1. Introduction
Cotton (
Gossypium hirsutum L.) is a natural white fiber and cash crop that is grown globally [
1]. The cotton plant is characterized by indeterminate growth habits and shows morphological and physiological adaptation to a wide range of environmental and management practices, including planting density and cultivar. An expanding population necessitates global efforts to increase crop production, especially those fulfilling food and fiber needs. Currently, numerous management practices have been introduced for cotton production systems, but lint production per unit area has remained stagnant [
2]. High input costs combined with multiple management and material inputs have threatened cotton productivity. [
2]. An efficient agricultural production system characterized by moderate planting density with one-time fertilization under a short growing season can reduce inputs without yield loss [
3,
4].
Planting density and choice of cultivar are important agronomic practices that have the potential to optimize the canopy photosynthetic rate and crop productivity of any cropping system [
5]. Changes in plant architecture and canopy dynamics in response to planting density can have impacts on disease incidence, water use, canopy temperature, and enzymatic activity of assimilate metabolism [
6]. Manipulations of planting density in cotton have remarkable impacts on biomass partitioning, nutrient uptake, boll distribution, changes in the light spectrum, and crop production [
3,
5,
7,
8], which can influence yield and profits for producers [
9]. Plants at high density can minimize evaporation and irrigation frequency, as well as increase the utilization of irrigation water [
10,
11]. In contrast, high-density planting can slow down leaf appearance and reduce open boll density [
12], boll weight, and boll number [
7]. It also delays leaf senescence [
13] and decreases nitrogen use efficiency and nitrogen recovery efficiency [
14]. A planting density of up to nine plants m
−2 has been reported to sustain leaf photosynthetic rate and reproductive organ biomass formation by increasing plant potassium uptake at various developmental stages. However, a sowing density of >10 plants m
−2 and subsequent shading can result in disease infestation, small boll size, fruit shedding, delayed maturity, and decreased individual plant development [
4]. Dense planting can also delay crop maturity by promoting vegetative growth and can substantially depress net photosynthetic rate [
4] due to decreased RuBP carboxylase activity and chlorophyll content [
15]. High planting density can increase the auxin (IAA) content and enhance auxin polar transport by increasing the expression of the auxin biosynthesis gene (Gh
YUC5) and the auxin polar transport gene (Gh
PIN1). It can also inhibit vegetative branching by decreasing IAA, cytokinin, gibberellic acid, and brassinosteroid contents, followed by increased strigolactone content due to differential expression of hormone-associated genes in the tips of vegetative branches [
15]. Optimal plant density can ensure healthy plant development by maintaining a core population of plants synchronizing boll number and fiber quality to achieve optimal yield [
16].
Leaf morphological and physiological attributes are important players in photosynthetic regulation [
4] and can provide a structural framework for gas exchange as well as optimize the photosynthetic function [
17]. Cotton leaf surface characteristics, including cuticular thickening, wax layer, and trichomes, play critical roles in the variability of optical properties [
18]. Generally, leaves developed under high sunlight can have thicker and smaller leaves with well-developed plastid tissues, greater stomatal density, and smaller granal stacks than shade leaves [
19]. Plants under low density planting have a lower chlorophyll content and a higher electron transfer rate and ribulose-1,5-bi-phasphate carboxylase/oxygenase compared with high-density planting [
20,
21]. Leaves developed under lower density (sun leaves) are tolerant to strong light; conversely, shade leaves have weak photoprotection potential and are more sensitive to high light [
17,
21].
Studies regarding cotton growth and lint yield in response to diverse populations are common [
22,
23,
24]. However, we are the first to report the effects of changing the planting density on cotton lint yield, leaf structure, chlorophyll fluorescence, and leaf gas exchange characteristics in subtropical regions. The objectives of this study were to investigate leaf structural and functional characteristics in response to different planting densities and varieties. It also explored optimal plant density and variety for improved lint yield in subtropical regions.
2. Materials and Methods
2.1. Plant Material and Experimental Site
Seeds of two cotton cultivars—V1 (Zhongmian-16) and V2 (J-4B)—were procured from the Cotton Research Institute, Chinese Academy of Agricultural Sciences and were grown under field conditions for two years. A replicated two-year (2017 and 2018 growing seasons) field experiment was conducted at Guangxi University, Nanning, China. The soil properties of the experimental field were sandy loam and yellowish, having a pH of 6.5; organic matter of 23.37 mg kg−1; and available nitrogen, phosphorus, and potassium content of 53.24, 77.58, and 6.30 mg kg−1, respectively. The experimental design layout was a balanced split plot with three replications.
