Didymella pinodes Affects N and P Uptakes and Their Efficiencies in a Tripartite Mutualism of Pea




Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Experimental Soil, Symbiont and Nutrient Management
2.3. Biological Materials
2.4. Plant Harvest and Nutrient Measurements
2.5. Statistical Analyses
3. Results
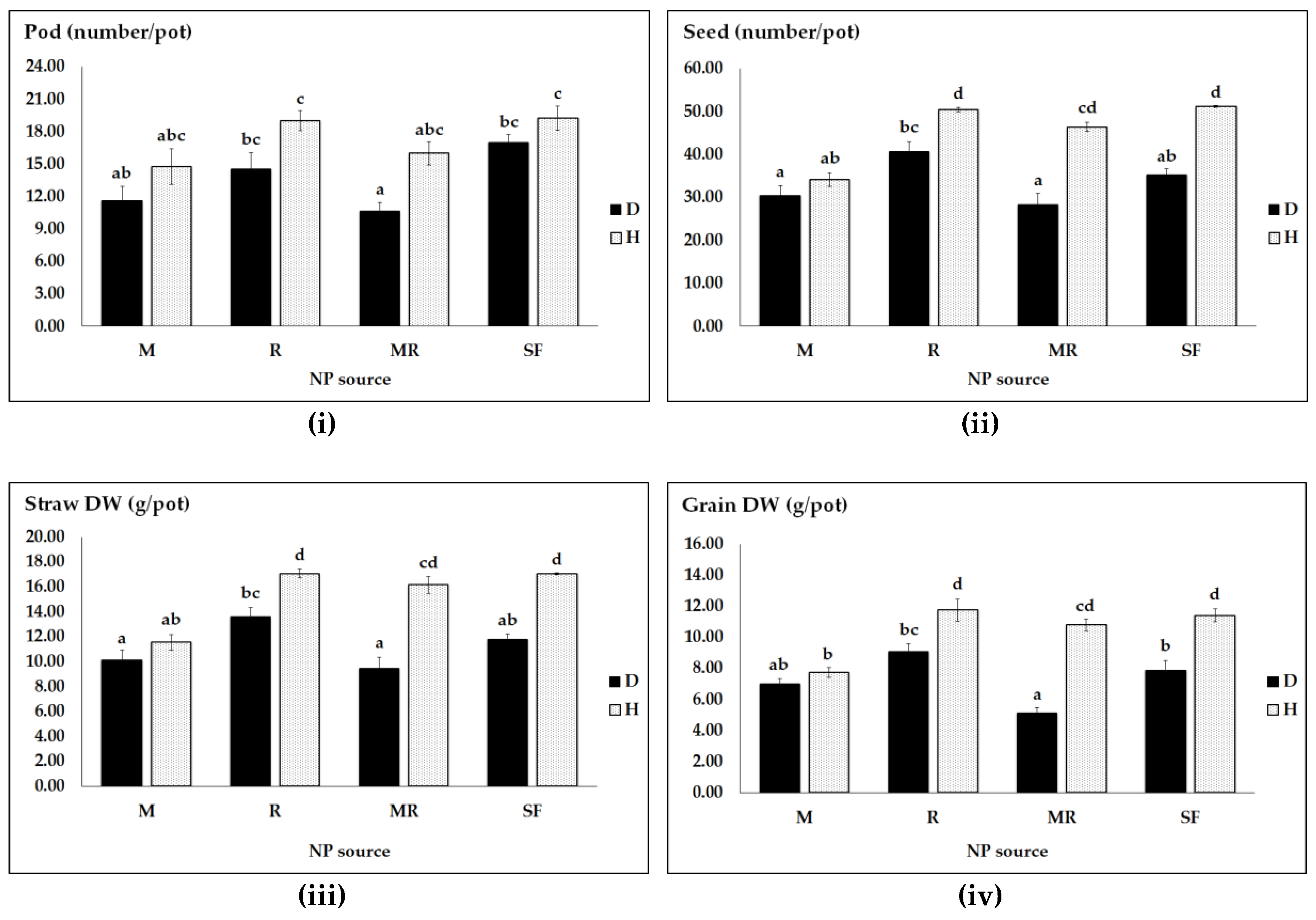
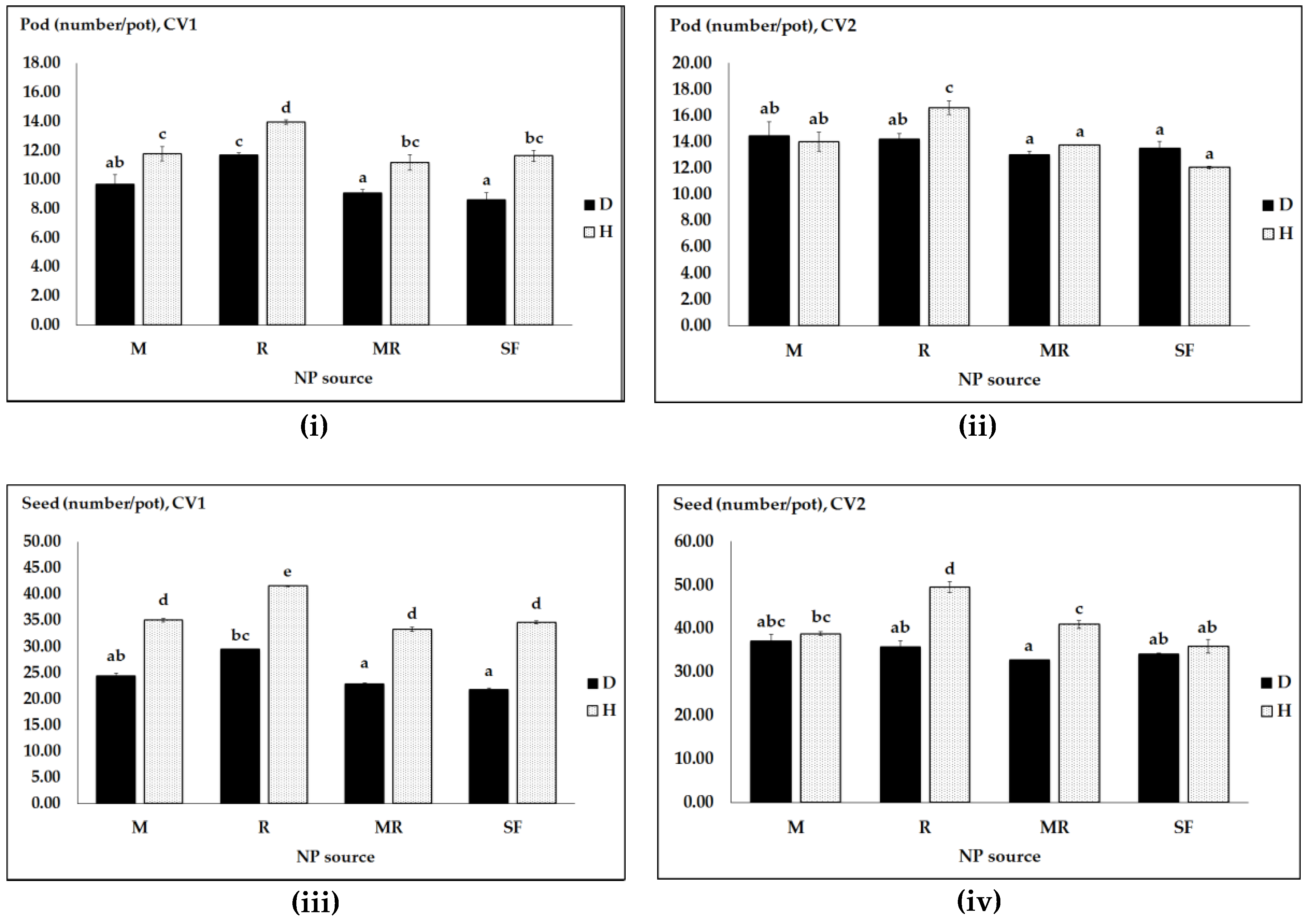
3.1. Effects of N and P Regime, Pathogen Infection and Genotype on Pea Productivity
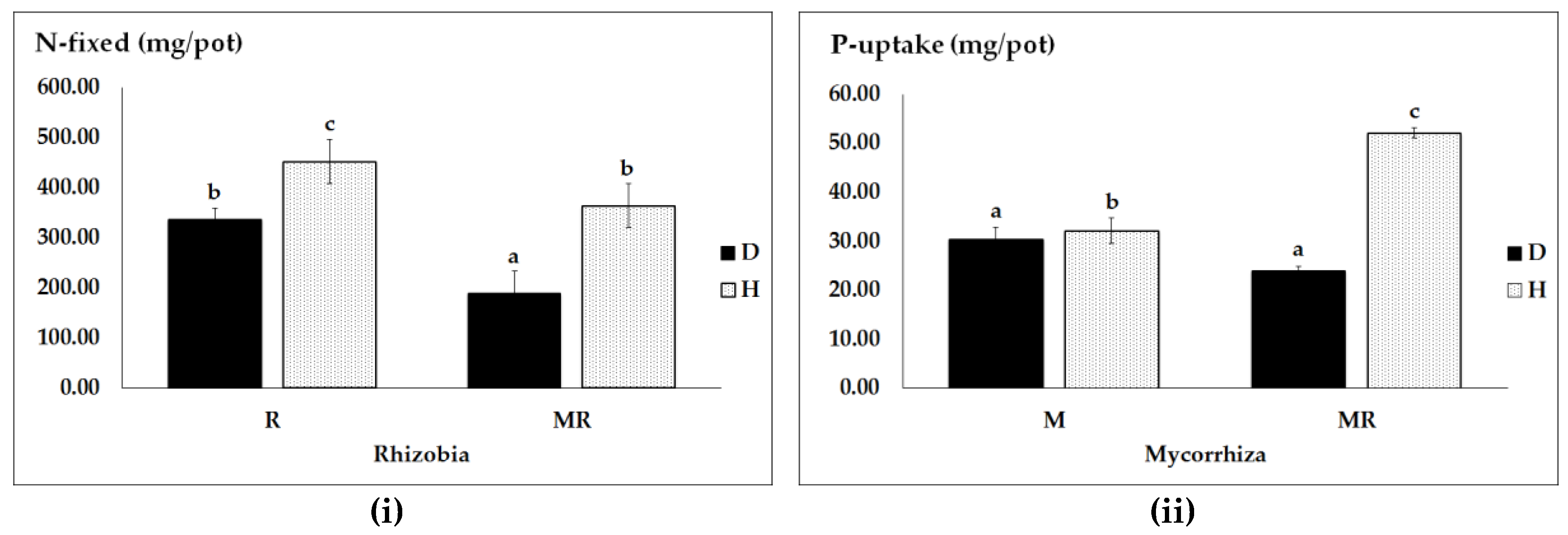
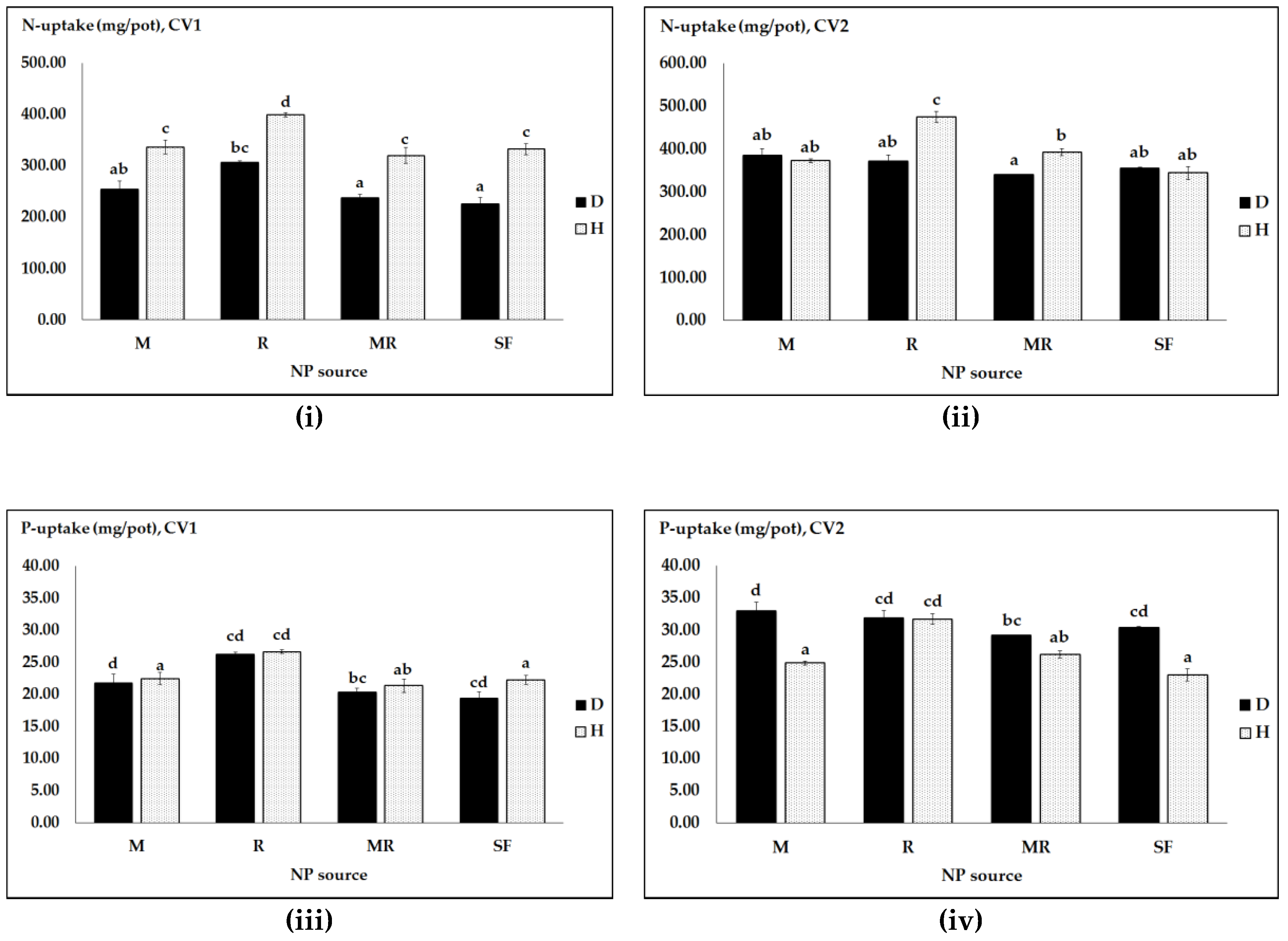
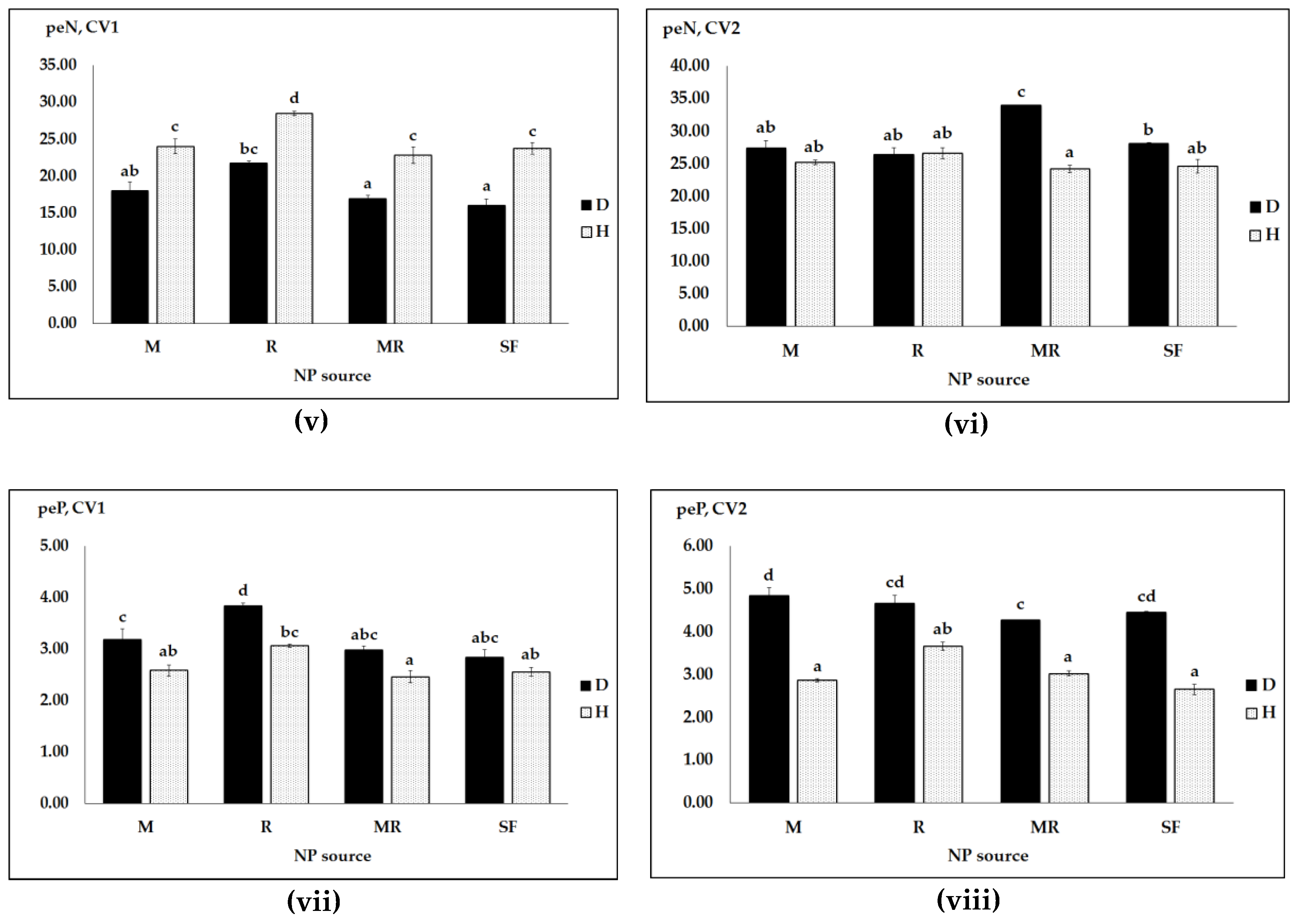
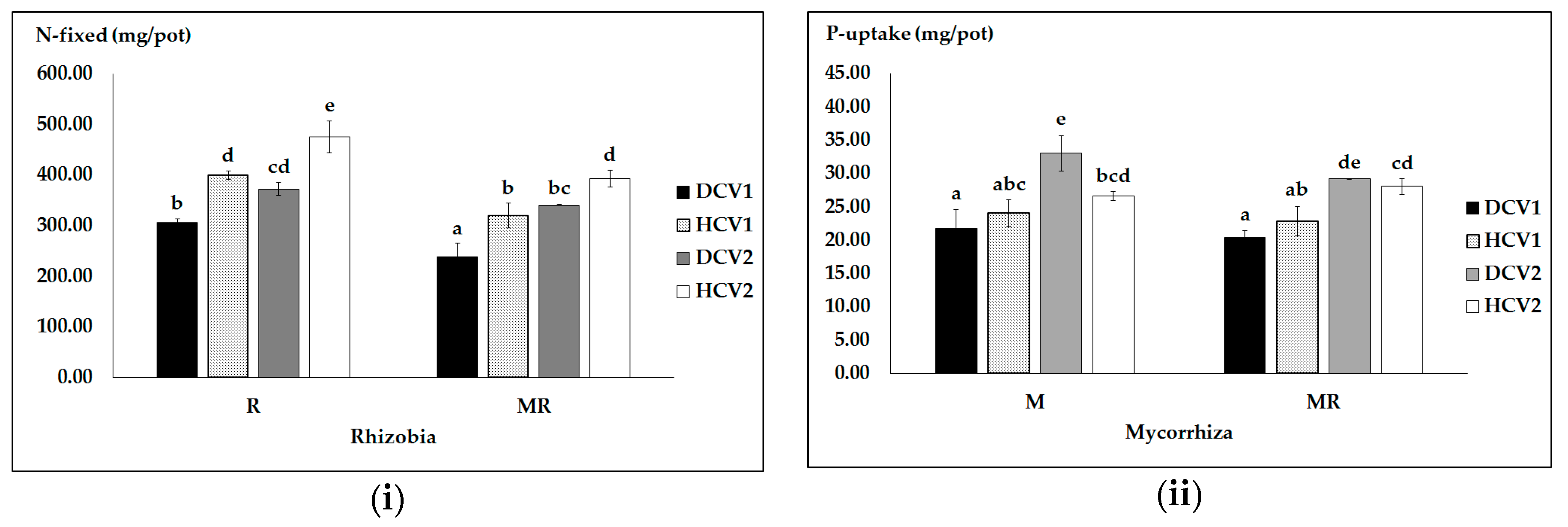
3.2. Effects of N and P Sources, Pathogen Infection and Genotype on N and P Uptake and Efficiency
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Xue, A.G.; Warkentin, T.D.; Kenaschuk, E.O. Effects of timings of inoculation with Mycosphaerella pinodes on yield and seed infection of field pea. Can. J. Plant Sci. 1997, 77, 685–689. [Google Scholar] [CrossRef]
- Tivoli, B.; Banniza, S. Comparison of the epidemiology of ascochyta blights on grain legumes. Eur. J. Plant Pathol. 2007, 119, 59–76. [Google Scholar] [CrossRef]
- Crozat, Y.; Aveline, A.; Coste, F.; Gillet, J.P.; Domenach, A.M. Yield performance and seed production pattern of field-grown pea and soybean in relation to N nutrition. Eur. J. Agron. 1994, 3, 135–144. [Google Scholar] [CrossRef]
- Jensen, E.S. Seasonal patterns of growth and nitrogen fixation in field-grown pea. Plant Soil. 1987, 101, 29–37. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D. Growth and carbon economy of arbuscular mycorrhizal symbionts. In Mycorrhizal Symbiosis; Elsevier: Amsterdam, The Netherlands, 2008; pp. 117–144. [Google Scholar]
