Biostimulants for Plant Growth Promotion and Sustainable Management of Phytoparasitic Nematodes in Vegetable Crops



Abstract
:1. Introduction
2. The State-of-the-Art
2.1. The Market Supply
2.2. The Literature Review
3. An Experimental Case Study
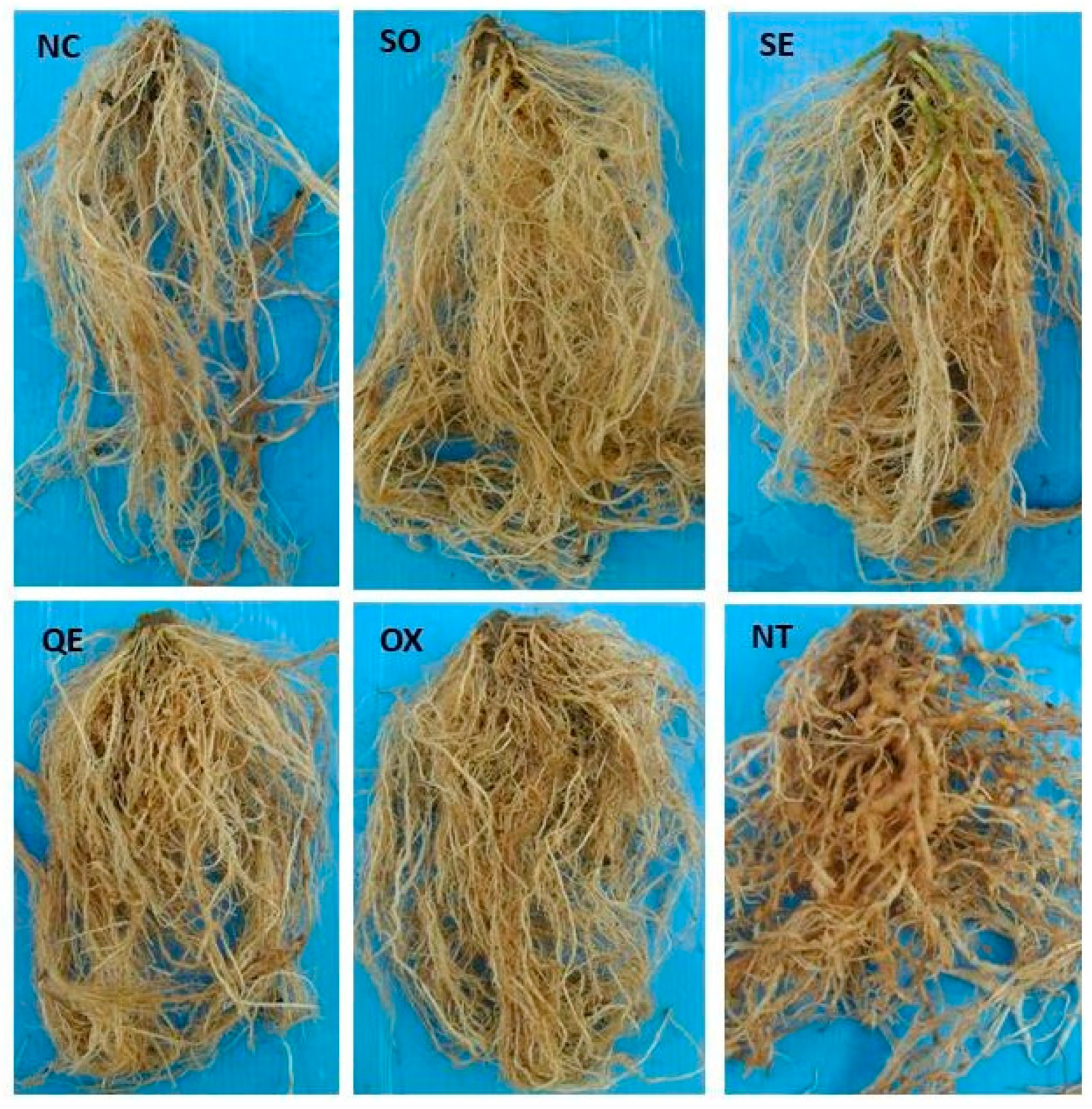
3.1. Materials and Methods
3.2. Results
4. Discussion
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Nicol, J.M.; Turner, S.J.; Coyne, D.L.; Den Nijs, L.; Hockland, S.; Maafi, Z.T. Current nematode threats to world agriculture. In Genomics and Molecular Genetics of Plant-Nematode Interactions; Jones, J., Gheysen, G., Fenoll, C., Eds.; Springer: Dordrecht, NL, USA, 2011; pp. 21–43. [Google Scholar]
- Sikora, R.A.; Fernandez, E. Nematode parasites of vegetables. In Plant Parasitic Nematodes in Subtropical and Tropical Agriculture; Luc, M., Sikora, R.A., Bridge, J., Eds.; CABI: Wallingford, UK, 2005; pp. 319–392. [Google Scholar]
- Le Mire, G.; Nguyen, M.L.; Fassotte, B.; du Jardin, P.; Verheggen, F.; Delaplace, P.; Jijakli, H. Implementing plant biostimulants and biocontrol strategies in the agroecological management of cultivated ecosystems, A review. Biotechnol. Agron. Soc. Environ. 2016, 20, 299–313. [Google Scholar]