2.2. Crop Management and Experimental Design
Before sowing, the experimental field was ploughed, laser leveled approximately three weeks prior, and covered with plastic film to conserve moisture and suppress weed germination. The experiments were designed in a split plot arrangement with three replications of each of the six treatment combinations. Two cotton varieties (V1, Zhongmian-16; V2, J-4B), were randomly allocated to the main plots and three plant population levels (D1, low, 3 × 104; D2, medium, 6 × 104; and D3, dense, 9 × 104 ha−1) were randomized in subplots. By increasing the precision of comparisons, split plot arrangements were adopted. Seeds were sown on 5 June in double rows on each raised plot (3.0 m wide and 11 m long), with a total plot size of 33.0 m2. Each subplot was 11 m long and 1.5 m wide, consisting of four rows with narrow (10 cm) and wide (66 cm) row spaces for a total of eight rows on each main plot. Plant-to-plant spacing was controlled according to the corresponding population level. Crops were irrigated one day after sowing to ensure uniform germination. Cotton seedlings were hand-thinned at the third leaf stage to the target population level for each plot. A basal application of phosphorus (P2O5) at 66 kg ha−1, nitrogen (N) at 170 N kg ha−1, and potash (K2O) at 190 kg ha−1 was applied using superphosphate (12% P2O5), urea (46% N), and potassium chloride (59% K2O), respectively, during the pinhead stage. A plant growth regulator (i.e., mepiquat chloride) at the rate of 0.057 active ingredient ha−1 was sprayed to control vegetative growth. All the necessary field management practices were performed according to crop requirements during the whole crop cycle.
2.3. Data Collection
Data on leaf structure, chlorophyll fluorescence, leaf gas exchange attributes, cotton yield, and yield contributors were recorded for each treatment in three replications. The details of each measurement are given below.
2.4. Yield and Yield Components
To assess cotton yield, fully opened bolls were hand-picked at three times in each treatment. The harvested seed cotton was sun-dried to ≤11% moisture content [
16]. The seed cotton was ginned to calculate seed cotton and lint yield. During the second picking, 100 mature bolls were manually picked to calculate single boll mass and lint percentage. Seed cotton yield of 100 bolls was divided by the number of bolls to assess individual boll weight. Lint % was determined using the lint yield of 100 bolls and divided by seed cotton mass.
2.5. Cotton Leaf Structure Attributes
Ten plants in each plot were randomly tagged to measure leaf structure and plant growth characteristics at the boll setting stage. Leaf thickness was determined on 10 fully expanded leaves from the upper part of three plants (functional leaves, i.e., upper fourth leaf). A hand-held micrometer (Mitutoyo Digital Micrometer Model 293-185, Kawasaki, Japan) with a digital display and a clutch that ensured uniform pressure [
25] was used for leaf thickness assessment. A 5 × 8 mm leaf section was removed for each treatment. Samples were then added into 10 mL tubes containing 50%, 5%, and 5% alcohol solution, formaldehyde, and glacial acetic acid, respectively. Scanning electron microscopy was performed at Guanxi Medical University using a SUPRA 55VP (Carl Zeiss AG, Oberkochen, Germany). Image software was used to assess cotton leaf stomatal length, width, density, and pore perimeter according to the method reported in [
26].
2.6. Chlorophyll Fluorescence Traits
Cotton leaf chlorophyll fluorescence attributes were measured on a fully expanded functional leaf (upper fourth leaf on the main stem) on a sunny day (between 1000 and 1200 h) via a portable mini PAM-2100 fluorometer coupled with a 2030-B leaf (Walz, Germany). Maximum (
Fm) and minimum (
Fo) fluorescence values of dark-adapted leaves (photosystem II (PSII) centers open) were measured using leaf clips. The maximum and minimum fluorescence values were assessed at 0.5 µmol m
−2 s
−1 with a frequency of 0.6 kHz and a 0.8 saturating pulse at >8000 µmol m
−2 s
−1, respectively. Maximum quantum yield of PSII photochemistry
(Fv/
Fm) was calculated as
Fv/
Fm = 1 − (
Fo/
Fm) [
27]. The effective quantum yield of PSII photochemistry of light-adapted leaves was determined by Φ
PSII (
Fm’ −
F)/
Fm’ [
28]. Coefficient of photochemical quenching (qP) was assessed using the formula qP
= (
Fm’ −
Fs)/(
Fm’ −
Fo’) [
29]. Minimal fluorescence of light-adapted leaves (
Fo’) was calculated according to the equation
Fo’ = Fo/(
Fv/
Fm +
Fo/
Fm’) done by [
28]. Nonphotochemical quenching (NPQ) was recorded according to [
6] as NPQ
= (
Fm −
Fm’)/
Fm’, where
Fm represents the value of the predawn observations. The electron transport rate (ETR) was assessed using a leaf absorptance of 0.85 and half of the absorbed light was partitioned to each photosystem: ETR = PSII × PPFD × 0.85 × 0.5 [
30].