- Selosse, M.-A.; Richard, F.; He, X.; Simard, S.W. Mycorrhizal networks: Des liaisons dangereuses? Trends Ecol. Evol. 2006, 21, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Garg, N.; Chandel, S. Arbuscular mycorrhizal networks: Process and functions. A review. Agron. Sustain. Dev. 2010, 30, 581–599. [Google Scholar] [CrossRef]
- Farzaneh, M.; Vierheilig, H.; Lössl, A.; Kaul, H.P. Arbuscular mycorrhiza enhances nutrient uptake in chickpea. Plant Soil Environ. 2011, 57, 465–470. [Google Scholar] [CrossRef] [Green Version]
- Aryal, U.K.; Xu, H.L.; Fujita, M. Rhizobia and AM fungal inoculation improve growth and nutrient uptake of bean plants under organic fertilization. J. Sustain. Agric. 2003, 21, 29–41. [Google Scholar] [CrossRef]
- Harris, D.; Pacovsky, R.S.; Paul, E.A. Carbon economy of soybean-Rhizobium-Glomus associations. New Phytol. 1985, 101, 427–440. [Google Scholar] [CrossRef]
- Kawai, Y.; Yamamoto, Y. Increase in the formation and nitrogen fixation of soybean nodules by vesicular-arbuscular mycorrhiza. Plant Cell Physiol. 1986, 27, 399–405. [Google Scholar]
- Mortimer, P.E.; Perez-Fernandez, M.A.; Valentine, A.J. The role of arbuscular mycorrhizal colonization in the carbon and nutrient economy of the tripartite symbiosis with nodulated Phaseolus vulgaris. Soil Biol. Biochem. 2008, 40, 1019–1027. [Google Scholar] [CrossRef]
- Paul, E.A.; Kucey, R.M.N. Carbon flow in plant microbial associations. Science 1981, 213, 473–474. [Google Scholar] [CrossRef] [PubMed]
- Redecker, D.; von Berswordt-Wallrabe, P.; Beck, D.P.; Werner, D. Influence of inoculation with arbuscular mycorrhizal fungi on stable isotopes of nitrogen in Phaseolus vulgaris. Biol. Fertil. Soils 1997, 24, 344–346. [Google Scholar] [CrossRef]
- Antunes, P.M.; Deaville, D.; Goss, M.J. Effect of two AMF life strategies on the tripartite symbiosis with Bradyrhizobium japonicum and soybean. Mycorrhiza 2006, 16, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Ide Franzini, V.; Azcón, R.; Latanze Mendes, F.; Aroca, R. Interactions between Glomus species and Rhizobium strains affect the nutritional physiology of drought-stressed legume hosts. J. Plant Physiol. 2010, 167, 614–619. [Google Scholar] [CrossRef] [PubMed]
