Modelling Nitrogen Uptake in Plants and Phytoplankton: Advantages of Integrating Flexibility into the Spatial and Temporal Dynamics of Nitrate Absorption



Abstract
:1. Introduction
1.1. Empirical Models Used to Determine the Kinetic Parameters of N Uptake Rate under Field Conditions for Different Levels of N Fertilization
1.2. Effects of N Fertilization on Temporal Structure of N Uptake Process in Brassica Napus Plants under Field Conditions
1.3. Effects of N Fertilization on N Dilution Curves and N Uptake Process in Brassica Napus Plants under Field Conditions
1.4. Origins of the Overestimation of N Uptake Capacities in Most Mechanistic Models
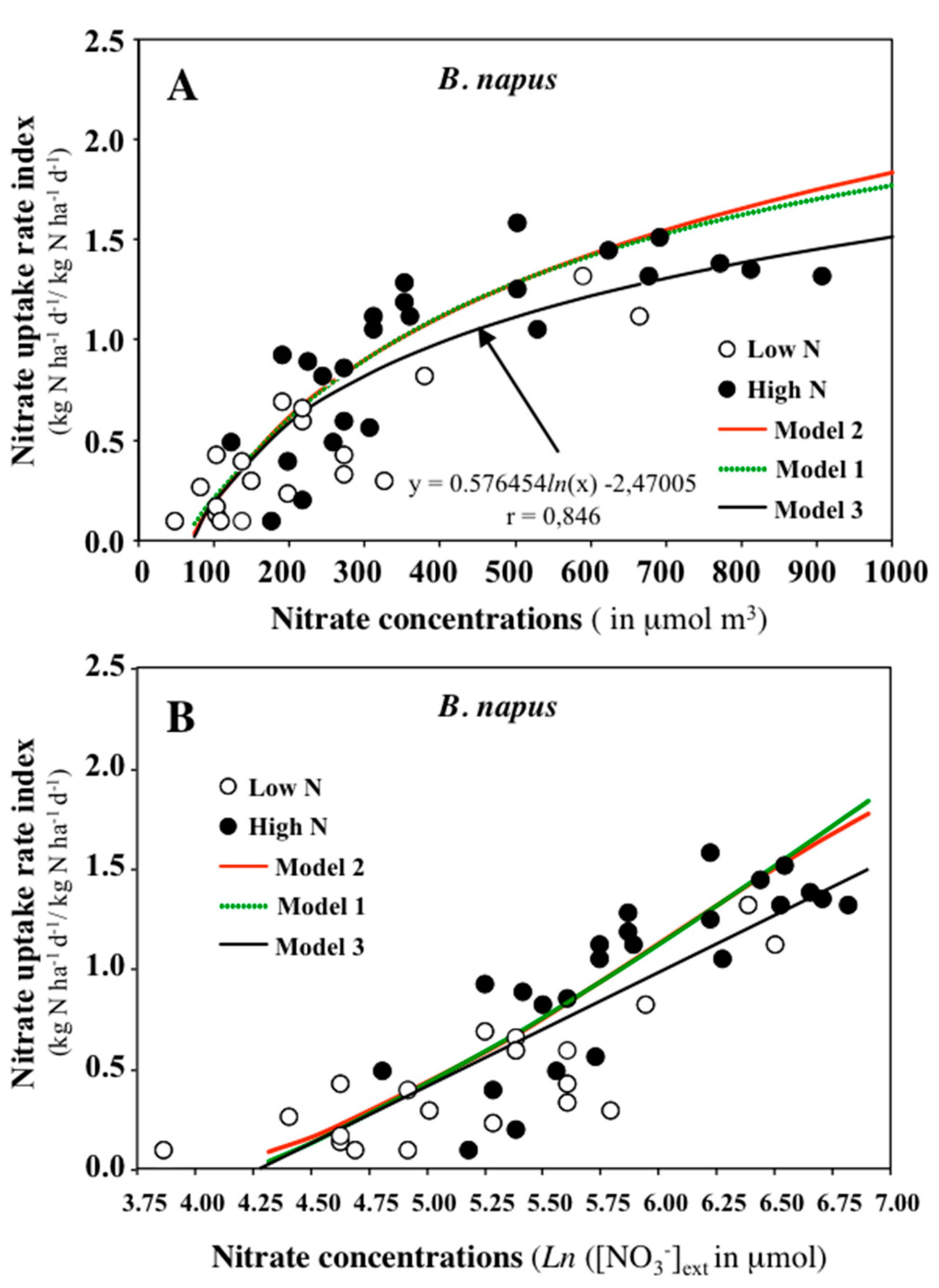
1.5. How Do We Interpret the Kinetic Parameters Deduced from the Mathematical Adjustment of Ion Uptake Isotherms?
1.6. Enzyme–Substrate Interpretation of Ion Uptake Isotherm: Deduction of Microscopic Parameters Vm and Km
Sje + C ⇆ CS → C + Sji
k−1
2. Flow–Force Interpretation of Kinetic Parameters of Nutrient Isotherms: Deduction of Lj and πj Macroscopic Parameters
3. Meaning and Validity of Kinetic Parameters of Nutrient Isotherms at Any Root Scale
4. Introduction of Spatial and Temporal Flexibility in ion Uptake Rate Modelling
4.1. Introduction of a Flexible Uptake Process by Regulating the Number of Uptake Sites at the Cell Membrane Level in Response to Changes in Nutrient Concentrations
4.1.1. Plastic MM Model Formalism
4.1.2. Trait-Based Model Formalism
4.2. Cross-Combination of the Flow–Force Theory with Temporal Variations of Root N Uptake Rate in Response to Environmental Changes
5. Spatial Variation of Root N Uptake Rate with Age along the Root Axes
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Silberbush, M.; Barber, S.A. Sensitivity analysis of parameters used in simulating K uptake with a mechanistic mathematical model 1. Agron. J. 1983, 75, 851–854. [Google Scholar] [CrossRef]
- Barber, S.A. Soil Nutrient Bioavailability: A Mechanistic Approach; John Wiley & Sons: New York, NY, USA, 1995; ISBN 0-471-58747-8. [Google Scholar]
- Tinker, P.B.H.; Nye, P.H. Solute Transport in the Rhizosphere, 2nd ed.; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Roose, T.; Kirk, G.J.D. The solution of convection–diffusion equations for solute transport to plant roots. Plant Soil 2009, 316, 257–264. [Google Scholar] [CrossRef]
