1. Introduction
Groundnut or peanut (
Arachis hypogaea L.) is an important oilseed crop in the world. It is a key crop for small farmers, especially in Africa and Asia, chiefly cultivated for its oil and food value. It contains about 45–56% of high quality edible oil, 25–30% protein, 20% carbohydrate, and is a rich source of dietary fiber, minerals, and vitamins E and B [
1,
2]. The haulm is a nutritious animal feed. In addition, groundnut helps to maintain soil fertility by fixing atmospheric nitrogen to the soil.
In 2016, the global groundnut production was about 44 million (M) tons from an area covering about 27.7 M hectares. The continent of Asia ranked first with 60.1% of global production, followed by Africa with about 29% of global production. The largest groundnut producing countries were China and India with 16.7 M and 6.9 M tons, respectively. Nigeria ranked first and third in Africa and the world, respectively, with about 3 M tons in 2016. In Burkina Faso, groundnut production was estimated at about 335,715 tons in 2016, obtained from an area of about 420,000 hectares, and with an average yield of about 799.3 kg/ha [
3]. In general, groundnut productivity in Africa had the lowest average yield (902.6 kg/ha) in 2016 compared to America (3381.4 kg/ha), Asia (2186.8 kg/ha), Oceania (1947.3 kg/ha), Europe (3102.1 kg/ha), and the global average yield (1590.1 kg/ha) [
3]. Several abiotic, biotic, and socio-economic constraints are responsible for the low yield in Africa. Among biotic factors, foliar fungal diseases are the major production constraints of groundnut [
1,
2]. Of these, early leaf spot (ELS), caused by
Cercospora arachidicola, is a major and widely distributed disease. This disease, combined with rust or late leaf spot, can cause from 50% to 70% yield losses [
4].
Control methods have been developed to minimize yield losses due to ELS, which include host plant resistance, and cultural and chemical control methods [
5]. Fungicides, such as Maneb and Mancozeb, which belong to the ethylene bisdithiocarbamate group of chemicals, are effective in controlling ELS. However, chemical controls may not be economical for smallholder farmers in developing countries due to a number of reasons. These include low basic yields; difficulties in obtaining fungicides and application machinery, and their high costs; the problem of access to sources of clean water and of transporting it in sufficient quantities for high- or medium-volume spraying; lack of expertise, and lack of advice on the use of spray machinery and on its maintenance; and low or fluctuating prices for groundnut, discouraging farmers from risk-taking investment in the crop [
6]. Besides, chemicals are potentially harmful to the environment and its inhabitants. Therefore, host plant resistance is considered the cheapest and most effective control method. Accordingly, the development of high yielding cultivars with resistance to ELS is an important breeding priority to increase groundnut productivity and reduce the impacts of the disease. A nine-point disease scale is used for measuring reactions to ELS [
6]. Although complete resistance to ELS has not been found in the cultivated groundnut, several promising lines were identified as resistance sources [
7,
8,
9]. The search for new sources of ELS resistance both from cultivated and wild relatives is a continuous process to developing ELS-resistant and high yielding varieties that are adaptable to target environments and preferred by farmers, markets, and consumers.
Understanding the inheritance of ELS is important to breeding for resistant groundnut cultivars. Previous studies revealed that resistance to ELS is recessive and independently inherited, and both quantitative and qualitative inheritance have been reported [
10]. Genetic analysis using diallel mating design is useful to identify genotypes with combining ability in a desirable direction, and to identify parents with additive and non-additive effects for specific traits that may be used in breeding [
11,
12,
13]. The knowledge on combining the ability and type of gene action responsible for the regulation of expression of ELS disease helps in planning appropriate breeding strategies. Therefore, this study was set up to generate data on ELS inheritance for groundnut breeding programs in West and Central Africa using groundnut genotypes from the region. The specific objectives were: (1) elucidating the mode of inheritance of genes governing the expression of ELS disease in populations under this study, (2) identifying good general combiners to be used as donor parents for ELS disease resistance, and (3) evaluating the breeding potential of the populations to identify ELS-resistant varieties.
4. Discussion
The six parental lines and their 30 F2 progenies showed highly significant differences for the ELS severity score and AUDPC. These differences were due to the considerable genetic diversity among the materials tested. High variability among groundnut genotypes in terms of leaf spot disease resistance, including ELS, were also previously reported [
1,
25,
26,
27].