- Scheublin, T.R.; van der Heijden, M.G.A. Arbuscular mycorrhizal fungi colonize nonfixing root nodules of several legume species. New Phytol. 2006, 172, 732–738. [Google Scholar] [CrossRef] [Green Version]
- Angelopoulou, D.J.; Naska, E.J.; Paplomatas, E.J.; Tjamos, S.E. Biological control agents (BCAs) of Verticillium wilt: Influence of application rates and delivery method on plant protection, triggering of host defence mechanisms and rhizosphere populations of BCAs. Plant Pathol. 2014, 63, 1062–1069. [Google Scholar] [CrossRef]
- Choudhary, D.K.; Prakash, A.; Johri, B.N. Induced systemic resistance (ISR) in plants: Mechanism of action. Indian J. Microbiol. 2007, 47, 289–297. [Google Scholar] [CrossRef]
- Choudhary, K.K.; Agrawal, S.B. Ultraviolet-B induced changes in morphological, physiological and biochemical parameters of two cultivars of pea (Pisum sativum L.). Ecotoxicol. Environ. Saf. 2014, 100, 178–187. [Google Scholar] [CrossRef]
- Jacott, C.N.; Murray, J.D.; Ridout, C.J. Trade-offs in arbuscular mycorrhizal symbiosis: Disease resistance, growth responses and perspectives for crop breeding. Agronormy 2017, 7, 75. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Van Pelt, J.A.; Verhagen, B.W.M.; Ton, J.; Van Wees, S.C.M.; Leon-Kloosterziel, K.M.; Van Loon, L.C. Induced systemic resistance by plant growth-promoting rhizobacteria. Symbiosis 2003, 35, 39–54. [Google Scholar]
