The Performance and Potentiality of Monoecious Hemp (Cannabis sativa L.) Cultivars as a Multipurpose Crop

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Site and Field Experiment Description
2.3. Plot Structure and Sampling Scheme
- (1)
- A 1.0 m2 area was randomly selected in each unit. This was identified immediately after sowing, and the number of plantlets until full emergence and at harvest were counted. The emergence date for each genotype was set when 75% of seedlings had emerged.
- (2)
- Fifteen representative plants were randomly selected in the 25 m2 plot to determine phenological stages [20] by twice-weekly surveys.
- (3)
- An area of 1.8 m2 for each experimental unit, discarding the border rows, was utilized to collect the inflorescences at the full flowering stage [20]. The fresh inflorescences were collected after manually cutting the upper part of the plant stem at the insertion level of the first basal flowers of the inflorescence. The fresh inflorescences were directly utilized to obtain the essential oil by distillation. The same plants, after the development of new inflorescences, were harvested at maturity to measure seed yield.
- (4)
- A 4 m2 area, discarding the border rows, was harvested to obtain the crop yield, both as biomass (fiber) and seeds. Plants were cut manually, at 10 cm from the soil surface. Plant height was measured from the base to the top of the inflorescence. Harvesting was performed at the seed maturity stage [20].
2.4. Determination of Biomass, Seed and Seed Oil Yield
2.5. Inflorescence Yield and Essential Oil Content Determination
2.6. Statistical Analysis
3. Results and Discussion
3.1. Climate during the Experimentation
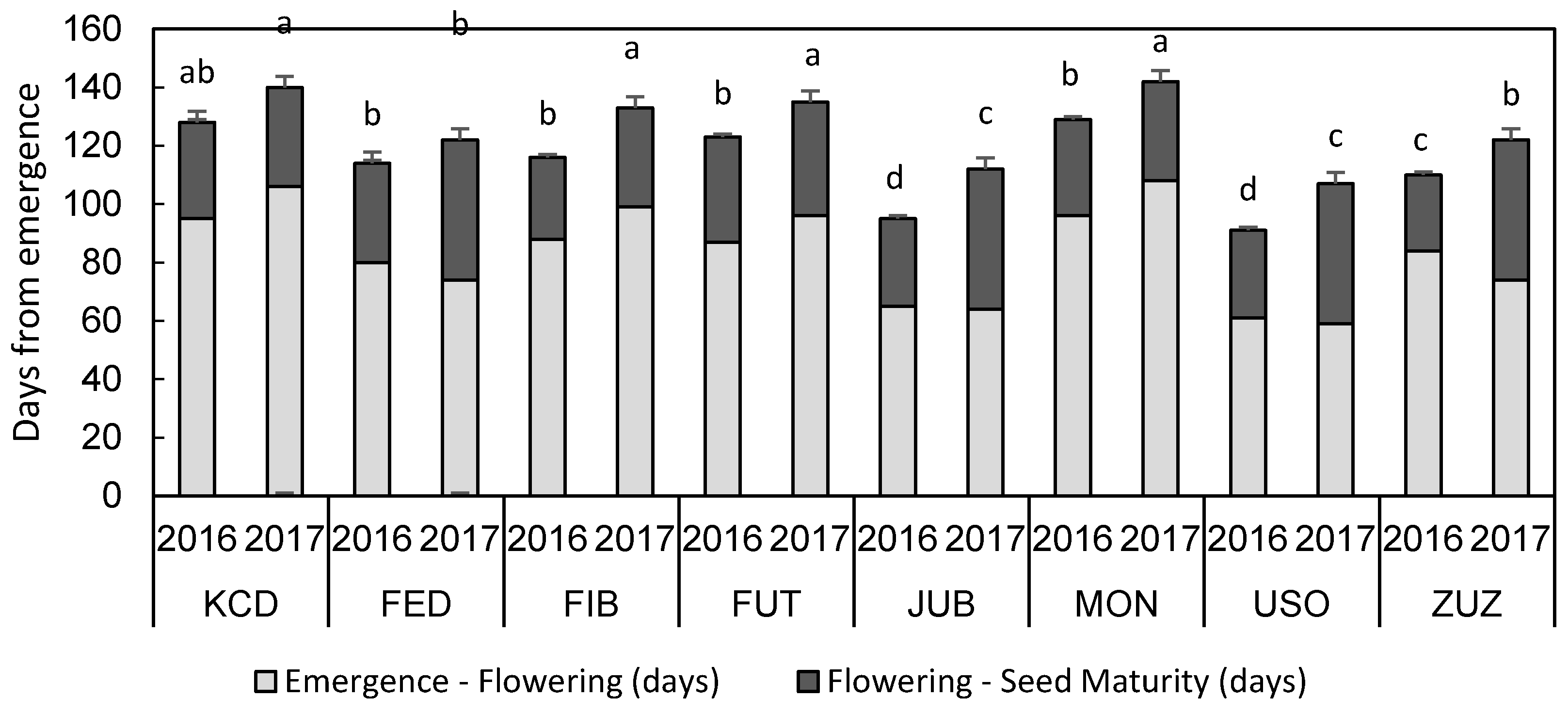
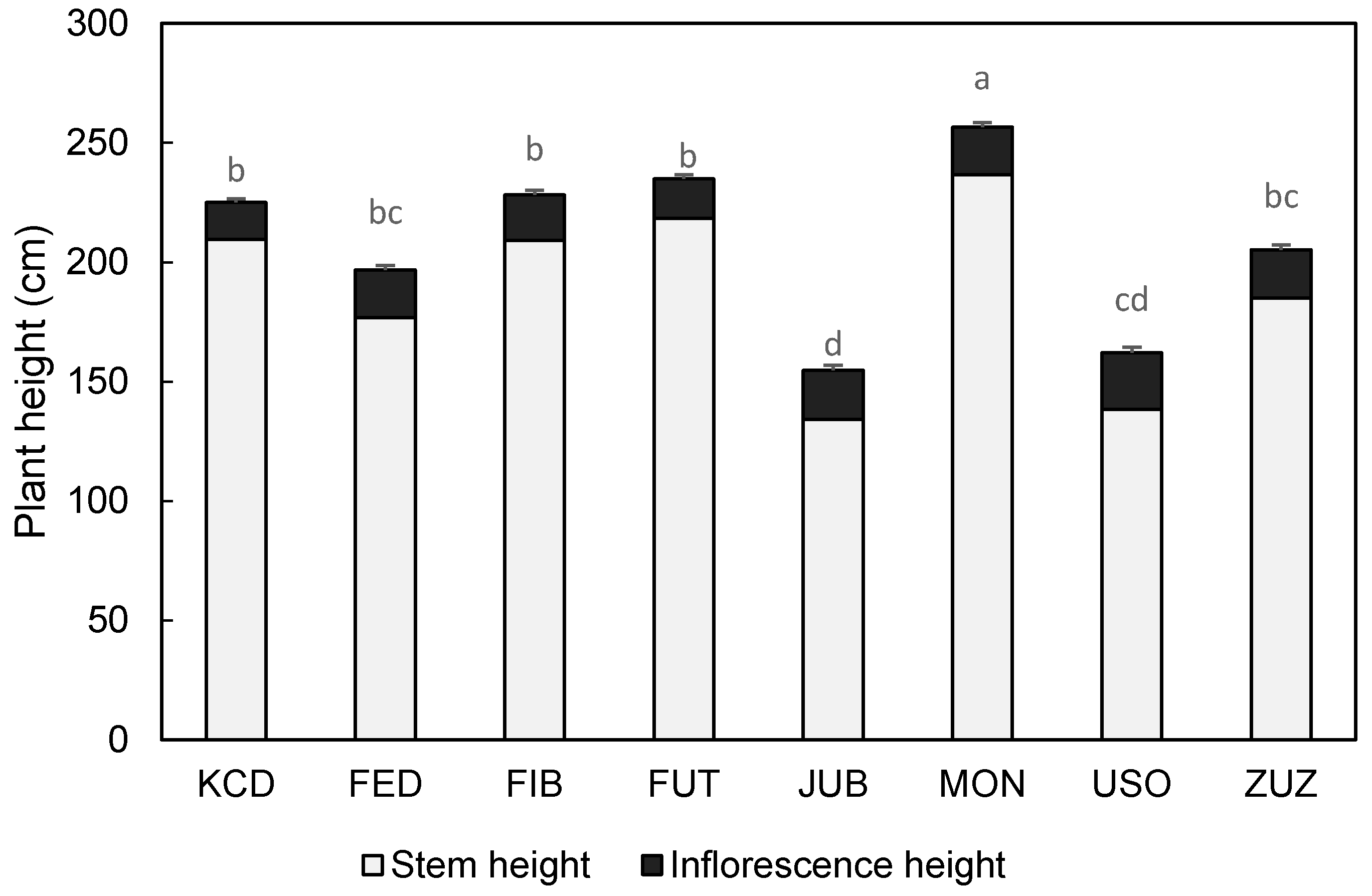
3.2. Phenology and Biometric Characteristics
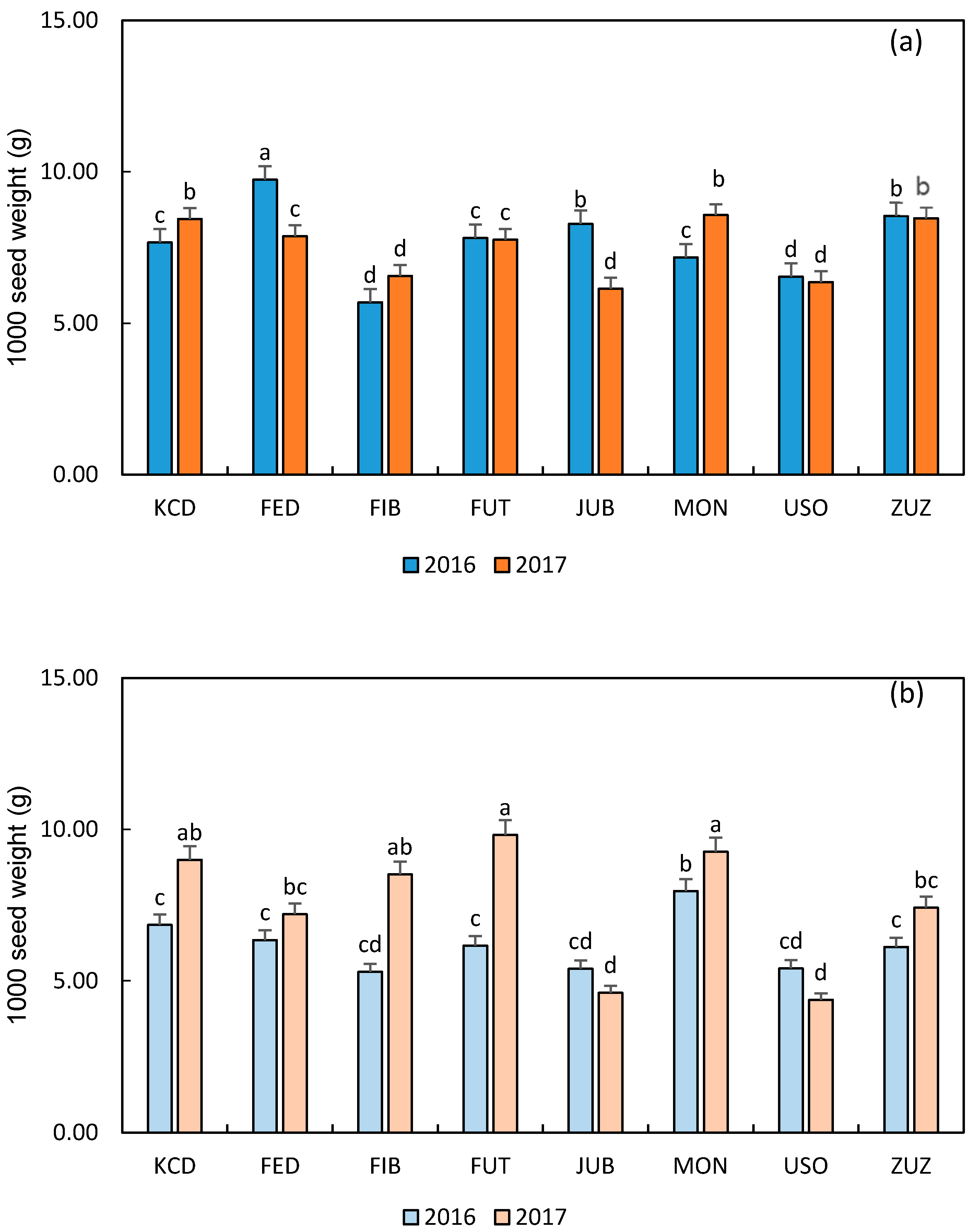
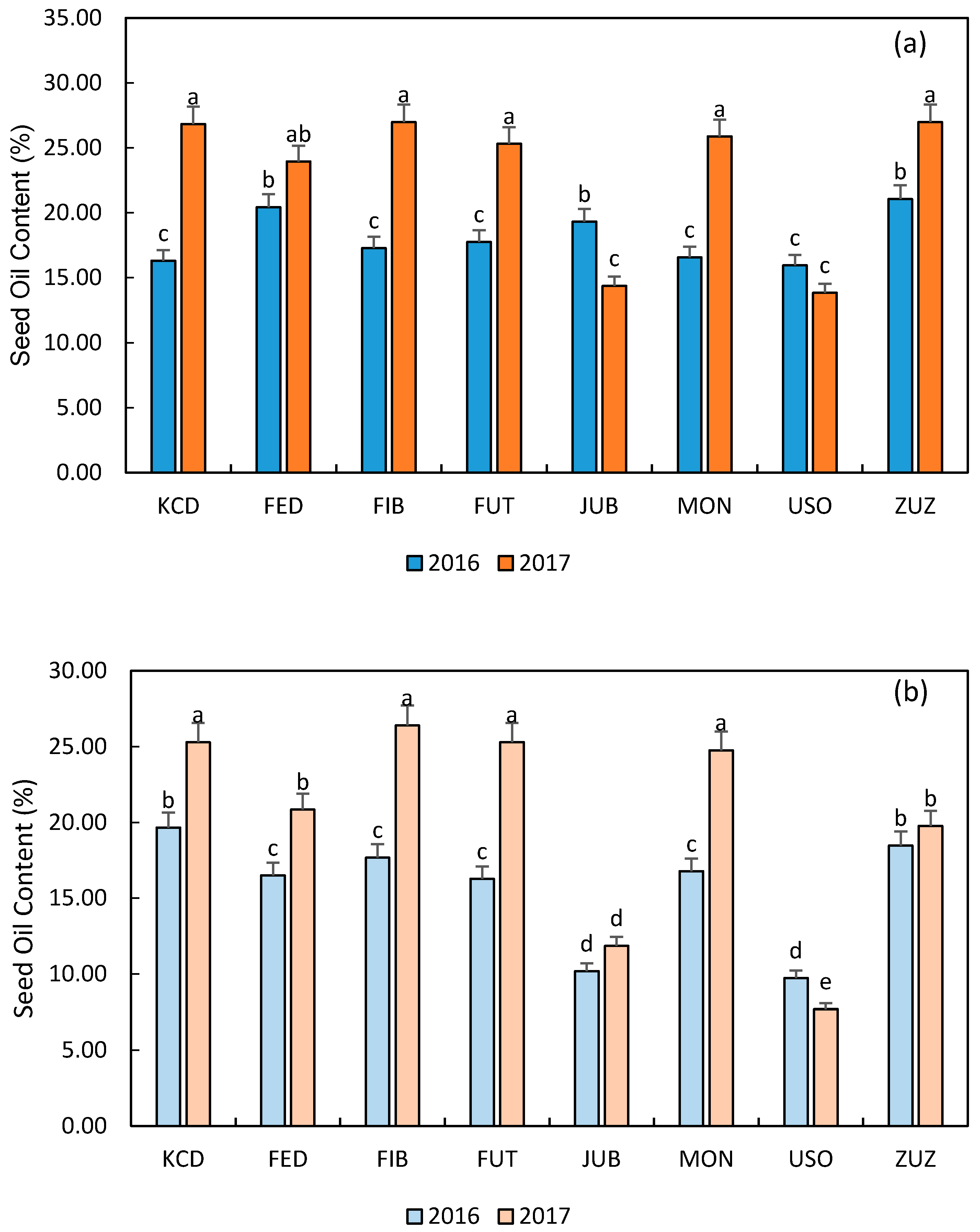
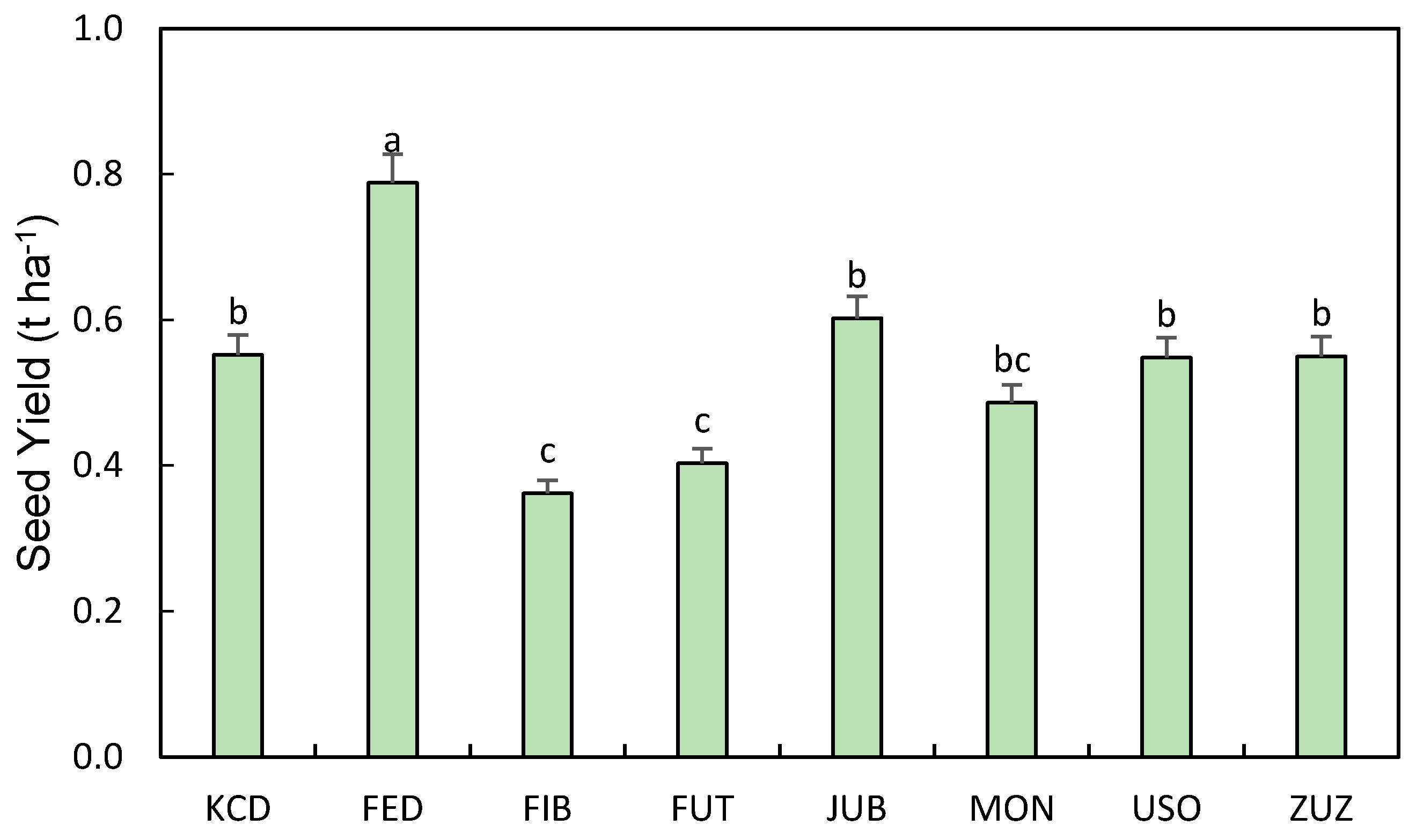
3.3. Main Seed Characteristics
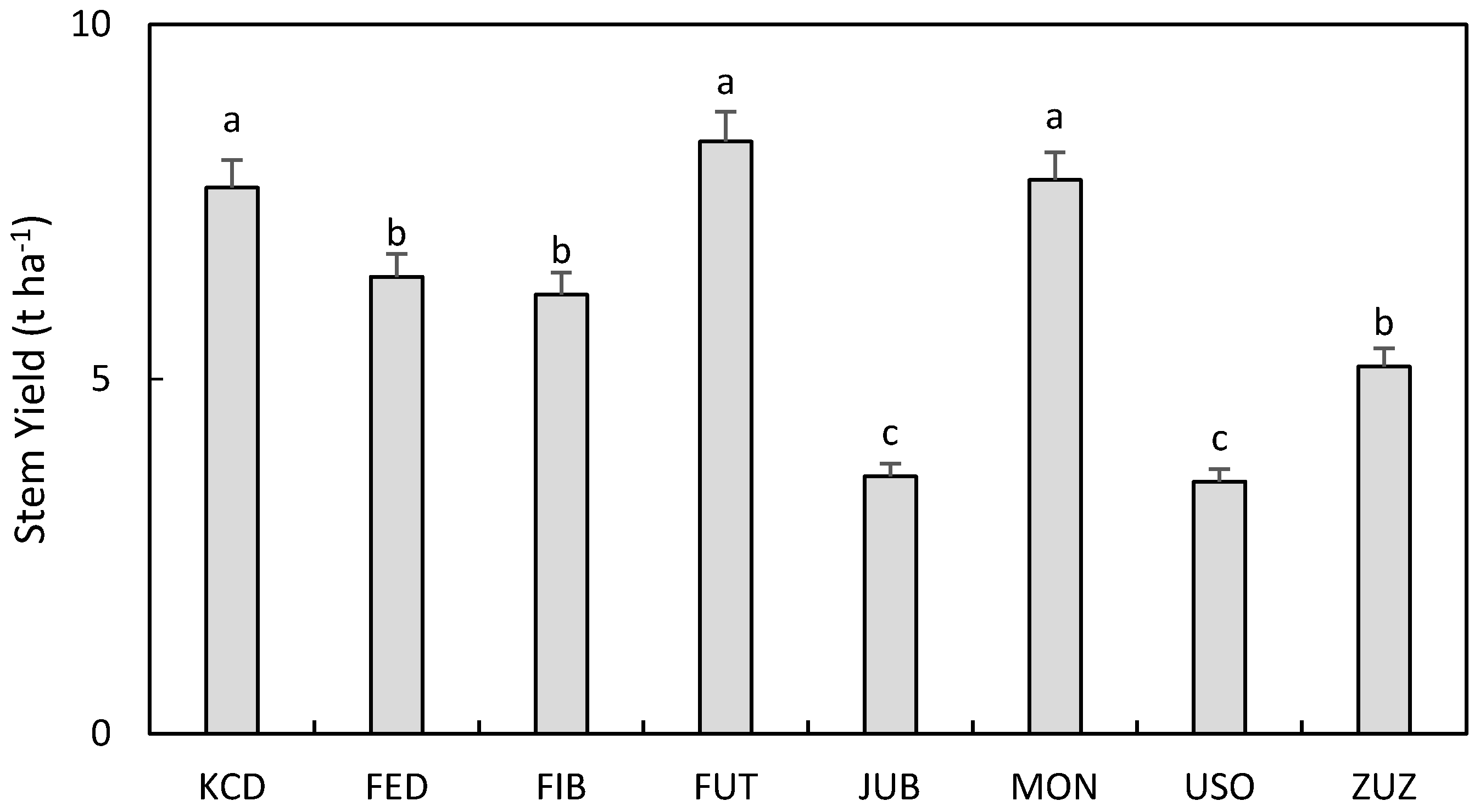
3.4. Biomass, Seed, and Oil Yield
3.5. Oil Fatty Acids Composition
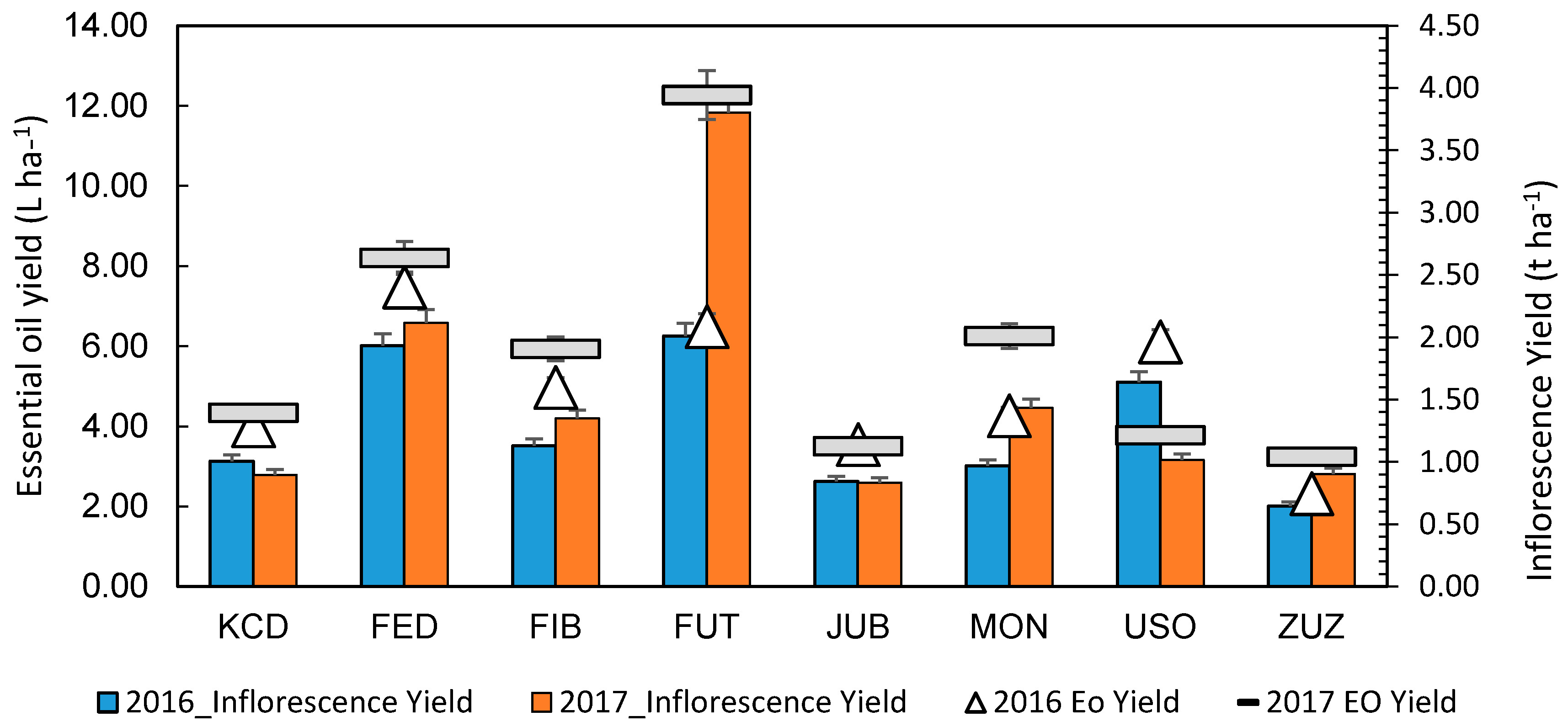
