Optimal LED Wavelength Composition for the Production of High-Quality Watermelon and Interspecific Squash Seedlings Used for Grafting


 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Germination
2.2. Growth Chamber and Light Conditions
2.3. Sampling and Measurements
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, J.M.; Kubota, C.; Tsao, S.J.; Bie, Z.; Hoyos Echevarria, P.; Morra, L.; Odag, M. Current status of vegetable grafting: Diffusion, grafting techniques, automation. Sci. Hortic. 2010, 127, 93–105. [Google Scholar] [CrossRef]
- Louws, F.J.; Rivard, C.L.; Kubota, C. Grafting fruiting vegetables to manage soilborne pathogens, foliar pathogens, arthropods and weeds. Sci. Hort. 2010, 127, 127–146. [Google Scholar] [CrossRef]
- Savvas, D.; Colla, G.; Rouphael, Y.; Schwarz, D. Amelioration of heavy metal and nutrient stress in fruit vegetables by grafting. Sci. Hort. 2010, 127, 156–161. [Google Scholar] [CrossRef]
- Schwarz, D.; Rouphael, Y.; Colla, G.; Venema, J.H. Grafting as a tool to improve tolerance of vegetables to abiotic stresses: Thermal stress, water stress and organic pollutants. Sci. Hort. 2010, 127, 162–171. [Google Scholar] [CrossRef]
- Bantis, F.; Koukounaras, A.; Siomos, A.; Menexes, G.; Koufakis, T. Quantitative criteria of watermelon and squash seedlings used as scion and rootstock during grafting. Acta. Hortic. 2019. Accept for publication. [Google Scholar]
- Bantis, F.; Koukounaras, A.; Siomos, A.; Menexes, G.; Dangitsis, C.; Kintzonidis, D. Assessing quantitative criteria for characterization of quality categories for grafted watermelon seedlings. Horticulturae 2019, 5, 16. [Google Scholar] [CrossRef] [Green Version]
- Pocock, T. Influence of light-emitting diodes (LEDs) on light sensing and signaling networks in plants. In Light Emitting Diodes for Agriculture; Dutta Gupta, S., Ed.; Springer: Singapore, 2017; pp. 37–58. [Google Scholar]
- Pattison, P.M.; Tsao, J.Y.; Brainard, G.C.; Bugbee, B. LEDs for photons, physiology and food. Nature 2018, 563, 493–500. [Google Scholar] [CrossRef]
- Massa, G.D.; Kim, H.-H.; Wheeler, R.M.; Mitchell, C.A. Plant productivity in response to LED lighting. HortScience 2008, 43, 1951–1956. [Google Scholar] [CrossRef]
- Dutta Gupta, S.; Agarwal, A. Artificial lighting system for plant growth and development: Chronological advancement, working principles, and comparative assessment. In Light Emitting Diodes for Agriculture; Dutta Gupta, S., Ed.; Springer: Singapore, 2017; pp. 1–25. [Google Scholar]
- Bantis, F.; Smirnakou, S.; Ouzounis, T.; Koukounaras, A.; Ntagkas, N.; Radoglou, K. Current status and recent achievements in the field of horticulture with the use of light-emitting diodes (LEDs). Sci. Hortic. 2018, 235, 437–451. [Google Scholar] [CrossRef]
- Chen, M.; Chory, J.; Fankhauser, C. Light signal transduction in higher plants. Annu. Rev. Genet. 2004, 38, 87–117. [Google Scholar] [CrossRef] [Green Version]