- Desalegn, G.; Turetschek, R.; Kaul, H.P.; Wienkoop, S. Microbial symbionts affect Pisum sativum proteome and metabolome under Didymella pinodes infection. J. Proteom. 2016, 143, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Turetschek, R.; Desalegn, G.; Epple, T.; Kaul, H.P.; Wienkoop, S. Key metabolic traits of Pisum sativum maintain cell vitality during Didymella pinodes infection: Cultivar resistance and the microsymbionts’ influence. J. Proteom. 2017, 169, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Stacey, G.; Zhang, X.C.; Freire, M.C.M.; Le, M.H.; Oliveira, L.O.D.; Pitkin, J.W.; Segers, G.; Concibido, V.C.; Baley, G.J.; Hartman, G.L.; Upchurch, G.; Pedley, K.F. Genetic diversity and origins of Phakopsora pachyrhizi isolates in the United States. Asian J. Plant Pathol. 2012, 6, 52–65. [Google Scholar] [CrossRef]
- Hoffmann, D.; Vierheilig, H.; Peneder, S.; Schausberger, P. Mycorrhiza modulates aboveground tri-trophic interactions to the fitness benefit of its host plant. Ecol. Entomol. 2011, 36, 574–581. [Google Scholar] [CrossRef]
- Broughton, W.J.; Dilworth, M.J. Handbook for rhizobia: Methods in legume-Rhizobium technology. In The Quarterly Review of Biology; Somasegaran, P., Hoben, H.J., Eds.; University of Chicago Press: Chicago, IL, USA, 1995; pp. 224–225. [Google Scholar]
- Davidson, J.A.; Krysinska-Kaczmarek, M.; Herdina; McKay, A.; Scott, E.S. Comparison of cultural growth and in planta quantification of Didymella pinodes, Phoma koolunga and Phoma medicaginis var. pinodella, causal agents of ascochyta blight on field pea (Pisum sativum). Mycologia 2012, 104, 93–101. [Google Scholar] [PubMed]
- Delogu, G.; Cattivelli, L.; Pecchioni, N.; De Falcis, D.; Maggiore, T.; Stanca, A.M. Uptake and agronomic efficiency of nitrogen in winter barley and winter wheat. Eur. J. Agron. 1998, 9, 11–20. [Google Scholar] [CrossRef]
- Jia, Y.; Gray, V.M. Interrelationships between nitrogen supply and photosynthetic parameters in Vicia faba L. Photosynthetica 2003, 41, 605–610. [Google Scholar] [CrossRef]