- Tournier, P.-H.; Hecht, F.; Comte, M. Finite element model of soil water and nutrient transport with root uptake: Explicit geometry and unstructured adaptive meshing. Transp. Porous. Med. 2015, 106, 487–504. [Google Scholar] [CrossRef]
- York, L.M.; Nord, E.; Lynch, J. Integration of root phenes for soil resource acquisition. Front. Plant Sci. 2013, 4, 355. [Google Scholar] [CrossRef] [PubMed]
- York, L.M.; Silberbush, M.; Lynch, J.P. Spatiotemporal variation of nitrate uptake kinetics within the maize (Zea mays L.) root system is associated with greater nitrate uptake and interactions with architectural phenes. J. Exp. Bot. 2016, 67, 3763–3775. [Google Scholar] [CrossRef] [PubMed]
- Dunbabin, V.; Rengel, Z.; Diggle, A.J. Simulating form and function of root systems: efficiency of nitrate uptake is dependent on root system architecture and the spatial and temporal variability of nitrate supply. Funct. Ecol. 2004, 18, 204–211. [Google Scholar] [CrossRef] [Green Version]
- Le Deunff, E.; Malagoli, P. An updated model for nitrate uptake modelling in plants. I. Functional component: Cross-combination of flow–force interpretation of nitrate uptake isotherms, and environmental and in planta regulation of nitrate influx. Ann. Bot. 2014, 113, 991–1005. [Google Scholar] [CrossRef] [PubMed]
- Malagoli, P.; Le Deunff, E. An updated model for nitrate uptake modelling in plants. II. Assessment of active root involvement in nitrate uptake based on integrated root system age: Measured versus modelled outputs. Ann. Bot. 2014, 113, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- Le Deunff, E.; Tournier, P.-H.; Malagoli, P. The thermodynamic flow-force interpretation of root nutrient uptake kinetics: A powerful formalism for agronomic and phytoplanktonic models. Front. Physiol. 2016, 7, 243. [Google Scholar] [CrossRef] [PubMed]
- Buis, R. Vers les modèles stucturés de croissance. In Biomathématique de la croissance, le cas des végétaux; Grenoble Science: Grenoble, France, 2016; pp. 409–462. ISBN 978-2-7598-1778-8. [Google Scholar]
- Sepaskhah, A.R.; Fahandezh-Saadi, S.; Zand-Parsa, S. Logistic model application for prediction of maize yield under water and nitrogen management. Agr. Water Manage. 2011, 99, 51–57. [Google Scholar] [CrossRef]
- Châlons Oilseed Rape Data Bas. Available online: https://ecosys.versaillesgrignon.inra.fr/ceres_mais/base/welcome.html (accessed on 21 February 2019).