- Folta, K.M.; Childers, K.S. Light as a growth regulator: Controlling plant biology with narrow-bandwidth solid-state lighting systems. HortScience 2008, 43, 1957–1964. [Google Scholar] [CrossRef] [Green Version]
- Ouzounis, T.; Rosenqvist, E.; Ottosen, C.-O. Spectral effects of artificial light on plant physiology and secondary metabolism. Hortscience 2015, 50, 1128–1135. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Li, G.; Wangb, H.; Wang Deng, X. Phytochrome Signaling Mechanisms. Arabidopsis Book 2011, 9, e0148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fankhauser, C.; Chory, J. Light control of plant development. Annu. Rev. Cell Dev. Biol. 1997, 13, 203–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dougher, T.A.; Bugbee, B.G. Long-term blue light effects on the histology of lettuce and soybean leaves and stems. J. Am. Soc. Hortic. Sci. 2004, 129, 467–472. [Google Scholar] [CrossRef] [Green Version]
- De Carbonnel, M.; Davis, P.; Roelfsema, M.R.G.; Inoue, S.i.; Schepens, I.; Lariguet, P.; Geisler, M.; Shimazaki, K.i.; Hangarter, R.; Fankhauser, C. The Arabidopsis PHYTOCHROME KINASE SUBSTRATE2 Protein Is a Phototropin Signaling Element That Regulates Leaf Flattening and Leaf Positioning. Plant Physiol. 2010, 152, 1391–1405. [Google Scholar] [CrossRef] [Green Version]
- Casal, J.J. Photoreceptor signaling networks in plant responses to shade. Annu. Rev. Plant Biol. 2013, 64, 403–427. [Google Scholar] [CrossRef]
- Wang, Y.; Folta, K.M. Contributions of green light to plant growth and development. Am. J. Bot. 2013, 100, 70–78. [Google Scholar] [CrossRef]
- Smith, H.L.; Mcausland, L.; Murchie, E.H. Don’t ignore the green light: Exploring diverse roles in plant processes. J. Exp. Bot. 2017, 68, 2099–2110. [Google Scholar] [CrossRef]
- Huché-Thélier, L.; Crespel, L.; Le Gourrierec, J.; Morel, P.; Sakr, S.; Leduc, N. Light signaling and plant responses to blue and UV radiations—Perspectives for applications in horticulture. Environ. Exp. Bot. 2016, 121, 22–38. [Google Scholar] [CrossRef]
- Brazaityte, A.; Duchovskis, P.; Urbonavicuite, A.; Samuoliene, G.; Jankauskiene, J.; Kasiuleviciute-Bonakere, A.; Bliznikas, Z.; Novickovas, A.; Breive, K.; Zukauskas, A. The effect of light-emitting diodes lighting on cucumber transplants and after-effect on yield. Zemdirbyste-Agriculture 2009, 96, 102–118. [Google Scholar]
- Hernandez, R.; Kubota, C. Physiological responses of cucumber seedlings under different blue and red photon flux ratios using LEDs. Environ. Exp. Bot. 2016, 121, 66–74. [Google Scholar] [CrossRef]
- Sager, J.C.; Smith, W.O.; Edwards, J.L.; Cyr, K.L. Photosynthetic efficiency and phytochrome photoequilibria determination using spectral data. Trans. ASAE 1988, 31, 1882–1889. [Google Scholar] [CrossRef]