3.6. Essential Oil (EO)
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Allegret, S. The Hystory of Hemp. In Hemp: Industrial Production and Uses Hemp; Bouloc, P., Allegret, S., Arnaud, L., Eds.; CAB international: Bar sur Aube, France, 2013; pp. 4–25. ISBN 9781845937928. [Google Scholar]
- Salentijn, E.M.J.; Zhang, Q.; Amaducci, S.; Yang, M.; Trindade, L.M. New developments in fiber hemp (Cannabis sativa L.) breeding. Ind. Crops Prod. 2015, 68, 32–41. [Google Scholar] [CrossRef]
- Amaducci, S.; Scordia, D.; Liu, F.H.; Zhang, Q.; Guo, H.; Testa, G.; Cosentino, S.L. Key cultivation techniques for hemp in Europe and China. Ind. Crops Prod. 2015, 68, 2–16. [Google Scholar] [CrossRef]
- Tang, K.; Struik, P.; Yin, C.X.; Thouminot, C.; Bjelková, M.; Stramkale, V.; Amaducci, S. Comparing hemp (Cannabis sativa L.) cultivars for dual-purpose production under contrasting environments. Ind. Crops Prod. 2016, 87, 33–44. [Google Scholar] [CrossRef]
- Carus, M.; Karst, S.; Kauffmann, A.; Hobson, J.; Bertucelli, S. The European Hemp Industry: Cultivation, Processing and Applications for Fibres, Shives and Seeds. European Hemp Industry Association, 2013. Available online: http://eiha.org/media/2014/10/13–06-European-Hemp-Industry.pdf (accessed on 5 June 2017).
- European Industrial Hemp Association. Hemp Cultivation Area in Europe. 2016. Available online: http://eiha.org/media/2017/03/17-02-Hemp-Cultivation-Area-Europe.png (accessed on 5 June 2017).
- Mihoc, M.; Pop, G.; Alexa, E.; Radulov, I. Nutritive quality of Romanian hemp varieties (Cannabis sativa L.) with special focus on oil and metal contents of seeds. Chem. Cent. J. 2012, 6, 122. [Google Scholar] [CrossRef] [PubMed]
- Mandolino, G.; Carboni, A.; Bagatta, M.; Moliterni, V.M.C.; Ranalli, P. Occurrence and frequency of putatively Y chromosome linked DNA markers in Cannabis sativa L. Euphytica 2002, 126, 211–218. [Google Scholar] [CrossRef]
- Bertoli, A.; Tozzi, S.; Pistelli, L.; Angelini, L.G. Fibre hemp inflorescences: From crop-residues to essential oil production. Ind. Crops Prod. 2010, 32, 329–337. [Google Scholar] [CrossRef]