- Gosse, G.; Cellier, P.; Denoroy, P.; Gabrielle, B.; Laville, P.; Leviel, B.; Justes, E.; Nicolardot, B.; Mary, B.; Recous, S.; et al. Water, carbon and nitrogen cycling in a rendzina soil cropped with winter oilseed rape: The Chalons Oilseed Rape Database. Agronomie 1999, 19, 119–124. [Google Scholar] [CrossRef]
- Buis, R. Growth activity and structure at various organization levels in plants. Acta Biotheor 1993, 41, 231. [Google Scholar] [CrossRef]
- Netzer, M.H.; Tittonnel, E.D.; Merrien, A. Evénements liés au passage du méristème de l’état végétatif à l’état reproducteur, chez le colza, Brassica napus L. var. oleifera Metzg. Agronomie 1989, 9, 151–159. [Google Scholar] [CrossRef]
- Malagoli, P.; Meuriot, F.; Laine, P.; Le Deunff, E.; Ourry, A. Modelling nitrate uptake and N dynamics in winter oilseed rape (Brassica napus L.) during the growth cycle. In Quantifying and Understanding Plant Nitrogen Uptake for Systems Modeling; Taylor and Francis: Boca Raton, FL, USA, 2008; pp. 47–69. [Google Scholar]
- Le Deunff, E.; Malagoli, P. Breaking conceptual locks in modelling root absorption of nutrients: Reopening the thermodynamic viewpoint of ion transport across the root. Ann. Bot. 2014, 114, 1555–1570. [Google Scholar] [CrossRef] [PubMed]
- Justes, E.; Mary, B.; Meynard, J.-M.; Machet, J.-M.; Thelier-Huché, L. Determination of a critical nitrogen dilution curve for winter wheat crops. Ann. Bot. 1994, 74, 397–407. [Google Scholar] [CrossRef]
- Colnenne, C.; Meynard, J.M.; Reau, R.; Justes, E.; Merrien, A. Determination of a critical nitrogen dilution curve for winter oilseed rape. Ann. Bot. 1998, 81, 311–317. [Google Scholar] [CrossRef]
- Devienne-Barret, F.; Justes, E.; Machet, J.M.; Mary, B. Integrated control of nitrate uptake by crop growth rate and soil nitrate availability under field conditions. Ann Bot 2000, 86, 995–1005. [Google Scholar] [CrossRef]
- Sadras, V.O.; Lemaire, G. Quantifying crop nitrogen status for comparisons of agronomic practices and genotypes. Field Crop. Res. 2014, 164, 54–64. [Google Scholar] [CrossRef]
- Greenwood, D.J.; Lemaire, G.; Gosse, G.; Cruz, P.; Draycott, A.; Neeteson, J.J. Decline in percentage N of C3 and C4 crops with increasing plant mass. Ann Bot 1990, 66, 425–436. [Google Scholar] [CrossRef]
- Lemaire, G.; Salette, J.; Sigogne, M.; Terrasson, J.-P. Relation entre dynamique de croissance et dynamiquede prélèvement d’azote pour un peuplement degraminées fourragères. I.—Etude de l’effet du milieu. Agronomie 1984, 4, 423–430. [Google Scholar] [CrossRef]
- Malagoli, P.; Laine, P.; Rossato, L.; Ourry, A. Dynamics of nitrogen uptake and mobilization in field-grown winter oilseed rape (Brassica napus L) from stem extension to harvest—I. Global N flows between vegetative and reproductive tissues in relation to leaf fall and their residual N. Ann. Bot. 2005, 95, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Barraclough, P.B. The growth and activity of winter wheat roots in the field: Nutrient inflows of high-yielding crops. J. Agricul. Sci. 1986, 106, 53–59. [Google Scholar] [CrossRef]
- Recous, S.; Machet, J.M.; Mary, B. The fate of labelled15N urea and ammonium nitrate applied to a winter wheat crop: II. plant uptake and N efficiency. Plant Soil 1988, 112, 215–224. [Google Scholar] [CrossRef]
- Thellier, M.; Ripoll, C.; Norris, V.; Nikolic, M.; Römheld, V. Solute Uptake in Plants: A Flow/Force Interpretation. In Progress in Botany; Lüttge, U., Beyschlag, W., Büdel, B., Francis, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 53–68. ISBN 978-3-540-68421-3. [Google Scholar]
- Epstein, E. Dual pattern of ion Absorption by plant cells and by plants. Nature 1966, 212, 1324. [Google Scholar] [CrossRef]
