haag, J.J.; Lamb, E.G.; Nyanumba, S.M.; Clair, C.C.S. Plants integrate information about nutrients and neighbors. Science 2010, 328, 1657–1666. [Google Scholar] [CrossRef] [PubMed]
- Bi, H.X.; Yun, L.; Zhu, Q.K. Study on the Interspecific Relationships of Agroforestry Systems in the Loess Area of Western Shanxi Province; Science Press: Beijing, China, 2011; pp. 20–109. ISBN 9787030313669. [Google Scholar]
- Gregory, P.J. Approaches to modeling the uptake of water and nutrients in agroforestry systems. Agrofor. Syst. 1996, 34, 51–65. [Google Scholar] [CrossRef]
- Van Noordwijk, M.; Purnomosidhi, P. Root architecture in relation to tree-soil-crop interactions and shoot pruning in agroforestry. Agrofor. Syst. 1995, 30, 161–173. [Google Scholar] [CrossRef]
- Livesley, S.J.; Gregory, P.J.; Buresh, R.J. Competition in tree row agroforestry systems. 1. Distribution and dynamics of fine roots length and biomass. Plant Soil 2000, 227, 149–161. [Google Scholar] [CrossRef]
- Xu, H.S.; Bi, H.X.; Xi, W.M.; Powell, R.L.; Gao, L.B.; Yun, L. Root distribution variation of crops under walnut-based intercropping systems in the Loess Plateau of China. Pak. J. Agri. Sci. 2014, 51, 773–778. [Google Scholar]
- Fan, W.; Lu, Q.; Gao, X.R. Distribution pattern and growing dynamics of the roots system in apple-wheat intercropping system. Acta Ecol. Sin. 1999, 19, 860–863. [Google Scholar]
- Meng, P.; Zhang, J.S.; Yin, C.J.; Ma, X.Q.; Feng, W.D. Experiments on characteristics of wheat roots difference between apple-wheat intercropping system and wheat monoculture system. For. Res. 2002, 15, 369–373. [Google Scholar]
- Xu, H.S.; Bi, H.X.; Gao, L.B.; Yun, L.; Chang, Y.F.; Xi, W.M.; Liao, W.C.; Bao, B. Distribution and morphological variation of fine root in a walnut-soybean intercropping system in the Loess plateau of China. Int. J. Agric. Biol. 2013, 15, 998–1002. [Google Scholar]
- Huxley, P.A.; Pinney, A.; Akunda, E.; Muraya, P. A tree/crop interface orientation experiment with a Grevillea robusta, hedgerow and maize. Agofor. Syst. 1994, 26, 23–45. [Google Scholar] [CrossRef]
- Mulia, R.; Dupraz, C. Unusual fine root distributions of two deciduous tree species in southern France: What consequences for modelling of tree root dynamics? Plant Soil 2006, 281, 71–85. [Google Scholar] [CrossRef]
- Cardinael, R.; Mao, Z.; Prieto, I.; Stokes, A.; Dupraz, C.; Kim, J.H.; Jourdan, C. Competition with winter crops induces deeper rooting of walnut trees in a Mediterranean alley cropping agroforestry system. Plant Soil 2015, 391, 219–235. [Google Scholar] [CrossRef]
- Zhang, W.; Ahanbieke, P.; Wang, B.J.; Xu, W.L.; Li, L.H.; Christie, P.; Li, L. Root distribution and interactions in jujube tree/wheat agroforestry system. Agrofor. Syst. 2013, 87, 929–939. [Google Scholar] [CrossRef]
- Ma, C.M.; Zhai, M.P.; Liu, C.P. Root distribution characteristics of Juglans regia in monoculture and intercropping. J. Beijing For. Univ. 2009, 31, 181–186. [Google Scholar]
- Duan, Z.P.; Gan, Y.W.; Wang, B.J.; Hao, X.D.; Xu, W.L.; Zhang, W.; Li, L.H. Interspecific interaction alters root morphology in young walnut/wheat agroforestry systems in northwest China. Agrofor. Syst. 2017, 1–16. [Google Scholar] [CrossRef]
- Zhang, J.S.; Meng, P.; Yin, C.J. Spatial distribution characteristics of apple tree roots in the apple-wheat intercropping. Sci. Silvae Sin. 2002, 38, 30–33. [Google Scholar]