- Hazekamp, A.; Fischedick, J.T. Cannabis—From cultivar to chemovar. Drug Test. Anal. 2012, 4, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Appendino, G.; Gibbons, S.; Giana, A.; Pagani, A.; Grassi, G.; Stavri, M.; Smith, E.; Rahman, M.M. Antibacterial cannabinoids from Cannabis sativa: A structure-activity study. J. Nat. Prod. 2008, 71, 1427–1430. [Google Scholar] [CrossRef] [PubMed]
- Nissen, L.; Zatta, A.; Stefanin, I.; Grandi, S.; Sgorbati, B.; Biavati, B.; Monti, A. Characterization and antimicrobial activity of essential oils of industrial hemp varieties (Cannabis sativa L.). Fitoterapia 2010, 81, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Mukhtar, T.; Kayani, M.Z.; Hussain, M.A. Nematicidal activities of Cannabis sativa L. and Zanthoxylum alatum Roxb. against Meloidogyne incognita. Ind. Crops Prod. 2013, 42, 447–453. [Google Scholar] [CrossRef]
- Grewal, P.S. Effects of leaf-matter incorporation on Aphelenchoides composticola (Nematoda), mycofloral composition, mushroom compost quality and yield of Agaricus bisporus. Ann. Appl. Biol. 1989, 115, 299–312. [Google Scholar] [CrossRef]
- Zia, A.; Aslam, M.; Naz, F.; Illyas, M. Bio-efficacy of some plant extracts against chickpea beetle, Callosobruchus chinensis Linnaeus (Coleoptera: Bruchidae) attacking chickpea. Pak. J. Zool. 2011, 43, 733–737. [Google Scholar]
- Górski, R.; Szklarz, M.; Kaniewski, R. Efficacy of hemp essential oil in the control of rosy apple aphid (Dysaphis plantaginea Pass.) occurring on apple tree. Prog. Plant Prot. 2009, 49, 2013–2016. [Google Scholar]
- Bedini, S.; Flamini, G.; Cosci, F.; Ascrizzi, R.; Benelli, G.; Conti, B. Cannabis sativa and Humulus lupulus essential oils as novel control tools against the invasive mosquito Aedes albopictus and fresh water snail Physella acuta. Ind. Crops Prod. 2016, 85, 318–323. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R.; Lupidi, G.; Nabissi, M.; Petrelli, R.; Ngahang Kamte, S.L.; Cappellacci, L.; Fiorini, D.; Sut, S.; Dall’Acqua, S.; et al. The crop-residue of fiber hemp cv. Futura 75: From a waste product to a source of botanical insecticides. Environ. Sci. Pollut. Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Amaducci, S.; Zatta, A.; Pelatti, F.; Venturi, G. Influence of agronomic factors on yield and quality of hemp (Cannabis sativa L.) fibre and implication for an innovative production system. Field Crops Res. 2008, 107, 161–169. [Google Scholar] [CrossRef]
- Mediavilla, V.; Jonquera, M.; Schmid-Slembrouck, I.; Soldati, A. Decimal code for growth stages of hemp (Cannabis sativa L.). J. Int. Hemp Assoc. 1998, 5, 68–74. [Google Scholar]
- Amaducci, S.; Colauzzi, M.; Bellocchi, G.; Venturi, G. Modelling post-emergent hemp phenology (Cannabis sativa L.): Theory and evaluation. Eur. J. Agron. 2008, 28, 90–102. [Google Scholar] [CrossRef]
- Bócsa, I.; Karus, M. The Cultivation of Hemp: Botany, Varieties, Cultivation and Harvesting; Hemptech: Sebastopol, CA, USA, 1998; p. 184. ISBN 978-1886874039. [Google Scholar]
- Heslop-Harrison, J.; Heslop-Harrison, Y. Cannabis sativa L. In The Induction of Flowering; Evans, L.T., Ed.; Macmillan: Canberra, Australia, 1969; pp. 205–226. [Google Scholar]
- De Meijer, E.P.M.; Keizer, L.C.P. Patterns of diversity in Cannabis. Genet. Resour. Crop Evol. 1996, 43, 41–52. [Google Scholar] [CrossRef]
- Cosentino, S.L.; Testa, G.; Scordia, D.; Copani, V. Sowing time and prediction of flowering of different hemp (Cannabis sativa L.) genotypes in southern Europe. Ind. Crops Prod. 2012, 37, 20–33. [Google Scholar] [CrossRef]
- Faux, A.M.; Draye, X.; Lambert, R.; d’Andrimont, R.; Raulier, P.; Bertin, P. The relationship of stem and seed yields to flowering phenology and sex expression in monoecious hemp (Cannabis sativa L.). Eur. J. Agron. 2013, 47, 11–22. [Google Scholar] [CrossRef]
- Cromack, H.T.H. The effect of cultivar and seed density on the production and fibre content of Cannabis sativa L. in southern England. Ind. Crops Prod. 1998, 7, 205–210. [Google Scholar] [CrossRef]
- House, J.D.; Neufeld, J.; Leson, G. Evaluating the quality of protein from hemp seed (Cannabis sativa L.) products through the use of the protein digestibility-corrected amino acid score method. J. Agric. Food Chem. 2010, 58, 11801–11807. [Google Scholar] [CrossRef] [PubMed]