- Shaw, A.; Takacs, I.; Pagilla, K.; Riffat, R.; DeClippeleir, H.; Wilson, C.; Murthy, S. Toward universal half-saturation coefficients: Describing extant KS as a function of diffusion. Water Environ. Res. 2015, 87, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Gloser, V.; Zwieniecki, M.A.; Orians, C.M.; Holbrook, N.M. Dynamic changes in root hydraulic properties in response to nitrate availability. J. Exp. Bot. 2007, 58, 2409–2415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakurai-Ishikawa, J.; Murai-Hatano, M.; Hayashi, H.; Ahamed, A.; Fukushi, K.; Matsumoto, T.; Kitagawa, Y. Transpiration from shoots triggers diurnal changes in root aquaporin expression. Plant Cell Environ. 2011, 34, 1150–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Ny, F.; Leblanc, A.; Beauclair, P.; Deleu, C.; Le Deunff, E. In low transpiring conditions, nitrate and water fluxes for growth of B. napus plantlets correlate with changes in BnNrt2.1 and BnNrt1.1 transporter expression. Plant Signal. Behav. 2013, 8, e22902. [Google Scholar] [CrossRef] [PubMed]
- Nye, P.H.; Tinker, P.B. Solute movement in the soil-root system. Stud. Ecol. 1977, 4, 342. [Google Scholar] [CrossRef]
- Robinson, D.; Linehan, D.J.; Caul, S. What limits nitrate uptake from soil? Plant Cell Environ. 1991, 14, 77–85. [Google Scholar] [CrossRef]
- Eschel, A.; Waisel, Y. Heterogeneity of ion uptake mechanisms along primary roots of corn seedlings. In Ions Transport in Plants; Anderson WP: London, UK, 1973; pp. 531–538. [Google Scholar]
- Lazof, D.B.; Rufty, T.W.; Redinbaugh, M.G. Localization of nitrate absorption and translocation within morphological regions of the corn root. Plant Physiol. 1992, 100, 1251–1258. [Google Scholar] [CrossRef] [PubMed]
- Colmer, T.D.; Bloom, A.J. A comparison of NH4+ and NO3− net fluxes along roots of rice and maize. Plant Cell Environ. 1998, 21, 240–246. [Google Scholar] [CrossRef]
- Sorgona, A.; Lupini, A.; Mercati, F.; Di Dio, L.; Sunseri, F.; Abenavoli, M.R. Nitrate uptake along the maize primary root: An integrated physiological and molecular approach. Plant Cell Environ. 2011, 34, 1127–1140. [Google Scholar] [CrossRef] [PubMed]
- Epstein, E. Mechanism of ion absorption by roots. Nature 1953, 171, 83. [Google Scholar] [CrossRef] [PubMed]
- MacIsaac, J.J.; Dugdale, R.C. The kinetics of nitrate and ammonia uptake by natural populations of marine phytoplankton. Deep Sea Res. Ocean. Abstr. 1969, 16, 45–57. [Google Scholar] [CrossRef]
- Dugdale, R.C. Nutrient Limitation in the Sea: Dynamics, Identification, and Significance1. Limnol. Oceanog. 1967, 12, 685–695. [Google Scholar] [CrossRef]
- Thellier, M. Interprétation électrocinétique de l’absorption minérale chez les végétaux. Bulletin de la Société Française de Physiologie Végétale 1969, 15, 127–139. [Google Scholar]
- Thellier, M. An Electrokinetic Interpretation of the functioning of biological systems and its application to the study of mineral salts absorption. Ann. Bot. 1970, 34, 983–1009. [Google Scholar] [CrossRef]
- Thellier, M. Non-equilibrium thermodynamics and electrokinetic interpretation of biological systems. J. Theor. Bio. 1971, 31, 389–393. [Google Scholar] [CrossRef]
- Thellier, M.; Steltz, T.; Ayadi, A. Application des relations et de la loi d’Onsager à l’interprétation d’interactions entre processus cellulaires globaux. Comptes-rendus de l’Académie des Sciences 1971, 273, 2346–2349. [Google Scholar]
- Thellier, M. Electrokinetic formulation of ionic absorption by plant samples. In Ion Transport in Plants; Elsevier: Amsterdam, The Netherlands, 1973; pp. 47–63. [Google Scholar]
- Teissier, G. Les lois quantitatives de la croissance. Annales de Physiologie et de Physicochimie Biologique 1936, 12, 527–573. [Google Scholar]
- Monod, J. Recherches sur la croissance des cultures bactériennes. Ph.D. Thesis, és Sciences Naturelles, Paris, France, 1941. [Google Scholar]
- Monod, J. The growth of bacterial cultures. Annu. Rev. Microbiol. 1949, 3, 371–394. [Google Scholar] [CrossRef]