2.7. Leaf Gas Exchange Attributes
At squaring, flowering, peak bloom, and boll setting stages, fully expanded leaves from the upper part of three plants (functional leaves, i.e., upper fourth leaf) were chosen to assess net photosynthetic rate (Pn), stomatal conductance (gs), intercellular CO2 concentration (Ci), and transpiration rate (E). Net rate of photosynthesis was measured from the six functional leaves of three plants in each treatment using a portable infrared gas exchange analyzer (Li-6400, Li-Cor, Lincoln, NE, USA). These observations were made on a clear day between 10:00 a.m. and 12:00 p.m. Beijing time in each experimental unit of four replications. Leaves in each plot followed the following adjustments: PAR, 1800 μmol m−2 s−2; air flow, 389.42 mmol−1 m−2 s−1; water vapor pressure into leaf chamber, 3.13 mbar; leaf temperature, 30 °C; ambient temperature, 33.69 °C; and ambient carbon dioxide concentration, 330–350 mol mol−1.
2.8. Statistical Analysis
All the data were processed using Microsoft Excel 2016. Figures were plotted using Sigma Plot 14.00 software. Analysis of variance was implemented using SAS software (version 8.1, SAS Institute, Cary, NC, USA). The initial combined data showed no interactions with years. Therefore, the data were pooled and presented across the two years. Means of planting density were separated using the least significant difference (LSD) test at the 5% probability level. Both planting densities and cultivars were taken as main factors and fixed effects with cropping season as the repetitive measured factor with a fixed effect. Similarly, the interaction was taken as fixed effects and treatment × replication interaction, which was taken as a random effect. Differences among treatments imply statistical difference (p = 0.05).
4. Discussion
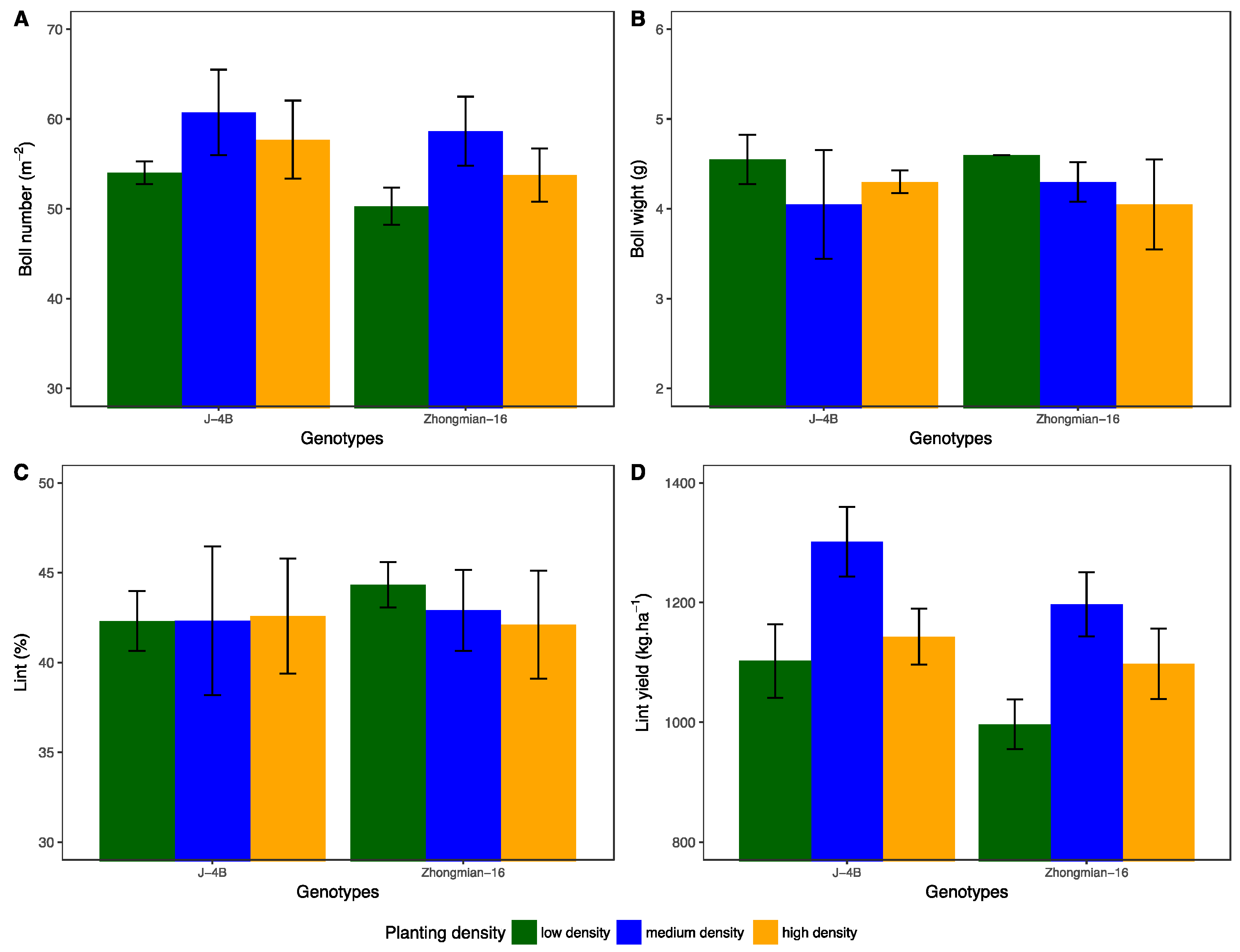
The current study has provided new data on the common perception that high planting density significantly decreases leaf structural characteristics, such as stomatal density, length, width, pore perimeter, and leaf thickness, as well as functional traits (leaf gas exchange and chlorophyll fluorescence traits), which leads to lint yield loss. However, we found that improved leaf functional and structural traits for J-4B under moderate density had a higher lint yield. Under high-density treatment, reductions in lint yield for Zhongmian-16 were associated with repression in leaf structural and functional attributes, which in turn caused depression in leaf photosynthetic capacity due to nutrient competition. The difference between varieties from changing planting density might be associated with canopy architecture and genetic variation. Therefore, these changes in varieties might have significant impacts on leaf structural and functional attributes and, ultimately, on yield formation.
High planting density responses to cotton lint yield, growth, biomass production, nutrient uptake, and fiber quality have been extensively investigated [
3,
4,
13,
22]. The mechanisms of interplant competitiveness under low-to-high planting density on leaf structure, chlorophyll fluorescence, and leaf gas exchange attributes for optimal cotton lint yield have not yet been reported. Across densities, the moderate population had a higher boll number m