The Griffing analysis of variance indicated a highly significant GCA for ELS severity and AUDPC, indicating the role of additive gene action in the control of the two traits. On the other hand, the SCA was significant for the ELS severity score only. The results suggest that both additive and non-additive gene effects are involved in the expression of ELS disease resistance in groundnut. Similar findings of additive and non-additive gene actions for yield-related traits and diseases, including ELS resistance, were reported earlier [
1,
26,
27,
28,
29]. In addition, the reciprocal effect was highly significant for the two traits, indicating the contribution of cytoplasm and nuclear genes effects to ELS resistance. The estimates of the genetic components indicated that variance due to GCA was higher in magnitude compared to variance due to SCA for the two traits. Their ratio was greater than unity, indicating the profound influence of additive gene action in the control of ELS disease in groundnut. This is a desirable phenomenon necessary for making progress in breeding for ELS resistance. These findings are in accordance with previous results [
1,
26,
27,
30].
The general combining ability indicates the average performance of a line in cross combination [
22]. In general, desirable parents are those with significant GCA effects in the right direction for the trait of interest [
31]. In this study, negative GCA effects were desirable, since they indicate a contribution to resistance, while positive values indicate a contribution to susceptibility. The NAMA parents, followed by PC79-79 and B188 recorded negative GCA, suggesting that the use of these parents in breeding for resistance to ELS resistance would be expected to produce progenies with increased resistance. In addition, these three parents showed low mean disease scores. Some crosses involving these resistant parents exhibited negative SCA effects. For the SCA effects, 10 of 30 possible combinations exhibited negative SCA effects for ELS resistance components, indicating that the resistance of these progenies was higher than would be expected from the average of their parents. For the ELS severity score, five cross combinations, including NAMA × B188, PC79-79 × TS32-1, PC79-79 × CN94C, QH243C × TS32-1, and QH243C × CN94C, showed negative SCA effects. For AUDPC, negative SCA effects were observed for nine combinations (NAMA × PC79-79, NAMA × B188, NAMA × CN94C, B188 × PC79-79, B188 × QH243C, PC79-79 × TS32-1, QH243C × TS32-1, QH243C × CN94C, and TS32-1 × CN94C). Four cross combinations (NAMA × B188, PC79-79 × TS32-1, PC79-79 × CN94C, and QH243C × TS32-1) showed negative SCA effects for both ELS severity score and AUDPC. These crosses could be considered the best cross combinations for ELS resistance.
Hayman’s analysis revealed that both the additive “a” and dominance “b” genetic components were significant, indicating their importance in the regulation of ELS resistance in groundnut. The results were similar to Griffing’s analysis. Additionally, the mean dominance effect “b1”, additional dominance effect due to the parents “b2”, and residual dominance effect “b3” were significant for both the ELS severity score and AUDPC. The significance of “b1” indicated that the dominance was unidirectional, while the significance of “b2” indicated an asymmetry of gene distribution. Thus, susceptible parents (QH243C, TS32-1, and CN94C) harbored considerably higher numbers of dominant genes than the resistant parents NAMA, PC79-79, and B188. The presence of specific dominance in some crosses (NAMA × B188, PC79-79 × TS32-1, QH243C × TS32-1, PC79-79 × CN94C, and QH243C × CN94C) was confirmed by the significance of “b3” for the ELS severity score. Similar results were earlier reported for rust [
25] and for leaf spot diseases of groundnut, including ELS [
15,
26].
The results also revealed a predominance of additive variance (D) compared to the components associated with dominance variance (H
1 and H
2) for the ELS severity score and AUDPC. These indicate that resistance to ELS is predominantly regulated by additive gene action. The findings are in harmony with the results of Griffing’s analysis. Considering the dominance components, the magnitude of H
1 was greater than H
2, indicating the non equality of dominant and recessive alleles in all loci controlling ELS resistance. For both the ELS severity score and AUDPC, positive F item values were observed, which measured the covariance of the additive and dominance effects, indicating the predominance of dominant alleles in the parents. The high F value also indicated that positive and negative alleles were not equal in proportion. The proportion of positive alleles was greater than that of the negative alleles in the parents at any locus. The ratio of (H
1/D)
1/2, which measures the average degree of dominance, and the ratio of the total number of dominant to recessive genes in all parents (kd/(kd + kr) were less than unity, suggesting partial dominance and confirming the presence of recessive alleles for ELS resistance in all parents. Heritability estimates showed high (>60%) broad sense heritability and narrow heritability for the two related traits studied, indicating that ELS resistance is highly heritable. Similar results were reported for ELS and late leaf spot [
32,
33,
34].