- Brown, P.; Saa, S. Biostimulants in agriculture. Front. Plant Sci. 2015, 6, 671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basak, A. Biostimulators. Definitions, classification and legislation. In Biostimulators in Modern Agriculture: General Aspects; Gawrońska, H., Ed.; Wie Jutra: Warsaw, Poland, 2008; pp. 7–17. [Google Scholar]
- Askary, T.H.; Martinelli, P.R.P. Biocontrol Agents of Phytonematodes; CAB International: Wallingford, UK, 2015; p. 470. [Google Scholar]
- D’Addabbo, T.; Laquale, S.; Lovelli, S.; Candido, V.; Avato, P. Biocide plants as a sustainable tool for the control of pests and pathogens in vegetable cropping systems. Ital. J. Agron. 2014, 9, 137–145. [Google Scholar] [CrossRef]
- Oka, Y. Mechanisms of nematode suppression by organic soil amendments—A review. Appl. Soil Ecol. 2010, 44, 101–115. [Google Scholar] [CrossRef]
- European Union. Regulation 2019/1009, 2019. (EU) of the European Parliament and of the Council of 5 June 2019 Laying Down Rules on the Making Available on the Market of EU Fertilising Products and Amending Regulations (EC) No 1069/2009 and (EC) No 1107/2009 and Repealing Regulation (EC) No 2003/2003. OJ L 170, 25.6.2019. pp. 1–114. Available online: http://data.europa.eu/eli/reg/2019/1009/oj (accessed on 6 August 2019).