- Bantis, F.; Koukounaras, A.; Siomos, A.S.; Fotelli, M.N.; Kintzonidis, D. Bichromatic red and blue LEDs during healing enhance the vegetative growth and quality of grafted watermelon seedlings. Sci. Hortic. 2019, 109000. [Google Scholar] [CrossRef]
- McGuire, R.G. Reporting of objective color measurements. HortScience 1992, 27, 1254–1255. [Google Scholar] [CrossRef] [Green Version]
- Dickson, A.; Leaf, A.L.; Hosner, J.F. Quality appraisal of white spruce and white pine seedling stock in nurseries. Forest. Chron. 1960, 36, 10–13. [Google Scholar] [CrossRef]
- Barrett, D.M.; Beaulieu, J.C.; Shewfelt, R. Color, flavor, texture, and nutritional quality of fresh-cut fruits and vegetables: Desirable levels, instrumental and sensory measurement, and the effects of processing. Crit. Rev. Food Sci. Nutr. 2010, 50, 369–389. [Google Scholar] [CrossRef]
- Fan, X.; Zhang, J.; Xu, Z.; Guo, s.; Jiao, X.; Liu, X.; Gao, Y. Effects of different light quality on growth, chlorophyll concentration and chlorophyll biosynthesis precursors of non-heading Chinese cabbage (Brassica campestris L.). Acta Physiol. Plant. 2013, 35, 2721–2726. [Google Scholar] [CrossRef]
- Snowden, M.C.; Cope, K.R.; Bugbee, B. Sensitivity of seven diverse species to blue and green light: Interactions with photon flux. PLoS ONE 2016, 11. [Google Scholar] [CrossRef]
- Ahmad, M.; Cashmore, A.R. The blue-light receptor cryptochrome 1 shows functional dependence on phytochrome A of phytochrome B in Arabidopsis thaliana. Plant J. 1997, 11, 421–427. [Google Scholar] [CrossRef]
- Nanzin, M.T.; Lefsrud, M.; Gravel, V.; Azad, M.O.K. Blue light added with red LEDs enhance growth characteristics, pigments content, and antioxidant capacity in lettuce, spinach, kale, basil, and sweet pepper in a controlled environment. Plants 2019, 8, 93. [Google Scholar]
- Hitz, T.; Hartung, J.; Graeff-Honninger, S.; Munz, S. Morphological response of soybean (Glycine max (L.) Merr.) cultivars to light intensity and red to far-red ratio. Agronomy 2019, 9, 428. [Google Scholar] [CrossRef] [Green Version]
- Hogewoning, S.W.; Peter, D.; Trouwborst, G.; van leperen, W.; Harbinson, J. An artificial solar spectrum substantially alters plant developmenr compared with usual climate room irradiance spectra. J. Exp. Bot. 2010, 61, 1267–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadalini, S.; Zucchi, P.; Andreotti, C. Effects of blue and red LED lights on soilless cultivated strawberry growth performances and fruit quality. Eur. J. Hortic. Sci. 2017, 82, 12–20. [Google Scholar] [CrossRef]
- Li, Y.; Xin, G.; Wei, M.; Shi, Q.; Yang, F.; Wang, X. Carbohydrate accumulation and sucrose metabolism responses in tomato seedling leaves when subjected to different light qualities. Sci. Hortic. 2017, 225, 490–497. [Google Scholar] [CrossRef]
- McCree, K.J. The action spectrum, absorptance and quantum yield of photosynthesis in crop plants. Agric. Metereol. 1972, 9, 191–216. [Google Scholar] [CrossRef]