- Vonapartis, E.; Aubin, M.P.; Seguin, P.; Mustafa, A.F.; Charron, J.B. Seed composition of ten industrial hemp cultivars approved for production in Canada. J. Food Compos. Anal. 2015, 39, 8–12. [Google Scholar] [CrossRef]
- Deferne, J.L.; Pate, D.W. Hemp seed oil: A source of valuable essential fatty acids. J. Int. Hemp Assoc. 1996, 3, 4–7. [Google Scholar]
- Pate, D.W. Hemp seed: A valuable food source. In Advances in Hemp Research, Ranalli, P., Ed.; The Haworth Press: NY, USA, 1999; pp. 243–255. [Google Scholar]
- Johnson, P. Industrial hemp: A critical review of claimed potentials for Cannabis sativa. Tappi J. 1999, 82, 113–123. [Google Scholar]
- Rondanini, D.; Mantese, A.; Savin, R.; Hall, A.J. Responses of sunflower yield and grain quality to alternating day/night high temperature regimes during grain filling: Effects of timing, duration and intensity of exposure to stress. Field Crops Res. 2006, 96, 48–62. [Google Scholar] [CrossRef]
- Rondanini, D.; Savin, R.; Hall, A.J. Dynamics of fruit growth and oil quality of sunflower (Helianthus annuus L.) exposed to brief intervals of high temperature during grain filling. Field Crops Res. 2003, 83, 79–90. [Google Scholar] [CrossRef]
- Connor, D.J.; Hall, A.J. Sunflower physiology. In Sunflower Technology and Production; Schneiter, A.A., Ed.; ASA, SCSA and SSSA: Madison, WI, USA, 1997; pp. 113–182. [Google Scholar]
- Harris, H.C.; McWilliam, J.R.; Mason, W.K. Influence of temperature on oil content and composition of sunflower seed. Aust. J. Agric. Res. 1978, 29, 1203–1212. [Google Scholar] [CrossRef]
- Chimenti, C.A.; Hall, A.J. Grain number responses to temperature during floret differentiation in sunflower. Field Crops Res. 2001, 72, 177–184. [Google Scholar] [CrossRef]
- Desanlis, F.; Cerruti, N.; Warner, P. Hemp agronomics and cultivation. In Hemp: Industrial Production and Uses Hemp; Bouloc, P., Allegret, S., Arnaud, L., Eds.; CAB international: Bar sur Aube, France, 2013; pp. 98–124. ISBN 9781845937928. [Google Scholar]