- D’Addabbo, T.; Radicci, V.; Lucarelli, G.; Carella, A.; Bernad, D.; Martin, E. Effectiveness of a formulation from Pedaliaceae plants (Nematon® EC) for the control of the root-knot nematode Meloidogyne incognita on greenhouse tomato. Acta Hortic. 2011, 914, 233–236. [Google Scholar] [CrossRef]
- Giannakou, I.O. Efficacy of a formulated product containing Quillaja saponaria plant extracts for the control of root-knot nematodes. Eur. J. Plant Pathol. 2011, 130, 587–596. [Google Scholar] [CrossRef]
- D’Addabbo, T.; Curto, G.; Greco, P.; Di Silvestro, D.; Coiro, M.I.; Lamberti, F.; Ferrari, V.; Santi, R.; Carella, A. Preliminary trials with extracts of Quillaja saponaria Molina for the control of root-knot nematodes. Nematol. Medit. 2005, 33, 29–34. [Google Scholar]
- Curto, G.; Dallavalle, E.; Matteo, R.; Lazzeri, L. Biofumigant effect of new defatted seed meals against the southern root-knot nematode, Meloidogyne incognita. Ann. Appl. Biol. 2016, 169, 1–10. [Google Scholar] [CrossRef]
- Rizvi, R.; Singh, G.; Safiuddin; Ali Ansari, R.; Ali Tiyagi, S.; Mahmood, I. Sustainable management of root-knot disease of tomato by neem cake and Glomus fasciculatum. Cogent Food Agric. 2015, 1, 1008859. [Google Scholar] [CrossRef]
- Abbasi, P.A.; Riga, E.; Conn, K.L.; Lazarovits, G. Effect of neem cake soil amendment on reduction of damping-off severity and population densities of plant-parasitic nematodes and soilborne plant pathogens. Can. J. Plant Pathol. 2005, 27, 38–45. [Google Scholar] [CrossRef]
- Khan, S.A.; Abid, M.; Hussain, F. Nematicidal activity of seaweeds against Meloidogyne javanica. Pak. J. Nematol. 2015, 33, 195–203. [Google Scholar]
- Rizvi, M.A.; Shameel, M. In vitro nematicidal activities of seaweed extracts from Karachi coast. Pak. J. Bot. 2006, 38, 1245–1248. [Google Scholar]
- Whapham, C.A.; Jenkins, T.; Blunden, G.; Hankins, S. The role of seaweed extracts, Ascophyllum nodosum, in the reduction in fecundity of Meloidogyne javanica. Fundam. Appl. Nematol. 1994, 17, 181–183. [Google Scholar]
- Wu, Y.; Jenkins, T.; Blunden, G.; Whapham, C.; Hankins, S.D. The role of betaines in alkaline extracts of Ascophyllum nodosum in the reduction of Meloidogyne javanica and M. incognita. Fundam. Appl. Nematol. 1997, 20, 99–102. [Google Scholar]
- Featonby-Smjth, B.C.; Van Staden, J. The effect of seaweed concentrate on the growth of tomato plants in nematode infested soil. Sci. Hortic. 1983, 20, 137–146. [Google Scholar] [CrossRef]
- El-Ansary, M.S.M.; Hamouda, R.A. Biocontrol of root-knot nematode infected banana plants by some marine algae. Russ. J. Mar. Biol. 2014, 40, 140–146. [Google Scholar] [CrossRef]
- Paracer, S.; Armin, C.; Tarjan, A.C.; Hodgson, L.M. Effective use of marine algal products in the management of plant parasitic nematodes. J. Nematol. 1987, 19, 194–200. [Google Scholar]
- Naqvi, B.S.; Khan, A.; Shaikh, D.; Shaikh, M.R. Nematicidal properties of selected marine algae from Karachi coast. J. Islam. Acad. Sci. 1992, 5, 171–172. [Google Scholar]
- De Waele, D.; McDonald, A.H.; De Waele, E. Influence of seaweed concentrate on the reproduction of Pratylenchus zeae (Nematoda) on maize. Nematologica 1988, 34, 71–77. [Google Scholar]