- Jia, Y.; Gray, V.M.; Straker, C.J. The influence of Rhizobium and arbuscular mycorrhizal fungi on nitrogen and phosphorus accumulation by Vicia faba. Ann. Bot. 2004, 94, 251–258. [Google Scholar] [CrossRef]
- Gray, V.M. The Role of the C:N:P Stoichiometry in the Carbon Balance Dynamics of the Legume–AMF–Rhizobium Tripartite Symbiotic Association. In Plant Growth and Health Promoting Bacteria; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Kumar, B.S.D.; Berggren, I.; Martensson, A.M. Potential for improving pea production by co-inoculation with fluorescent Pseudomonas and Rhizobium. Plant Soil 2001, 229, 25–34. [Google Scholar] [CrossRef]
- Larrainzar, E.; Wienkoop, S. A proteomic view on the role of legume symbiotic interactions. Front. Plant Sci. 2017, 8, 1267. [Google Scholar] [CrossRef] [PubMed]
- Bourion, V.; Laguerre, G.; Depret, G.; Voisin, A.S.; Salon, C.; Duc, G. Genetic variability in nodulation and root growth affects nitrogen fixation and accumulation in pea. Ann. Bot. 2007, 100, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Baldy, V.; Thiebaut, G.; Fernandez, C.; Sagova-Mareckova, M.; Korboulewsky, N.; Monnier, Y.; Perez, T.; Tremolieres, M. Experimental assessment of the water quality influence on the phosphorus uptake of an invasive aquatic plant: Biological responses throughout its phenological stage. PLoS ONE 2015, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Paul, N.D. Effects of fungal pathogens on nitrogen phosphorus and sulphur relations of individual plants and populations. In Nitrogen, Phosphorus and Sulphur Utilisation by Fungi: Symposium of the British Mycological Society; Boddy, L., Marchant, R., Read, D.J., Eds.; Cambridge University Press: Cambridge, UK, 1989; pp. 155–179. [Google Scholar]
- Walters, D.R.; Ayres, P.G. Phosphate uptake and translocation by roots of powdery mildew infected barley. Physiol. Plant Pathol. 1981, 18, 195–205. [Google Scholar] [CrossRef]