- Galasso, I.; Russo, R.; Mapelli, S.; Ponzoni, E.; Brambilla, I.M.; Battelli, G.; Reggiani, R. Variability in seed traits in a collection of Cannabis sativa L. genotypes. Front. Plant Sci. 2016, 7, 688. [Google Scholar] [CrossRef] [PubMed]
- Calloway, J.C.; Laakkonen, T.T. Cultivation of Cannabis oil seed varieties in Finland. J. Int. Hemp Assoc. 1996, 3, 32–34. [Google Scholar]
- Da Porto, C.; Decorti, D.; Tubaro, F. Fatty acid composition and oxidation stability of hemp (Cannabis sativa L.) seed oil extracted by supercritical carbon dioxide. Ind. Crops Prod. 2012, 36, 401–404. [Google Scholar] [CrossRef]
- Leizer, C.; Ribnicky, D.; Poulev, A.; Dushenkov, S.; Raskin, I. The composition of hemp seed oil and its potential as an important source of nutrition. J. Nutraceuticals Funct. Med. Foods 2000, 2, 35–53. [Google Scholar] [CrossRef]
- Garces, R.; Mancha, M. In vitro oleate desaturate in developing sunflower seed. Phytochemistry 1991, 30, 2127–2130. [Google Scholar] [CrossRef]
- Verma, R.S.; Padalia, R.C.; Verma, S.K.; Chauhan, A.; Darokar, M.P. The essential oil of ‘bhang’ (Cannabis sativa L.) for non-narcotic applications. Curr. Sci. 2014, 107, 645–650. [Google Scholar]
- Meier, C.; Mediavilla, V. Factors influencing the yield and the quality of hemp (Cannabis sativa L.) essential oil. J. Int. Hemp Assoc. 1998, 5, 16–20. [Google Scholar]









{kind=link}
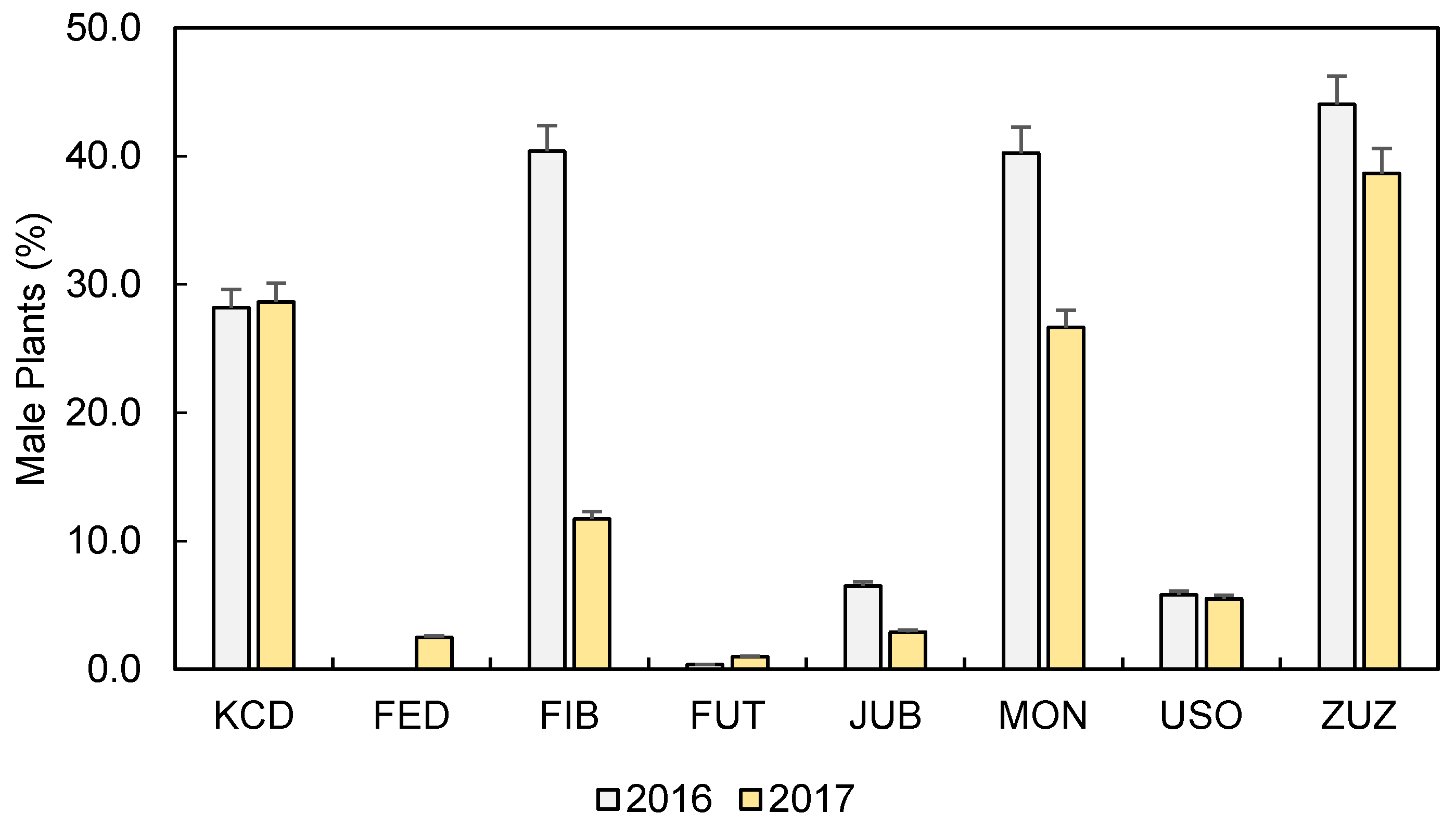{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Origin | Maturity Group |
|---|---|---|
| Jubileu | Romania | Very Early |
| Uso-31 | Ukraine | Early |
| KC Zuzana | Hungary | Early |
| Fedora 17 | France | Medium-Early |
| Fibrol | Hungary | Medium |
| Monoica | Hungary | Medium |
| KC Dora | Hungary | Late |
| Futura 75 | France | Late |
| Month | 2016 | 2017 | 1992–2016 Average | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Tmin (°C) | Tmax (°C) | Rainfall (mm) | Tmin (°C) | Tmax (°C) | Rainfall (mm) | Tmin (°C) | Tmax (°C) | Rainfall (mm) | |
| April | 8.5 | 19.2 | 66.3 | 7.4 | 19.7 | 147.5 | 7.4 | 18.3 | 121.6 |
| May | 11.1 | 22.3 | 238.3 | 11.8 | 24.2 | 80.6 | 12.2 | 23.4 | 122.6 |