- Epstein, E.; Hagen, C.E. A kinetic study of the absorption of alkali cations by barley roots. Plant Physiol. 1952, 27, 457–474. [Google Scholar] [CrossRef] [PubMed]
- Epstein, E.; Rains, D.W.; Elzam, O.E. Resolution of dual mechanisms of potassium absorption by barley roots. Proc. Natl. Acad. Sci. USA 1963, 49, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Le Bot, J.; Adamowicz, S.; Robin, P. Modelling plant nutrition of horticultural crops: A review. Sci. Hortic. 1998, 74, 47–82. [Google Scholar] [CrossRef]
- Ma, L.; Ahuja, L.R.; Bruulsema, T.W. Quantifying and Understanding Plant Nitrogen Uptake for Systems Modeling; Taylor & Francis Group: Boca Raton, FL, USA, 2008. [Google Scholar]
- Smith, S.L.; Yamanaka, Y.; Pahlow, M.; Oschlies, A. Optimal uptake kinetics: Physiological acclimation explains the pattern of nitrate uptake by phytoplankton in the ocean. Mar. Ecol. Prog. Ser. 2009, 384, 1–12. [Google Scholar] [CrossRef]
- Smith, S.L.; Merico, A.; Hohn, S.; Brandt, G. Sizing-up nutrient uptake kinetics: Combining a physiological trade-off with size-scaling of phytoplankton traits. Mar. Ecol. Prog. Ser. 2014, 511, 33–39. [Google Scholar] [CrossRef]
- Aksnes, D.L.; Cao, F.J. Inherent and apparent traits in microbial nutrient uptake. Mar. Ecol. Prog. Ser. 2011, 440, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Fiksen, Ø.; Follows, M.J.; Aksnes, D.L. Trait-based models of nutrient uptake in microbes extend the Michaelis-Menten framework. Limnol. Oceanogr. 2013, 58, 193–202. [Google Scholar] [CrossRef]
- Morel, F.M.M. Kinetics of Nutrient Uptake and Growth in Phytoplankton1. J. Phycol. 1987, 23, 137–150. [Google Scholar] [CrossRef]
- Healey, F.P. Slope of the Monod equation as an indicator of advantage in nutrient competition. Microb. Ecol. 1980, 5, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Aksnes, D.L.; Egge, J.K. A theoretical model for nutrient uptake in phytoplankton. Mar. Ecol. Prog. Ser. 1991, 70, 65–72. [Google Scholar] [CrossRef]
- Lobry, J.R. Re-évaluation du modèle de croissance de Monod. Effet des antibiotiques sur l’énergie de maintenance. Ph.D. Thesis, Université Claude Bernard—Lyon I, Lyon, France, 1991. [Google Scholar]
- Lobry, J.R.; Flandrois, J.P.; Carret, G.; Pave, A. Monod’s bacterial growth model revisited. Bull Math. Biol. 1992, 54, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Bloom, A.J. Wild and cultivated barleys show similar affinities for mineral nitrogen. Oecologia 1985, 65, 555–557. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.B.; Drew, M.C. Nitrogen-13 Studies of Nitrate Fluxes in Barley Roots II. Effect of plant N-status on the kinetic parameters of nitrate efflux. J. Exp. Bot. 1986, 37, 1768–1779. [Google Scholar] [CrossRef]
- Siddiqi, M.Y.; Glass, A.D.M.; Ruth, T.J.; Rufty, T.W. Studies of the Uptake of Nitrate in Barley I. Kinetics of 13NO3− Influx. Plant Physiol. 1990, 93, 1426–1432. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.; Travis, R.L.; Huffaker, R.C. Comparative Kinetics and Reciprocal Inhibition of Nitrate and Nitrite Uptake in Roots of Uninduced and Induced Barley (Hordeum vulgare L.) Seedlings. Plant Physiol. 1992, 99, 1124–1133. [Google Scholar] [CrossRef] [PubMed]
- Touraine, B.; Glass, A. NO3− and ClO3− Fluxes in the chl1-5 Mutant of Arabidopsis thaliana (Does the CHL1-5 Gene Encode a Low-Affinity NO3− Transporter?). Plant Physiol. 1997, 114, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Filleur, S.; Dorbe, M.-F.; Cerezo, M.; Orsel, M.; Granier, F.; Gojon, A.; Daniel-Vedele, F. An Arabidopsis T-DNA mutant affected in Nrt2 genes is impaired in nitrate uptake. FEBS Letters 2001, 489, 220–224. [Google Scholar] [CrossRef]
- Li, W.; Wang, Y.; Okamoto, M.; Crawford, N.M.; Siddiqi, M.Y.; Glass, A.D.M. Dissection of the AtNRT2.1:AtNRT2.2 Inducible High-Affinity Nitrate Transporter Gene Cluster. Plant Physiol. 2007, 143, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Neame, K.D.; Richards, T.G. Elementary Kinetics of Membrane Carrier Transport; Blackwell Scientific Publications: Oxford/London, UK, 1972; pp. 80–94. [Google Scholar]