- Morgan, K.T.; Tarjan, A.C. Management of sting nematode on centipedegrass with kelp extracts. Proc. Flor. State Hortic. Soc. 1980, 93, 97–99. [Google Scholar]
- Tarjan, A.C. Kelp derivatives for nematode-infected citrus trees. J. Nematol. 1977, 9, 287. [Google Scholar]
- Radwan, M.A.; Farrag, S.A.; Abu-Elamayem, M.M.; Ahmed, N.S. Extraction, characterization, and nematicidal activity of chitin and chitosan derived from shrimp shell wastes. Biol. Fert. Soils 2012, 48, 463–468. [Google Scholar] [CrossRef]
- Escudero, N.; Lopez-Moya, F.; Ghahremani, Z.; Zavala-Gonzalez, E.A.; Alaguero-Cordovilla, A.; Ros-Ibañez, C.; Lacasa, A.; Sorribas, F.J.; Lopez-Llorca, L.V. Chitosan increases tomato root colonization by Pochonia chlamydosporia and their combination reduces root-knot nematode damage. Front. Plant Sci. 2017, 8, 1415. [Google Scholar] [CrossRef] [PubMed]
- Asif, M.; Ahmad, F.; Tariq, M.; Khan, A.; Ansari, T.; Khan, F.; Siddiqui, A.M. Potential of chitosan alone and in combination with agricultural wastes against the root-knot nematode, Meloidogyne incognita infesting eggplant. J. Plant Prot. Res. 2017, 57, 288–295. [Google Scholar] [CrossRef]
- Mota, L.C.; dos Santos, M.A. Chitin and chitosan on Meloidogyne javanica management and on chitinase activity in tomato plants. Trop. Plant Pathol. 2016, 41, 84–90. [Google Scholar] [CrossRef]
- Liang, W.; Yu, A.; Wang, G.; Zheng, F.; Jia, J.; Xu, H. Chitosan-based nanoparticles of avermectin to control pine wood nematodes. Int. J. Biol. Macromol. 2018, 112, 258–263. [Google Scholar] [CrossRef]
- Mwaheb, M.A.; Hussain, M.; Tian, J.; Zhang, X.; Hamid, M.I.; El-Kassim, N.A.; Hassan, G.M.; Xiang, M.; Liu, X. Synergetic suppression of soybean cyst nematodes by chitosan and Hirsutella minnesotensis via the assembly of the soybean rhizosphere microbial communities. Biol. Contr. 2017, 115, 86–94. [Google Scholar] [CrossRef]
- Nunes da Silva, M.; Cardoso, A.R.; Ferreira, D.; Brito, M.; Pintado, M.E.; Vasconcelos, M.W. Chitosan as a biocontrol agent against the pinewood nematode (Bursaphelenchus xylophilus). For. Pathol. 2014, 44, 420–423. [Google Scholar] [CrossRef]
- Candido, V.; Campanelli, G.; D’Addabbo, T.; Castronuovo, D.; Perniola, M.; Camele, I. Growth and yield promoting effect of artificial mycorrhization on field tomato at different irrigation regimes. Sci. Hort. 2015, 187, 35–43. [Google Scholar] [CrossRef]
- Colella, T.; Candido, V.; Campanelli, G.; Camele, I.; Battaglia, D. Effect of irrigation regimes and artificial mycorrhization on insect pest infestations and yield in tomato crop. Phytoparasitica 2014, 42, 235–246. [Google Scholar] [CrossRef]
- Flor-Peregrín, E.; Verdejo-Lucas, S.; Talavera, M. Combined use of plant extracts and arbuscular mycorrhizal fungi to reduce root-knot nematode damage in tomato. Biol. Agric. Hortic. 2017, 33, 115–124. [Google Scholar] [CrossRef]
- Flor-Peregrín, E.; Azcón, R.; Martos, V.; Verdejo-Lucas, S.; Talavera, M. Effects of dual inoculation of mycorrhiza and endophytic, rhizospheric or parasitic bacteria on the root-knot nematode disease of tomato. Biocontr. Sci. Technol. 2014, 24, 1122–1136. [Google Scholar] [CrossRef]