| June | 15.9 | 26.5 | 122.4 | 16.2 | 29.3 | 163.6 | 15.5 | 26.9 | 124.9 |
| July | 17.7 | 30.6 | 92.3 | 17.4 | 30.3 | 61.8 | 17.0 | 29.4 | 117.7 |
| August | 15.0 | 29.1 | 68.7 | 17.4 | 31.3 | 47.0 | 16.7 | 29.4 | 131.6 |
| September | 14.4 | 27.4 | 113.9 | 11.8 | 21.6 | 343.9 | 12.6 | 24.5 | 162.4 |
| October | 8.2 | 18.4 | 123.8 | 7.2 | 19.8 | 38.2 | 9.0 | 19.0 | 167.8 |
| Mean | 13.0 | 24.8 | 12.8 | 25.2 | 12.9 | 24.4 | |||
| Total | 825.7 | 882.76 | 948.6 | ||||||
| Source | Plant Height | Male Plants Percentage | Inflorescence Height | Stem Yield | Oil Yield | Seed | ||
|---|---|---|---|---|---|---|---|---|
| Yield | SW a | Oil Content | ||||||
| Year (Y) | *** | ** | ns | ** | * | *** | ns | *** |
| Cultivar (C) | *** | *** | ns | *** | *** | *** | *** | *** |
| C × Y | ns | * | ns | ns | ns | ns | * | *** |
| Cultivar | Seed Yield (t ha−1) | Oil Yield (t ha−1) | ||
|---|---|---|---|---|
| Control (Regular Plants) | With Inflorescence Harvested | Control (Regular Plants) | With Inflorescence Harvested | |
| Kc Dora | 0.55 b | 0.37 b | 0.12 b | 0.08 ab |
| Fedora 17 | 0.79 a | 0.51 a | 0.17 a | 0.09 a |
| Fibrol | 0.36 e | 0.23 e | 0.07 cd | 0.05 c |
| Futura 75 | 0.40 d | 0.30 c | 0.09 c | 0.06 c |
| Jubileu | 0.60 b | 0.38 b | 0.10 c | 0.04 cd |
| Monoica | 0.49 c | 0.26 d | 0.09 c | 0.05 c |
| Uso-31 | 0.55 b | 0.37 b | 0.08 c | 0.03 d |
| Kc Zuzana | 0.55 b | 0.46 a | 0.13 b | 0.08 a |
| Average | 0.55 | 0.36 | 0.11 | 0.06 |
| Source | Seed Yield | Oil Content | 1000 Seed Weight | Oil Yield |
|---|---|---|---|---|
| Year (Y) | *** | *** | *** | ** |
| Cultivar (C) | *** | *** | *** | ** |
| C × Y | ns | ns | ** | ns |
| Cultivar | Fatty Acid (%) | ω3/ω6 Ratio | ||||||
|---|---|---|---|---|---|---|---|---|
| Palmitic | Stearic | Oleic | Linoleic | α-Linolenic | γ-Linolenic | Others | ||
| Fedora 17 | 7.8 ± 0.35 | 2.9 ± 0.18 | 14.1 ± 0.67 | 56.3 ± 0.39 | 15.7 ± 0.11 | 2.2 ± 0.02 | 0.7 ± 0.40 | 0.32 |
| Fibrol | 8.1 ± 0.13 | 2.8 ± 0.01 | 16.7 ± 0.27 | 55.1 ± 0.14 | 15.5 ± 0.25 | 1.2 ± 0.22 | 0.5 ± 0.01 | 0.30 |