- Siddiqi, M.Y.; Glass, A.D.M.; Ruth, T.J.; Fernando, M. Studies of the regulation of nitrate influx by barley seedlings using 13NO3−. Plant Physiol. 1989, 90, 806–813. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.-H.; Huang, C.-Y.; Tsay, Y.-F. CHL1 Is a dual-affinity nitrate transporter of Arabidopsis involved in multiple phases of nitrate uptake. Plant Cell 1999, 11, 865–874. [Google Scholar] [CrossRef] [PubMed]
- Kiba, T.; Feria-Bourrellier, A.-B.; Lafouge, F.; Lezhneva, L.; Boutet-Mercey, S.; Orsel, M.; Bréhaut, V.; Miller, A.; Daniel-Vedele, F.; Sakakibara, H.; et al. The Arabidopsis nitrate transporter NRT2.4 plays a double role in roots and shoots of nitrogen-starved plants. Plant Cell 2012, 24, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Glass, A.D.M.; Kotur, Z. A Reevaluation of the Role of Arabidopsis NRT1.1 in high-affinity nitrate transport. Plant Physiol. 2013, 163, 1103–1106. [Google Scholar] [CrossRef] [PubMed]
- Lezhneva, L.; Kiba, T.; Feria-Bourrellier, A.-B.; Lafouge, F.; Boutet-Mercey, S.; Zoufan, P.; Sakakibara, H.; Daniel-Vedele, F.; Krapp, A. The Arabidopsis nitrate transporter NRT2.5 plays a role in nitrate acquisition and remobilization in nitrogen-starved plants. Plant J. 2014, 80, 230–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.L. Consistently modeling the combined effects of temperature and concentration on nitrate uptake in the ocean. J. Geophys. Res. 2011, 116. [Google Scholar] [CrossRef] [Green Version]
- Pasciak, W.J.; Gavis, J. Transport limitation of nutrient uptake in phytoplankton1: Transport limitation of uptake. Limnol. Ocean. 1974, 19, 881–888. [Google Scholar] [CrossRef]
- Franks, P.J.S. Planktonic ecosystem models: Perplexing parameterizations and a failure to fail. J. Plankton Res. 2009, 31, 1299–1306. [Google Scholar] [CrossRef]
- Sommer, U. Nitrate- and silicate-competition among antarctic phytoplankton. Mar. Biol. 1986, 91, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Litchman, E.; Klausmeier, C.A.; Schofield, O.M.; Falkowski, P.G. The role of functional traits and trade-offs in structuring phytoplankton communities: Scaling from cellular to ecosystem level. Ecol. Lett. 2007, 10, 1170–1181. [Google Scholar] [CrossRef] [PubMed]
- Slot, J.C.; Hallstrom, K.N.; Matheny, P.B.; Hibbett, D.S. Diversification of NRT2 and the Origin of Its Fungal Homolog. Mol. Biol. Evol. 2007, 24, 1731–1743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, S.M.; Plant, J.N.; Worden, A.Z. The mixed lineage nature of nitrogen transport and Assimilation in marine eukaryotic phytoplankton: A case study of micromonas. Mol. Biol. Evol. 2010, 27, 2268–2283. [Google Scholar] [CrossRef] [PubMed]
- Rogato, A.; Amato, A.; Iudicone, D.; Chiurazzi, M.; Ferrante, M.I.; d’Alcalà, M.R. The diatom molecular toolkit to handle nitrogen uptake. Mar. Genom. 2015, 25 Pt 1, 95–108. [Google Scholar] [CrossRef]
- Kang, L.-K.; Wang, H.-F.; Chang, J. Diversity of Phytoplankton nitrate transporter sequences from isolated single cells and mixed samples from the East China Sea and mRNA quantification. Appl. Environ. Microbiol. 2011, 77, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Bonachela, J.A.; Raghib, M.; Levin, S.A. Dynamic model of flexible phytoplankton nutrient uptake. Proc. Natl. Acad. Sci. USA 2011, 108, 20633–20638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldman, J.; Carpenter, E. A Kinetic approach to the effect of temperature on algal growth. Limnol. Oceanogr. 1974, 19, 756–766. [Google Scholar] [CrossRef]
- Harrison, P.J.; Parslow, J.S.; Conway, H.L. Determination of nutrient uptake kinetic parameters: A comparison of methods. Mar. Ecol. Prog. Ser. 1989, 52, 301–312. [Google Scholar] [CrossRef]