- Affokpon, A.; Coyne, D.L.; Lawouin, L.; Tossou, C.; Agbèdè, R.D.; Coosemans, J. Effectiveness of native West African arbuscular mycorrhizal fungi in protecting vegetable crops against root-knot nematodes. Biol. Fertil. Soils 2011, 47, 207–217. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, J.; Christie, P.; Li, X. Pre-inoculation with arbuscular mycorrhizal fungi suppresses root knot nematode (Meloidogyne incognita) on cucumber (Cucumis sativus). Biol. Fertil. Soils 2008, 45, 205. [Google Scholar] [CrossRef]
- Marro, N.; Lax, P.; Cabello, M.; Doucet, M.E.; Becerra, A.G. Use of the arbuscular mycorrhizal fungus Glomus intraradices as biological control agent of the nematode Nacobbus aberrans parasitizing tomato. Braz. Arch. Biol. Technol. 2014, 57, 668–674. [Google Scholar] [CrossRef]
- Jonathan, E.I.; Cannayane, I.; Samiyappan, R. Field application of biocontrol agents for the management of spiral nematode, Helicotylenchus multicinctus, in banana. Nematol. Mediterr. 2004, 32, 169–173. [Google Scholar]
- Radwan, M.A.; Farrag, S.A.A.; Abu-Elamayem, M.M.; Ahmed, N.S. Biological control of the root-knot nematode, Meloidogyne incognita on tomato using bioproducts of microbial origin. Appl. Soil Ecol. 2012, 56, 58–62. [Google Scholar] [CrossRef]
- El-Haddad, M.E.; Mustafa, M.I.; Selim, S.M.; El-Tayeb, T.S.; Mahgoob, A.E.; Aziz, N.H. The nematicidal effect of some bacterial biofertilizers on Meloidogyne incognita in sandy soil. Braz. J. Microbiol. 2011, 42, 105–113. [Google Scholar] [CrossRef]
- Ismail, A.E.; Hasabo, S.A. Evaluation of some new Egyptian commercial biofertilizers, plant nutrients and a biocide against Meloidogyne incognita root knot nematode infecting sunflower. Pak. J. Nematol. 2000, 18, 39–49. [Google Scholar]
- Biliavska, L.O.; Tsygankova, V.A.; Kozyritska, V.E.; Iutynska, G.O.; Andrusevich, Y.V.; Babich, O.A. Application of new microbial plant resistance/plant growth protection inducers for increasing Chinese cabbage plant tolerance against parasitic nematode Heterodera schachtii Schmidt. Int. J. Res. Biosci. 2016, 5, 64–82. [Google Scholar]
- Khan, Z.; Tiyagi, S.A.; Mahmood, I.; Rizvi, R. Effects of N fertilization, organic matter, and biofertilizers on the growth and yield of chilli in relation to management of plant-parasitic nematodes. Turk. J. Bot. 2012, 36, 73–78. [Google Scholar]
- Bridge, J.; Page, S.L.J. Estimation of root-knot nematode infestation levels on roots using a rating chart. Int. J. Pest Manag. 1980, 26, 296–298. [Google Scholar] [CrossRef]
- Hussey, P.S.; Barker, K.R. A comparison of methods of collecting inocula of Meloidogyne spp., including a new technique. Plant Dis. Rep. 1973, 57, 1025–1028. [Google Scholar]
- Chitwood, J.D. Phytochemical based strategies for nematode control. Ann. Rev. Phytopathol. 2002, 40, 21–49. [Google Scholar] [CrossRef]
- Verkleij, F.N. Seaweed extracts in agriculture and horticulture: A review. Biol. Agric. Hort. 1992, 8, 309–324. [Google Scholar] [CrossRef]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in plant science: A global perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef]