−2 with improved lint yield for J-4B compared with Zhongmian-16 across two years. High-density plants substantially reduced yield and yield components in both years, probably due to competition for nutrients. The phenomenon of increased lint yield under moderate density can be associated with improved leaf structural and chlorophyll fluorescence traits and higher leaf photosynthetic capacity, which resulted in higher boll density m
-2 compared with other densities.
Moderate density favors dry matter partitioning to the reproductive structures rather than vegetative organs [
31] and less fruit shedding compared with denser plants. The reductions in lint yield under high density can be attributed to decreased leaf structural and physiological traits, which were observed in this study. The differences that existed between varieties for yield when changing planting density might be attributable to canopy architecture. Differences in plant canopy architectural traits among varieties have an impact on growth characteristics and lint yield. These data further confirmed that an appropriate selection of variety and optimal density can contribute to successful cotton production. Reducing population density may also have other implications, such as decreased frequency and insecticide inputs per season without any yield loss to increase profit. Moreover, high plant density can substantially depress leaf structural and physiological attributes, which in turn cause a severe yield penalty.
Plants respond to ambient and management interventions via architectural and structural changes. Plant growth and leaf morphological attributes, including stomatal density, size, number of pores, width, length, and leaf thickness features, are pivotal windows regulating leaf photosynthetic capacity [
10,
25] and offer a structural framework for CO
2 exchange and optimization of photosynthetic activities, which in turn can improve crop yield [
17]. In this study, high planting density substantially decreased leaf thickness, stomatal density, leaf length, width, and number of stomatal pores. Limitations in these attributes disrupted the photosynthetic capacity of plants by restricting entry of CO
2 to the mesophyll through the stomata of leaves, which is extremely responsive to light environments. Thus, the exchange of CO
2 by means of stomata might be restricted [
32]. Higher stomatal density, thicker leaves, and rapid metabolite transfer between the mesophyll and bundle sheet cells can favor higher leaf photosynthetic capacity [
33]. Increasing planting density has been proposed to decrease the stomatal density of wheat leaves [
34]. A greater stomatal size can facilitate CO
2 distribution into the leaf due to its conductance being proportional to the square of the effective radius of the stomatal pore, resulting in increased stomatal conductance [
35]. However, the responses of leaf structural attributes vary under different abiotic stresses in different plant species or varieties [
36]. These data suggest that plants under high-density conditions have significantly decreased leaf morphological characteristics, which might be particularly responsible for depressing leaf photosynthetic capacity.