| Futura 75 | 7.5 ± 0.31 | 2.9 ± 0.10 | 15.0 ± 0.19 | 56.2 ± 0.20 | 16.0 ± 0.69 | 1.4 ± 0.01 | 0.9 ± 0.46 | 0.31 |
| Jubileu | 7.2 ± 0.54 | 3.0 ± 0.08 | 14.4 ± 0.45 | 55.6 ± 0.40 | 16.6 ± 0.11 | 2.4 ± 0.10 | 0.8 ± 0.08 | 0.34 |
| KC Dora | 7.6 ± 0.49 | 3.1 ± 0.07 | 13.7 ± 0.11 | 56.0 ± 0.90 | 17.7 ± 0.24 | 1.2 ± 0.05 | 0.7 ± 0.16 | 0.34 |
| Monoica | 7.3 ± 0.67 | 2.5 ± 0.08 | 16.2 ± 0.61 | 56.7 ± 0.62 | 16.1 ± 0.63 | 0.9 ± 0.34 | 0.4 ± 0.06 | 0.30 |
| Uso-31 | 6.9 ± 0.17 | 2.8 ± 0.09 | 14.9 ± 1.02 | 56.1 ± 0.40 | 16.2 ± 0.58 | 2.3 ± 0.11 | 0.8 ± 0.04 | 0.33 |
| KC Zuzana | 7.0 ± 0.00 | 2.8 ± 0.00 | 15.1 ± 0.00 | 55.9 ± 0.00 | 16.8 ± 0.00 | 1.6 ± 0.00 | 0.9 ± 0.00 | 0.33 |
| Average | 7.4 ± 0.30 | 2.9 ± 0.12 | 15.0 ± 0.41 | 56.0 ± 0.39 | 16.3 ± 0.29 | 1.7 ± 0.14 | 0.7 ± 0.14 | 0.32 |
| Source | Inflorescence Yield | EO Concentration | EO Yield |
|---|---|---|---|
| Year (Y) | * | ns | * |
| Cultivar (C) | *** | ** | *** |
| C × Y | * | ns | ** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baldini, M.; Ferfuia, C.; Piani, B.; Sepulcri, A.; Dorigo, G.; Zuliani, F.; Danuso, F.; Cattivello, C. The Performance and Potentiality of Monoecious Hemp (Cannabis sativa L.) Cultivars as a Multipurpose Crop. Agronomy 2018, 8, 162. https://doi.org/10.3390/agronomy8090162
Baldini M, Ferfuia C, Piani B, Sepulcri A, Dorigo G, Zuliani F, Danuso F, Cattivello C. The Performance and Potentiality of Monoecious Hemp (Cannabis sativa L.) Cultivars as a Multipurpose Crop. Agronomy. 2018; 8(9):162. https://doi.org/10.3390/agronomy8090162
Chicago/Turabian StyleBaldini, Mario, Claudio Ferfuia, Barbara Piani, Angela Sepulcri, Gaia Dorigo, Fabio Zuliani, Francesco Danuso, and Costantino Cattivello. 2018. "The Performance and Potentiality of Monoecious Hemp (Cannabis sativa L.) Cultivars as a Multipurpose Crop" Agronomy 8, no. 9: 162. https://doi.org/10.3390/agronomy8090162
APA StyleBaldini, M., Ferfuia, C., Piani, B., Sepulcri, A., Dorigo, G., Zuliani, F., Danuso, F., & Cattivello, C. (2018). The Performance and Potentiality of Monoecious Hemp (Cannabis sativa L.) Cultivars as a Multipurpose Crop. Agronomy, 8(9), 162. https://doi.org/10.3390/agronomy8090162