- Oliveira, R.D.; Dhingra, O.D.; Lima, A.O.; Jham, G.N.; Berhow, M.A.; Holloway, R.K.; Vaughn, S.F. Glucosinolate content and nematicidal activity of Brazilian wild mustard tissues against Meloidogyne incognita in tomato. Plant Soil 2011, 341, 155–164. [Google Scholar] [CrossRef]
- Pretali, L.; Bernardo, L.; Butterfield, T.S.; Trevisan, M.; Lucini, L. Botanical and biological pesticides elicit a similar induced systemic response in tomato (Solanum lycopersicum) secondary metabolism. Phytochemistry 2016, 130, 56–63. [Google Scholar] [CrossRef]
- Bargiacchi, E.; Miele, S.; Romani, A.; Campo, M. Biostimulant activity of hydrolyzable tannins from sweet chestnut (Castanea sativa mill.). Acta Hortic. 2013, 1009, 111–116. [Google Scholar] [CrossRef]
- Sofo, A.; Nuzzaci, M.; Vitti, A.; Tataranni, G.; Scopa, A. Control of biotic and abiotic stresses in cultivated plants by the use of biostimulant microorganisms. In Improvement of Crops in the Era of Climatic Changes; Ahmad, P., Wani, M.R., Azooz, M.M., PhanTran, L.S., Eds.; Springer Science+Business Media: New York, NY, USA, 2014; pp. 107–117. [Google Scholar]
- Vos, C.; Schouteden, N.; Van Tuinen, D.; Chatagnier, O.; Elsen, A.; De Waele, D.; Gianinazzi-Pearson, V. Mycorrhiza-induced resistance against the root–knot nematode Meloidogyne incognita involves priming of defense gene responses in tomato. Soil Biol. Biochem. 2013, 60, 45–54. [Google Scholar] [CrossRef]
- Mahanty, T.; Bhattacharjee, S.; Goswami, M.; Bhattacharyya, P.; Das, B.; Ghosh, A.; Tribedi, P. Biofertilizers: A potential approach for sustainable agriculture development. Env. Sci. Poll. Res. 2017, 24, 3315–3335. [Google Scholar] [CrossRef] [PubMed]
- Schouteden, N.; De Waele, D.; Panis, B.; Vos, C.M. Arbuscular mycorrhizal fungi for the biocontrol of plant-parasitic nematodes: A review of the mechanisms involved. Front. Microbiol. 2015, 6, 1280. [Google Scholar] [CrossRef] [PubMed]
- Youssef, M.M.; Eissa, M.F. Biofertilizers and their role in management of plant parasitic nematodes, A review. J. Biotechnol. Pharm. Res. 2014, 5, 1–6. [Google Scholar]
- Vasyukova, N.I.; Zinov’Eva, S.V.; Il’Inskaya, L.I.; Perekhod, E.A.; Chalenko, G.I.; Gerasimova, N.G.; Il’ina, A.V.; Varlamov, V.P.; Ozeretskovskaya, O.L. Modulation of plant resistance to diseases by water-soluble chitosan. Appl. Biochem. Microbiol. 2001, 37, 103–109. [Google Scholar] [CrossRef]
- Reichert, C.L.; Salminen, H.; Weiss, J. Quillaja Saponin Characteristics and Functional Properties. Ann. Rev. Food Sci. Technol. 2019, 10, 43–73. [Google Scholar] [CrossRef]
- Sharma, S.H.S.; Lyons, G.; McRoberts, C.; McCall, D.; Carmichael, E.; Andrews, F.; McCormack, R. Brown seaweed species from Strangford Lough: Compositional analyses of seaweed species and biostimulant formulations by rapid instrumental methods. J. Appl. Phycol. 2012, 24, 1141–1157. [Google Scholar]
- Dong, L.Q.; Zhang, K.Q. Microbial control of plant-parasitic nematodes: A five-party interaction. Plant Soil 2006, 288, 31–45. [Google Scholar] [CrossRef]