- Meng, Q.; Runkle, E.S. Far-red radiation interacts with relative and absolute blue and red photon flux densities to regulate growth, morphology, and pigmentation of lettuce and basil seedlings. Sci. Hortic. 2019, 255, 269–280. [Google Scholar] [CrossRef]

 : Lowest not acceptable;
: Lowest not acceptable;  : lowest acceptable;
: lowest acceptable;  : optimum;
: optimum;  : highest acceptable. Each percentage was computed from three trays each having n = 108 or 80 seedlings for watermelon and interspecific squash, respectively.
: Lowest not acceptable; : lowest acceptable; : optimum; : highest acceptable. Each percentage was computed from three trays each having n = 108 or 80 seedlings for watermelon and interspecific squash, respectively.
: highest acceptable. Each percentage was computed from three trays each having n = 108 or 80 seedlings for watermelon and interspecific squash, respectively.
: Lowest not acceptable; : lowest acceptable; : optimum; : highest acceptable. Each percentage was computed from three trays each having n = 108 or 80 seedlings for watermelon and interspecific squash, respectively.
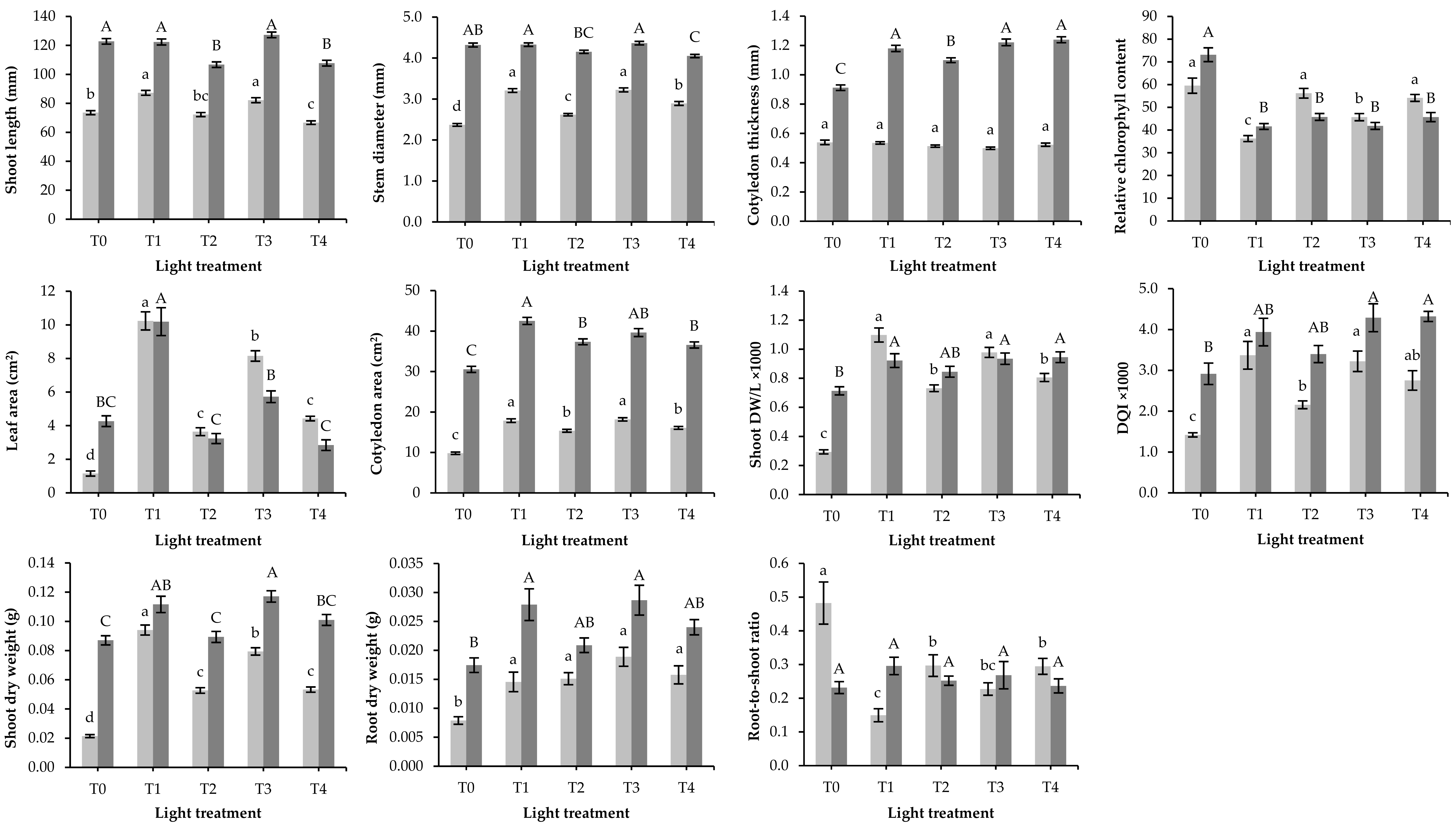
 ) and interspecific squash (
) and interspecific squash (  ) seedlings after 13 or 12 days, respectively, in a growth chamber under five different light treatments described in Table 1. DW/L: shoot dry weight-to-length ratio; DQI: Dickson’s quality index. Bars followed by different letters are significantly different (p ≤ 0.05). Each mean value was computed from n = 36 seedlings except for root dry weight, root-to-shoot ratio, and DQI which were computed from n = 18 seedlings.
) and interspecific squash ( ) seedlings after 13 or 12 days, respectively, in a growth chamber under five different light treatments described in Table 1. DW/L: shoot dry weight-to-length ratio; DQI: Dickson’s quality index. Bars followed by different letters are significantly different (p ≤ 0.05). Each mean value was computed from n = 36 seedlings except for root dry weight, root-to-shoot ratio, and DQI which were computed from n = 18 seedlings.
) seedlings after 13 or 12 days, respectively, in a growth chamber under five different light treatments described in Table 1. DW/L: shoot dry weight-to-length ratio; DQI: Dickson’s quality index. Bars followed by different letters are significantly different (p ≤ 0.05). Each mean value was computed from n = 36 seedlings except for root dry weight, root-to-shoot ratio, and DQI which were computed from n = 18 seedlings.
) and interspecific squash ( ) seedlings after 13 or 12 days, respectively, in a growth chamber under five different light treatments described in Table 1. DW/L: shoot dry weight-to-length ratio; DQI: Dickson’s quality index. Bars followed by different letters are significantly different (p ≤ 0.05). Each mean value was computed from n = 36 seedlings except for root dry weight, root-to-shoot ratio, and DQI which were computed from n = 18 seedlings.