Chlorophyll fluorescence is a nondestructive evaluation of PSII activity. In plant physiology, this technique is commonly used and has become a classical method for crop improvement, screening of beneficial traits, and linking genomic knowledge to phenological response. Due to the sensitivity of PSII to undesirable ambient conditions, this is a useful method for understanding photosynthetic mechanisms and a good indicator of how plants respond to ambient change [
37,
38].
ΦPSII is a measure of light energy capture efficiency, which reflects the actual primary sunlight energy conversion efficiency of the PSII reaction center [
15]. In this study, ΦPSII substantially declined under high-density conditions. Probably, a lower ΦPSII value under high-density conditions did not efficiently convert photon energy to chemical energy; however, this phenomenon needs further exploration. Under shading conditions, a low ΦPSII may be responsible for depressing Pn due to the adjustment in photochemical reaction centers [
39], which was observed in our study. The efficient use of limited light energy and the degree of the PSII reaction center openness can increase, resulting in improved energy conversion efficiency. This is associated with the increase of
Fv/
Fm, ΦPSII, and qP at early shading [
40]. The maximal photochemical efficiency of PSII (
Fv/
Fm) determines the potential quantum efficiency of PSII [
41]. In this study,
Fv/
Fm had higher values under low rather than high planting density, which is consistent with [
32], and reductions in
Fv/
Fm values might be due to the lower values of
Fm and increased values of
Fo. The ETR is an important chlorophyll fluorescence attribute affected by the external light environment. The rate of ETR declined from low to high density in this study, which corresponds with [
40], and shading can significantly decrease ETR values by affecting PSII photochemical reaction centers and consequently diminish the primary stable quinine acceptor of PSII, leading to a decrease in the activity of photosynthetic electron transport efficiency via PSII [
27,
33]. NPQ can have critical roles in the nonradiative dissipation of surplus light energy [
42]. A low-light environment can cause a reduction in NPQ, possibly associated with reduced light energy [
32,
43]. In this study, a severe decline in NPQ values was noted under high-density compared with low-density crops. This can be explained as the decreased NPQ being associated with the decreased efficiency of photochemical reactions through the reduced fraction of incident light in photochemical energy utilization, which resulted in lower thermal dissipation in PSII [
44]. The rate of photochemical quenching (qP) under dense crops showed a substantial reduction compared with low and moderate densities. Probably, a low-light environment can cause reductions in the amount of pigment and the efficiency of photochemical energy conversion, resulting in the depressed quantum yield of PSII and decreased qP. The qP reflects the efficiency of light quantum harvested by PSII to chemical energy and represents the openness degree of the PSII reaction center, and a greater qP results in greater activity of electron transfer in PSII.
Leaf gas exchange traits can play a central role in biomass formation and the prime determination of cotton lint yield [
45]. High planting density results in rapid canopy closure and an increase in radiation interception, which reduces weed competition [
46], but this impedes leaf gas exchange traits, leading to yield loss [
47]. In the current study, cotton leaf gas exchange parameters were substantially depressed under close planting at different growth stages. Accordingly, high-density conditions resulted in reductions in leaf stomatal density, length, width, pores, and leaf thickness, probably due to mutual shading, which may be responsible for depressing stomatal conductance (
gs) and CO
2 uptake through the stomata, which in turn suppressed the photosynthetic capacity. Plants under high-density conditions can significantly decrease
gs and
Ci, which can negatively influence the photosynthetic system [
38]. The CO
2 concentration plays a central role in net photosynthetic rate (Pn), but this varies across species and ambient conditions [
48,
49]. The
gs might respond to alterations in Pn and thus prevents
Ci near saturation. The primary function of stomata is to avoid desiccation and enable the passage of CO
2. Stomata induce a substantial disruption in the CO
2 assimilation rate, which reduces more in C
4 than C
3 plants. The stomatal limitation of Pn is the role of stomatal resistance to contribute to “resistance” to CO
2 uptake and stomatal limitation in spite of a decline in
Ci [
50]. The higher transpiration (E) rates in low-density conditions may have been due to low mutual shading, which allowed rapid stomata opening. Our data showed that high plant density substantially decreased leaf thickness, stomatal density, width, length, and stomatal pores and resulted in lower
Ci and
gs, which in turn depressed leaf photosynthetic capacity.

 ,
, 




{kind=link}