- Khalil, M.S.; Badawy, M.E. Nematicidal activity of a biopolymer chitosan at different molecular weights against root-knot nematode, Meloidogyne incognita. Plant Prot. Sci. 2012, 48, 170–178. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Commercial Name | Formulation 1 | Raw Materials | Activity 2 |
|---|---|---|---|
| Aegis ™ | P | Micorrhizal fungi | 1, 4, 5 |
| Alg-a-Mic ™ | L | Seaweed extract | 1, 4, 5, 7 |
| Algafit ™ | L | Seaweed extract | 1, 4 |
| Ascogreen ™ | L | Seaweed extract | 1, 4 |
| Biofence ™ | P | Brassica meal | 1, 3, 5, 6 |
| Biofence 10 ™ | P | Brassica meal | 1, 3, 5, 6 |
| Biofence FL ™ | L | Brassica extract | 1, 2, 4, 6 |
| Bioki ™ | p | Neem oil | 1, 3, 7 |
| Cogisin ™ | L | Plant extracts | 1, 2, 4, 7 |
| Ecoessen NP ™ | P | Bone meal, neem cake | 1, 3, 6 |
| Ekoprop Nemax ™ | P | Mycorrhizal fungi | 1, 2, 4 |
| Ergo Bio ™ | L | Humic and fulvic acids | 1, 3, 4, 5, 8 |
| Ergon ™ | L | Seaweed extract | 1, 4 |
| Fertineem ™ | L | Neem oil | 1, 4 |
| Force 4 ™ | L | Seaweed extract | 1, 2, 4, 5 |
| Hunter ™ | L | Plant extracts | 1, 4 |
| Ilsaneem ™ | P | Neem cake | 1, 2, 3, 7 |
| Kendal Nem ™ | L | Plant extracts | 1, 2, 4, 6 |
| Keos Guardian ™ | L | Chitosan | 1, 5 |
| Micofort ™ | P | Micorrhizal fungi | 1, 2, 4, 5 |
| Micosat F ™ | P | Micorrhizal fungi | 1, 4, 5 |
| Micosat Jolly ™ | P | Micorrhizal fungi | 1, 4, 5 |
| Mychodeep ™ | P | Micorrhizal fungi | 1, 2, 4 |
| Neem Soil ™ | P | Neem cake | 1, 3, 4, 6 |
| Neem Care FL ™ | L | Plant extracts | 1, 2, 4 |
| Nema 300 WW ™ | L | Plant oils | 1, 2, 4 |
| Nemaforce ™ | L | Humic and fulvic acids, plant extracts | 1, 2, 4, 5, 7 |
| Nematec ™ | L | Seaweed extract | 1, 2, 4 |
| Nematiller ™ | L | Plant extracts | 1, 2, 4 |
| Nematon EC ™ | L | Sesame oil | 1, 4 |
| NeMax ™ | L | Sesame oil | 1, 4 |
| Nutrich ™ | P | Neem and pongamia cake | 1, 2, 3, 4 |
| Propoli oleoso ™ | L | Propolis oil | 1, 4 |
| Rigenera Active | L | Seaweed macerate, plant extracts | 1, 2, 4 |
| Sesamin EC ™ | L | Sesame oil | 2, 4, 5 |
| Tagete ™ | L | Tagetes extract | 1, 2, 4, 8 |
| Tequil Multi ™ | L | Quillay and yucca extracts | 1, 2, 4, 8 |
| Tyson ™ | L | Propolis oil | 1, 4, 8 |
| Xedaneem ™ | P | Neem cake | 1, 6 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Addabbo, T.; Laquale, S.; Perniola, M.; Candido, V. Biostimulants for Plant Growth Promotion and Sustainable Management of Phytoparasitic Nematodes in Vegetable Crops. Agronomy 2019, 9, 616. https://doi.org/10.3390/agronomy9100616
D’Addabbo T, Laquale S, Perniola M, Candido V. Biostimulants for Plant Growth Promotion and Sustainable Management of Phytoparasitic Nematodes in Vegetable Crops. Agronomy. 2019; 9(10):616. https://doi.org/10.3390/agronomy9100616
Chicago/Turabian StyleD’Addabbo, Trifone, Sebastiano Laquale, Michele Perniola, and Vincenzo Candido. 2019. "Biostimulants for Plant Growth Promotion and Sustainable Management of Phytoparasitic Nematodes in Vegetable Crops" Agronomy 9, no. 10: 616. https://doi.org/10.3390/agronomy9100616
APA StyleD’Addabbo, T., Laquale, S., Perniola, M., & Candido, V. (2019). Biostimulants for Plant Growth Promotion and Sustainable Management of Phytoparasitic Nematodes in Vegetable Crops. Agronomy, 9(10), 616. https://doi.org/10.3390/agronomy9100616