- Roberts, A.M.; Walters, D.R. Shoot: Root interrelationships in leeks infected with the rust, Puccinia allii Rud.: Growth and nutrient relations. New Phytol. 1989, 111, 223–228. [Google Scholar] [CrossRef]
- Diaz, C.; Purdy, S.; Christ, A.; Morot-Gaudry, J.F.; Wingler, A.; Masclaux-Daubresse, C.L. Characterization of markers to determine the extent and variability of leaf senescence in Arabidopsis. A metabolic profiling approach. Plant Physiol. 2005, 138, 898–908. [Google Scholar] [CrossRef] [PubMed]
- Malagoli, P. Dynamics of nitrogen uptake and mobilization in field-grown winter oilseed rape (Brassica napus) from stem extension to harvest: I. Global N flows between vegetative and reproductive tissues in relation to leaf fall and their residual N. Ann. Bot. 2005, 95, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Musse, M.; De Franceschi, L.; Cambert, M.; Sorin, C.; Le Caherec, F.; Burel, A.; Bouchereau, A.; Mariette, F.; Leport, L. Structural changes in senescing oilseed rape leaves at tissue and subcellular levels monitored by nuclear magnetic resonance relaxometry through water status. Plant Physiol. 2013, 163, 392–406. [Google Scholar] [CrossRef] [PubMed]
- Barraclough, P.B.; Lopez-Bellido, R.; Hawkesford, M.J. Genotypic variation in the uptake, partitioning and remobilisation of nitrogen during grain-filling in wheat. Field Crop. Res. 2014, 156, 242–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, J.; Cheng, C.; Yang, J.; Wang, Q. Pathogen infection drives patterns of nutrient resorption in citrus plants. Sci. Rep. 2015, 5, 14675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masclaux, C.; Quillere, I.; Gallais, A.; Hirel, B. The challenge of remobilisation in plant nitrogen economy. A survey of physio-agronomic and molecular approaches. Ann. Appl. Biol. 2001, 138, 69–81. [Google Scholar] [CrossRef]