- Berg, H.C.; Purcell, E.M. Physics of chemoreception. Biophys. J. 1977, 20, 193–219. [Google Scholar] [CrossRef] [Green Version]
- Zwanzig, R. Diffusion-controlled ligand binding to spheres partially covered by receptors: An effective medium treatment. Proc. Natl. Acad. Sci. USA 1990, 87, 5856–5857. [Google Scholar] [CrossRef] [PubMed]
- Bonachela, J.A.; Allison, S.D.; Martiny, A.C.; Levin, S.A. A model for variable phytoplankton stoichiometry based on cell protein regulation. eScholarship 2013, 10, 4341–4356. [Google Scholar] [Green Version]
- Delhon, P.; Gojon, A.; Tillard, P.; Passama, L. Diurnal regulation of NO3− uptake in soybean plants I. Changes in NO3− influx, efflux, and N utilization in the plant during the day/night cycle. J. Exp. Bot. 1995, 46, 1585–1594. [Google Scholar] [CrossRef]
- Beuve, N.; Rispail, N.; Laine, P.; Cliquet, J.-B.; Ourry, A.; Deunff, E.L. Putative role of γ-aminobutyric acid (GABA) as a long-distance signal in up-regulation of nitrate uptake in Brassica napus L. Plant Cell Environ. 2004, 27, 1035–1046. [Google Scholar] [CrossRef]
- Malagoli, P.; Laine, P.; Le Deunff, E.; Rossato, L.; Ney, B.; Ourry, A. Modeling nitrogen uptake in oilseed rape cv capitol during a growth cycle using influx kinetics of root nitrate transport systems and field experimental data. Plant Physiol. 2004, 134, 388–400. [Google Scholar] [CrossRef] [PubMed]
- Moore, D.H. A study of age group track and field records to relate age and running speed. Nature 1975, 253, 264. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Pan, W.L.; Koenig, R.T. Integrated root system age in relation to plant nutrient uptake activity. Agron. J. 1998, 90, 505–510. [Google Scholar] [CrossRef]
- Warncke, D.D.; Barber, S.A. Nitrate uptake effectiveness of four plant species. J. Environ. Qual. 1974. [Google Scholar] [CrossRef]
- Edwards, P.H.; Barber, S.A. Phosphorus uptake rate of soybean roots as influenced by plant age, root trimming, and solution P concentration. Agron. J. 1976, 68, 973–975. [Google Scholar]
- Bath, K.K.S.; Brereton, A.J.; Nye, P.H. The possibility of predicting solute uptake and plant growth response from independently measured soil and plant characteristics VII. The growth and nitrate uptake of rape in soil at two nitrate concentrations and a comparison of the results with model predictions. Plant Soil 1979, 53, 169–191. [Google Scholar]
- van der Werf, A.; Kooijman, A.; Welschen, R.; Lambers, H. Respiratory energy costs for the maintenance of biomass, for growth and for ion uptake in roots of Carex diandra and Carex acutiformis. Physiologia Plantarum 1988, 72, 483–491. [Google Scholar] [CrossRef]
- Lambers, H.; Atkin, O.K.; Millenaar, F.F. Respiratory patterns in roots in relation to their functioning. In Plant Roots. The Hidden Half; Waisel, Y., Eshel, A., Kafkaki, K., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 1996; pp. 323–362. [Google Scholar]
- Ceccon, C.; Tagliavini, M.; Schmitt, A.O.; Eissenstat, D.M. Untangling the effects of root age and tissue nitrogen on root respiration in Populus tremuloides at different nitrogen supply. Tree Physiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Zwieniecki, M.A.; Thompson, M.V.; Holbrook, N.M. Understanding the hydraulics of porous pipes: Tradeoffs between water uptake and root length utilization. J. Plant Growth Regul. 2002, 21, 315–323. [Google Scholar] [CrossRef]
- Biondini, M. Allometric scaling laws for water uptake by plant roots. J. Theor. Biol. 2008, 251, 35–59. [Google Scholar] [CrossRef] [PubMed]
- Zarebanadkouki, M.; Meunier, F.; Couvreur, V.; Cesar, J.; Javaux, M.; Carminati, A. Estimation of the hydraulic conductivities of lupine roots by inverse modelling of high-resolution measurements of root water uptake. Ann. Bot. 2016, 118, 853–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vetterlein, D.; Doussan, C. Root age distribution: How does it matter in plant processes? A focus on water uptake. Plant Soil 2016, 407, 145–160. [Google Scholar] [CrossRef]