- Yun, L.; Bi, H.X.; Gao, L.B.; Zhu, Q.K.; Ma, W.J.; Cui, Z.W.; Wilcox, B.P. Soil moisture and soil nutrient content in walnut-crop intercropping systems in the Loess Plateau of China. Arid Land Res. Manag. 2012, 26, 285–296. [Google Scholar] [CrossRef]
- Gale, M.R.; Grigal, D.F. Vertical root distributions of northern tree species in relation to successional status. Can. J. For. Res. 1987, 17, 829–834. [Google Scholar] [CrossRef]
- Casper, B.B.; Jackson, R.B. Plant competition underground. Annu. Rev. Ecol. Syst. 1997, 28, 545–570. [Google Scholar] [CrossRef]
- Zhu, Q.K.; Zhu, J.Z. Sustainable Management Technology for Conversion of Cropland to Forest in Loess Area; Chinese Forestry Press: Beijing, China, 2003; pp. 179–186. ISBN 7503828455. [Google Scholar]
- Kasperbauer, M.J.; Busscher, W.J. Genotypic differences in cotton root penetration of a compacted sub-soil layer. Crop Sci. 1991, 31, 1376–1378. [Google Scholar] [CrossRef]
- Ren, Y.Z.; Xu, Y.H.; Ding, J.P.; Ma, Y.S.; Pei, D.L.; Li, C.W.; Tong, Y.P. Regulation of abiotic factors on the plasticity of plant root development. Chin. Agric. Bull. 2011, 27, 34–38. [Google Scholar]
- Isaac, M.E.; Anglaaere, L.C.N.; Borden, K.; Adu-Bredu, S. Intraspecific root plasticity in agroforestry systems across edaphic conditions. Agric. Ecosyst. Environ. 2014, 185, 16–23. [Google Scholar] [CrossRef]
- McIntyre, B.D.; Riha, S.J.; Ong, C.K. Light interception and evapotranspiration in hedgerow agroforestry systems. Agric. For. Meteorol. 1996, 81, 31–40. [Google Scholar] [CrossRef]
- May, R.; McLean, A.R. Interspecific competition and multispecies coexistence. In Theoretical Ecology: Principles and Applications, 3rd ed.; McLean, A.R., Ed.; Oxford University Press: New York, NY, USA, 2007; pp. 84–97. ISBN 9780199209996. [Google Scholar]
- Yun, L.; Bi, H.X.; Ren, Y.; Wu, J.; Chen, P.P.; Ma, W.J. Research on soil moisture relations among types of agroforestry system in the Loess Region. Bull. Soil Water Conserv. 2008, 28, 110–114. [Google Scholar]
- Eastham, J.; Rose, C.W. Tree/pasture interactions at a range of tree densities in an agroforestry experiment. 1. Rooting patterns. Aust. J. Agric. Res. 1990, 41, 683–695. [Google Scholar] [CrossRef]
- Schroth, G.; Zech, W. Root length dynamics in agroforestry with Gliricidia sepium as compared to sole cropping in the semi-deciduous rainforest zone of West Africa. Plant Soil 1995, 170, 297–306. [Google Scholar] [CrossRef]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Bi, H.; Xu, H.; Duan, H.; Peng, R.; Wang, J. Variation of Fine Roots Distribution in Apple (Malus pumila M.)–Crop Intercropping Systems on the Loess Plateau of China. Agronomy 2018, 8, 280. https://doi.org/10.3390/agronomy8120280
Sun Y, Bi H, Xu H, Duan H, Peng R, Wang J. Variation of Fine Roots Distribution in Apple (Malus pumila M.)–Crop Intercropping Systems on the Loess Plateau of China. Agronomy. 2018; 8(12):280. https://doi.org/10.3390/agronomy8120280
Chicago/Turabian StyleSun, Yubo, Huaxing Bi, Huasen Xu, Hangqi Duan, Ruidong Peng, and Jingjing Wang. 2018. "Variation of Fine Roots Distribution in Apple (Malus pumila M.)–Crop Intercropping Systems on the Loess Plateau of China" Agronomy 8, no. 12: 280. https://doi.org/10.3390/agronomy8120280
APA StyleSun, Y., Bi, H., Xu, H., Duan, H., Peng, R., & Wang, J. (2018). Variation of Fine Roots Distribution in Apple (Malus pumila M.)–Crop Intercropping Systems on the Loess Plateau of China. Agronomy, 8(12), 280. https://doi.org/10.3390/agronomy8120280