- Sistani, N.R.; Kaul, H.P.; Desalegn, G.; Wienkoop, S. Rhizobium impacts on seed productivity, quality, and protection of Pisum sativum upon disease stress caused by didymella pinodes: Phenotypic, proteomic, and metabolomic traits. Front. Plant Sci. 2017, 8, 1961. [Google Scholar] [CrossRef] [PubMed]
- Habte, M.; Osorio, N.W. Arbuscular Mycorrhizas: Producing and Applying Arbuscular Mycorrhizal Inoculum. Honolulu (HI); University of Hawaii: Honolulu, HI, USA, 2001; p. 47. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crop Productivity (F-Values) | ||||
|---|---|---|---|---|
| Sources | Pod Number | Seed Number | Straw DW | Grain DW |
| Pathogen (PH) | 9.79 ** | 163.15 *** | 88.35 *** | 163.07 *** |
| NP sources (NPS) | 4.56 * | 43.91 *** | 25.26 *** | 43.90 *** |
| PH × NPS | 0.46 ns | 19.19 *** | 5.25 ** | 19.18 *** |
| Crop Productivity (F-Values) | ||||
|---|---|---|---|---|
| Sources | Pod Number | Seed Number | Straw DW | Grain DW |
| Cultivar (CV) | 208.59 *** | 194.58 *** | 204.11 *** | 198.03 *** |
| NP sources (NPS) | 35.04 *** | 36.62 *** | 35.59 *** | 36.28 *** |
| Pathogen (PH) | 34.69 *** | 258.08 *** | 85.55 *** | 185.37 *** |
| CV X NPS | 0.68 ns | 0.67 ns | 0.67 ns | 0.67 ns |
| CV X PH | 34.77 *** | 22.04 *** | 30.21 *** | 24.81 *** |
| NPS X PH | 5.32 ** | 7.13 *** | 5.86 ** | 6.67 *** |
| CV X PH X NPS | 7.08 *** | 7.01 *** | 7.09 *** | 7.01 *** |
| Sources | Seed N and P Uptake and Efficiency (F-Values) | |||
|---|---|---|---|---|
| N Uptake | peN | P Uptake | peP | |
| PH | 128.98 *** | 562.14 *** | 124.30 *** | 3.3 |
| NPS | 50.22 *** | 7.50 *** | 60.45 *** | 7.92 *** |
| PH X NPS | 7.95 *** | 4.52 ** | 12.11 *** | 8.61 *** |
| Pathogen | NP Sources | Seed NP Uptake and Efficiency | |||
|---|---|---|---|---|---|
| N Uptake (mg/pot) | peN (mg/pot) | P Uptake (mg/pot) | peP (mg/pot) | ||
| Infected | M | 342.66 a | 35.36 b | 38.06 a | 3.11 abc |