{kind=link}
{kind=link}
{kind=link}
| Parameters | Light Treatment | ||||
|---|---|---|---|---|---|
| T0 | T1 | T2 | T3 | T4 | |
| UV %; 380–399 nm | 0.11 | 0.02 | 0.02 | 0.02 | 0.36 |
| Blue %; 400–499 nm | 20.04 | 7.62 | 10.90 | 11.38 | 20.59 |
| Green %; 500–599 nm | 40.97 | 2.34 | 18.54 | 13.85 | 36.46 |
| Red %; 600–699 nm | 34.75 | 67.25 | 62.20 | 56.48 | 36.92 |
| Far-red %; 700–780 nm | 4.12 | 22.77 | 8.34 | 18.28 | 5.68 |
| Blue peak wavelength (nm) | 436 | 448 | 448 | 448 | 461 |
| Red peak wavelength (nm) | 612 | 660 | 631 | 660 | 660 |
| R:B | 0.63 | 10.12 | 5.58 | 6.39 | 1.93 |
| R:FR | 1.98 | 2.69 | 5.30 | 2.68 | 5.80 |
| PSS | 0.89 | 0.75 | 0.88 | 0.73 | 0.87 |
| CCT (K) | 3830 | 0 | 1624 | 2143 | 5034 |
| CRI | 83.5 | 0 | 66.1 | 71.0 | 87.7 |
| Species | Parameters | Light Treatment | ||||
|---|---|---|---|---|---|---|
| T0 | T1 | T2 | T3 | T4 | ||
| Watermelon | L* | 43.33 ± 0.41 a | 42.60 ± 0.26 ab | 40.94 ± 0.33 c | 42.09 ± 0.57 bc | 42.69 ± 0.98 bc |
| C* | 21.15 ± 0.33 c | 25.31 ± 0.51 a | 20.45 ± 0.44 c | 22.69 ± 0.54 b | 19.82 ± 0.75 c | |
| h° | 127.0 ± 0.27 b | 126.6 ± 0.21 b | 128.6 ± 0.22 a | 127.7 ± 0.16 b | 128.8 ± 0.28 a | |
| a*/b* | −0.76 ± 0.01 a | −0.74 ± 0.01 a | −0.80 ± 0.01 b | −0.77 ± 0.00 a | −0.81 ± 0.01 b | |
| Interspecific Squash | L* | 38.27 ± 0.45 d | 42.05 ± 0.32 ab | 40.15 ± 0.28 c | 42.48 ± 0.40 a | 40.96 ± 0.30 bc |
| C* | 20.81 ± 0.41 c | 26.86 ± 0.35 a | 24.34 ± 0.40 b | 27.61 ± 0.41 a | 24.68 ± 0.32 b | |
| h° | 129.5 ± 0.22 a | 127.0 ± 0.15 cd | 128.0 ± 0.17 b | 126.7 ± 0.19 d | 127.6 ± 0.12 bc | |
| a*/b* | −0.83 ± 0.01 d | −0.75 ± 0.00 ab | −0.78 ± 0.00 c | −0.75 ± 0.01 a | −0.77 ± 0.00 bc | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bantis, F.; Koukounaras, A.; Siomos, A.S.; Radoglou, K.; Dangitsis, C. Optimal LED Wavelength Composition for the Production of High-Quality Watermelon and Interspecific Squash Seedlings Used for Grafting. Agronomy 2019, 9, 870. https://doi.org/10.3390/agronomy9120870
Bantis F, Koukounaras A, Siomos AS, Radoglou K, Dangitsis C. Optimal LED Wavelength Composition for the Production of High-Quality Watermelon and Interspecific Squash Seedlings Used for Grafting. Agronomy. 2019; 9(12):870. https://doi.org/10.3390/agronomy9120870
Chicago/Turabian StyleBantis, Filippos, Athanasios Koukounaras, Anastasios S. Siomos, Kalliopi Radoglou, and Christodoulos Dangitsis. 2019. "Optimal LED Wavelength Composition for the Production of High-Quality Watermelon and Interspecific Squash Seedlings Used for Grafting" Agronomy 9, no. 12: 870. https://doi.org/10.3390/agronomy9120870
APA StyleBantis, F., Koukounaras, A., Siomos, A. S., Radoglou, K., & Dangitsis, C. (2019). Optimal LED Wavelength Composition for the Production of High-Quality Watermelon and Interspecific Squash Seedlings Used for Grafting. Agronomy, 9(12), 870. https://doi.org/10.3390/agronomy9120870