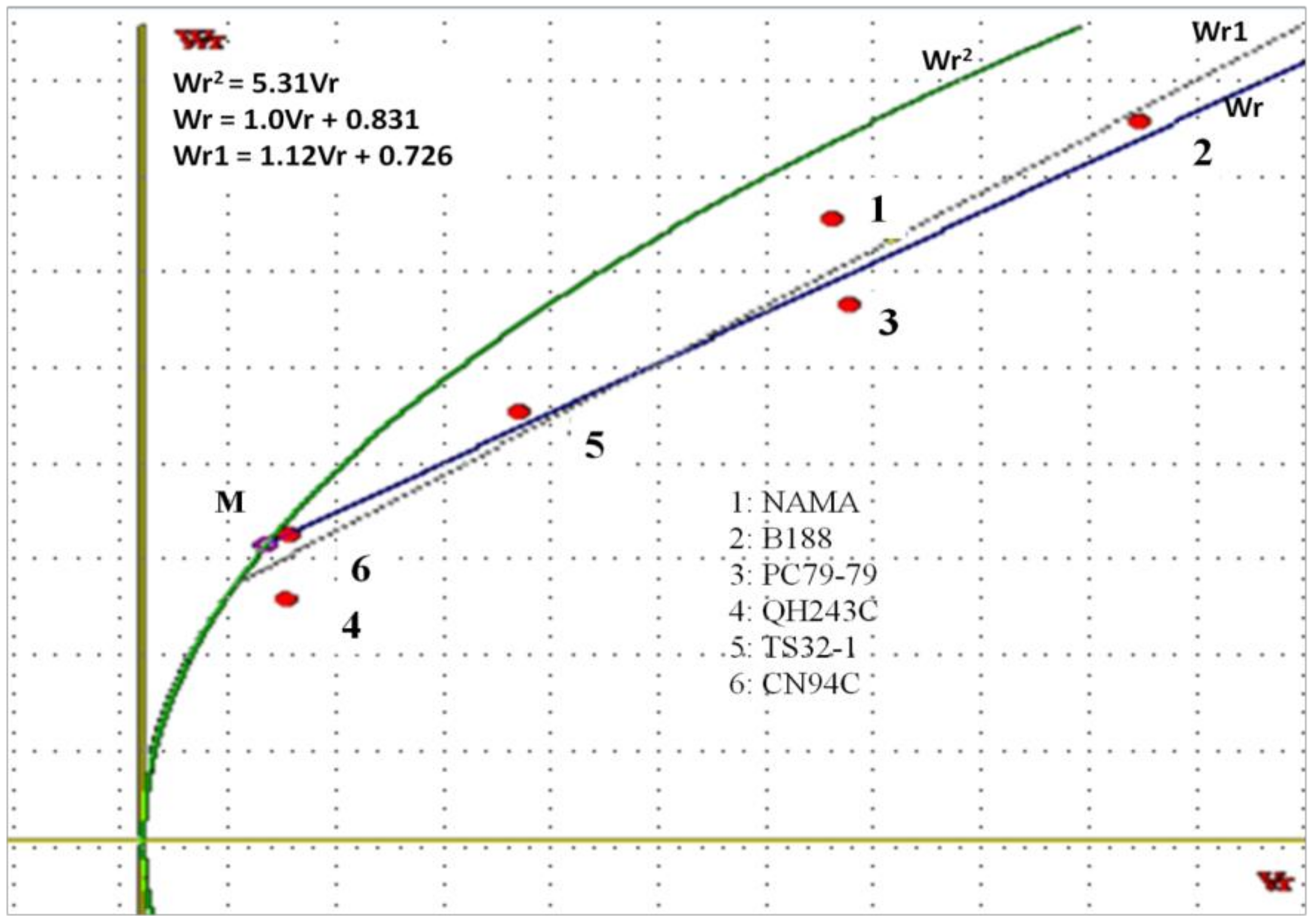
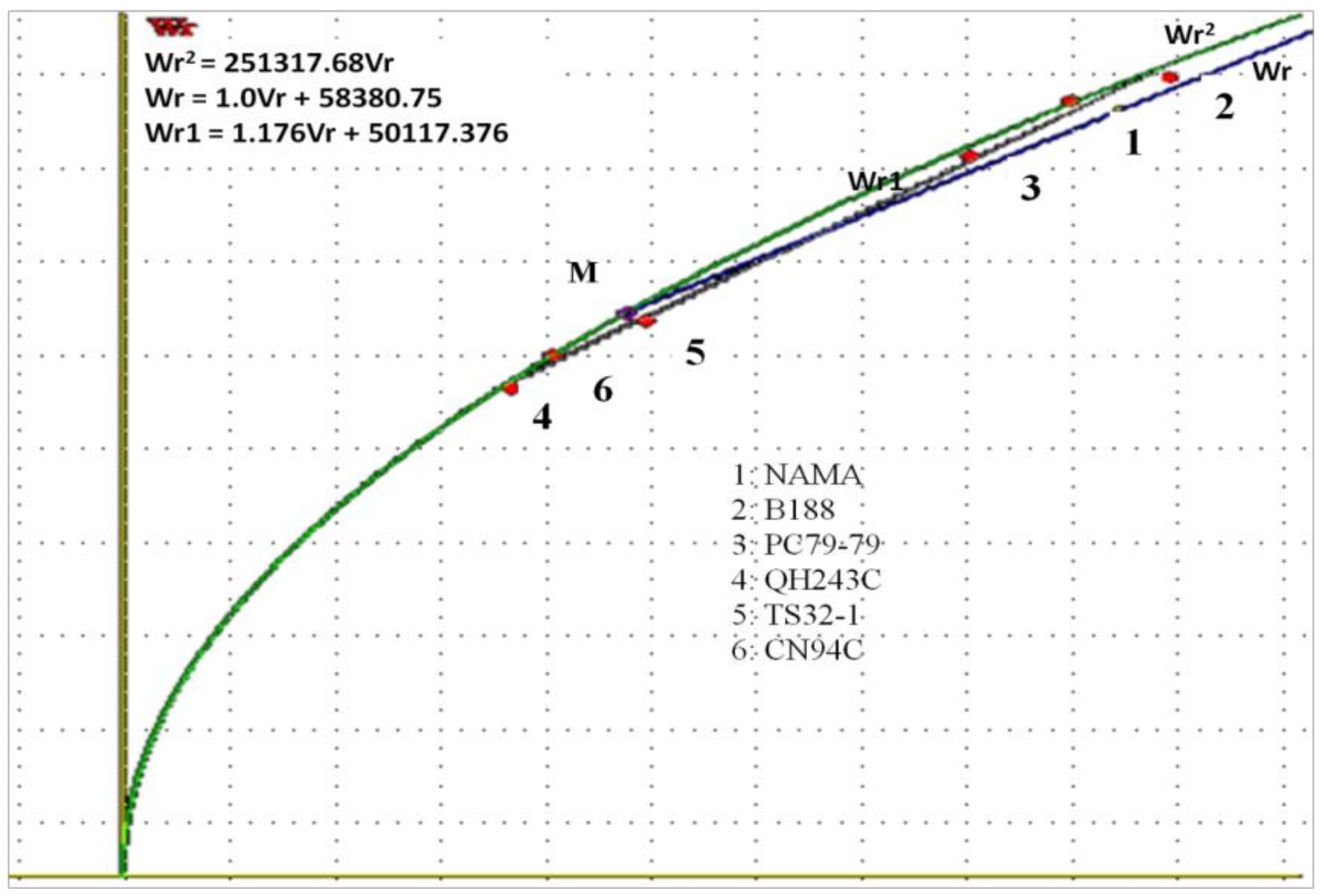
The Vr–Wr graph showed a coefficient of regression (1.01) not significantly different from unity for the two traits, suggesting the absence of epistasis, and rather additive and dominance gene action in ELS expression. An average partial dominance might be considered, since the regression line crossed the Wr axis above the origin. These results are in accordance with those reported for yield and yield-related traits [
1]. Considering the distribution of array points along with the regression line, the parents QH243C, TS32-1, and CN94C were clustered closer to the origin of the regression line, suggesting that they contain larger proportions of dominant alleles. However, the resistant parents NAMA, PC79-79, and B188 were farthest from the origin and closer to the middle of the regression line, suggesting that they contained equal proportions of dominant and recessive alleles.
In summary, based on both the GCA and SCA effects, the study has identified suitable parents that can be used for an ELS resistance breeding program in West and central Africa. In addition, the study has revealed the existence of additive and dominance gene action for controlling ELS disease resistance with a predominance of additive gene action. This suggests that selection for the two ELS resistance traits could be easily practiced in early generations to develop ELS-resistant lines. Effective early generation selection for resistance would be advantageous and allow for procedures such as independent culling, tandem selection, or index selection involving other traits, such as yield, seed quality, and multiple pest resistance [
26]. Earlier studies on early generation selection for yield and ELS indicated a selection advantage among crosses compared to individual plant selection or within-family selection [
35,
36]. However, the positive correlation of ELS resistance traits with yield and its components reported elsewhere [
12,
32,
34,
37,
38] requires developing large number of crosses to break the undesirable association, and identify high-yielding genotypes with acceptable ELS resistance.
5. Conclusions
The study, using populations developed from varieties in West and Central Africa, was useful to understanding the mode of inheritance, identifying good combiners, and evaluating the breeding potential of populations for ELS resistance. The results indicated more recessive alleles in the resistant parents, confirming the recessive inheritance of ELS. Both additive and dominance gene actions control ELS disease resistance, with a predominance of additive gene action. The results suggest that selection of appropriate parents for breeding programs for ELS resistance should be based on both GCA and SCA effects. NAMA, PC79-79, and B188 are good combiners for use in breeding for ELS resistance in the region. The best crosses, including NAMA × PC79-79, B188 × PC79-79, B188 × QH243C, and NAMA × CN94C, were identified for further evaluation.





{kind=link}
{kind=link}