- Drew, M.C.; Saker, L.R. Nutrient Supply and the Growth of the Seminal Root System in Barley II.Localized, compensatory increases in lateral root growth and rates of nitrate uptake when nitrate supply is restricted to only part of the root system. J. Exp. Bot. 1975, 26, 79–90. [Google Scholar] [CrossRef]
- Orieux, C.; Demarest, G.; Decau, M.-L.; Beauclair, P.; Bataillé, M.-P.; Le Deunff, E. Changes in 15NO3- Availability and Transpiration Rate Are Associated With a Rapid Diurnal Adjustment of Anion Contents as Well as 15N and Water Fluxes Between the Roots and Shoots. Front. Plant Sci. 2018, 9, 1751. [Google Scholar] [CrossRef] [PubMed]
- Litchman, E.; Klausmeier, C.A.; Yoshiyama, K. Contrasting size evolution in marine and freshwater diatoms. Proc. Natl. Acad. Sci. USA 2009, 106, 2665–2670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daufresne, M.; Lengfellner, K.; Sommer, U. Global warming benefits the small in aquatic ecosystems. Proc. Natl. Acad. Sci. USA 2009, 106, 12788–12793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peter, K.H.; Sommer, U. Phytoplankton Cell Size: Intra- and Interspecific Effects of Warming and Grazing. PLoS ONE 2012, 7, e49632. [Google Scholar] [CrossRef] [PubMed]
- Buis, R. Contribution à la théorie logistique de la croissance: Structure temporelle et potentiel de croissance. C. R. Biol. 2003, 326, 729–738. [Google Scholar] [CrossRef]
- Buis, R.; Lück, J. Sur la notion de limite interne et externe dans les fonctions de croissance monotones. Une reformulation de l’équation logistique. C. R. Biol. 2006, 329, 880–891. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phase | N Uptake Duration (in %) | Amount of N Uptake ( in %) | ||||
|---|---|---|---|---|---|---|
| Fertilization levels | N0 0 kg N ha−1 | N1 135 kg N ha−1 | N2 272 kg N ha−1 | N0 0 kg N ha−1 | N1 135 kg N ha−1 | N2 272 kg N ha−1 |
| P1 | 44.0 | 41.2 | 42.6 | 22.8 | 4 | 0.7 |
| P2 | 4.1 | 2.7 | 2.7 | 26.0 | 21.8 | 23 |
| P3 | 4.1 | 2.7 | 1.4 | 29.5 | 33.4 | 22.4 |
| P4 | 47.8 | 53.3 | 53.3 | 21.8 | 40.7 | 53.9 |
| NO3− Uninduced Plants | NO3− Induced plants (1 mM 6–24 h) | |||
| References | Km (μmol) | Vmax (μmol h−1 (DW or FW) g−1) | Km (μmol) | Vmax (μmol h−1 (Dry or Fresh Weight)g−1) |
| Hordeum vulgare | ||||
| [65] | nd | nd | 7–62–86 * | 0.5–1.5–2.5 *(DW) |
| [66] | 6.4–7.8 | 21.7–22.4 (FW) | 12–14 | 43.9–48.7 (FW) |
| [67] | 20 | 0.34 (FW) | 79 | 9.4 (FW) |
| [68] | 6 | 0.82 (FW) | 36 | 8.4 (FW) |
| Arabidopsis thaliana | ||||
| [69] [70] [71] | nd nd nd | nd nd nd | 92 52.2 14.1 | 1 (FW) 147 (DW) 11.16 (FW) § |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le Deunff, E.; Malagoli, P.; Decau, M.-L. Modelling Nitrogen Uptake in Plants and Phytoplankton: Advantages of Integrating Flexibility into the Spatial and Temporal Dynamics of Nitrate Absorption. Agronomy 2019, 9, 116. https://doi.org/10.3390/agronomy9030116
Le Deunff E, Malagoli P, Decau M-L. Modelling Nitrogen Uptake in Plants and Phytoplankton: Advantages of Integrating Flexibility into the Spatial and Temporal Dynamics of Nitrate Absorption. Agronomy. 2019; 9(3):116. https://doi.org/10.3390/agronomy9030116
Chicago/Turabian StyleLe Deunff, Erwan, Philippe Malagoli, and Marie-Laure Decau. 2019. "Modelling Nitrogen Uptake in Plants and Phytoplankton: Advantages of Integrating Flexibility into the Spatial and Temporal Dynamics of Nitrate Absorption" Agronomy 9, no. 3: 116. https://doi.org/10.3390/agronomy9030116
APA StyleLe Deunff, E., Malagoli, P., & Decau, M. -L. (2019). Modelling Nitrogen Uptake in Plants and Phytoplankton: Advantages of Integrating Flexibility into the Spatial and Temporal Dynamics of Nitrate Absorption. Agronomy, 9(3), 116. https://doi.org/10.3390/agronomy9030116