| R | 486.23 bc | 47.37 c | 67.99 bc | 3.33 c | |
| MR | 338.33 a | 33.46 b | 36.16 a | 2.48 a | |
| SF | 421.19 ab | 41.04 bc | 62.32 b | 3.25 bc | |
| Healthy | M | 393.79 a | 6.89 a | 42.08 a | 2.50 ab |
| R | 652.08 d | 9.06 a | 96.84 d | 3.54 c | |
| MR | 563.83 c | 8.36 a | 82.05 cd | 3.57 c | |
| SF | 659.01 d | 9.94 a | 97.10 d | 3.50 c | |
| Sources | Seed N and P Uptake and Efficiency (F-Values) | |||
|---|---|---|---|---|
| N Uptake | peN | P Uptake | peP | |
| Cultivar (CV) | 201.62 *** | 201.29 *** | 220.18 *** | 231.98 *** |
| NP Sources (NPS) | 35.92 *** | 35.98 *** | 33.15 *** | 29.88 *** |
| Pathogen (PH) | 126.26 *** | 133.82 *** | 16.22 *** | 326.76 *** |
| CV X NPS | 0.66 ns | 0.66 ns | 0.76 ns | 1.00 ns |
| CV X PH | 27.75 *** | 27.36 *** | 49.27 *** | 72.49 *** |
| NPS X PH | 6.21 *** | 6.29 *** | 3.94 * | 2.70 ns |
| CV X PH X NPS | 7.06 *** | 7.06 *** | 7.05 *** | 6.85 *** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Desalegn, G.; Turetschek, R.; Wienkoop, S.; Kaul, H.-P. Didymella pinodes Affects N and P Uptakes and Their Efficiencies in a Tripartite Mutualism of Pea. Agronomy 2019, 9, 52. https://doi.org/10.3390/agronomy9020052
Desalegn G, Turetschek R, Wienkoop S, Kaul H-P. Didymella pinodes Affects N and P Uptakes and Their Efficiencies in a Tripartite Mutualism of Pea. Agronomy. 2019; 9(2):52. https://doi.org/10.3390/agronomy9020052
Chicago/Turabian StyleDesalegn, Getinet, Reinhard Turetschek, Stefanie Wienkoop, and Hans-Peter Kaul. 2019. "Didymella pinodes Affects N and P Uptakes and Their Efficiencies in a Tripartite Mutualism of Pea" Agronomy 9, no. 2: 52. https://doi.org/10.3390/agronomy9020052
APA StyleDesalegn, G., Turetschek, R., Wienkoop, S., & Kaul, H. -P. (2019). Didymella pinodes Affects N and P Uptakes and Their Efficiencies in a Tripartite Mutualism of Pea. Agronomy, 9(2), 52. https://doi.org/10.3390/agronomy9020052