Impact of Nitrogen Fertilization on Tuber Yield, Sugar Composition and Nitrogen Uptake of Two Yacon (Smallanthus sonchifolius Poepp. & Endl.) Genotypes


Abstract
:1. Introduction
2. Materials and Methods
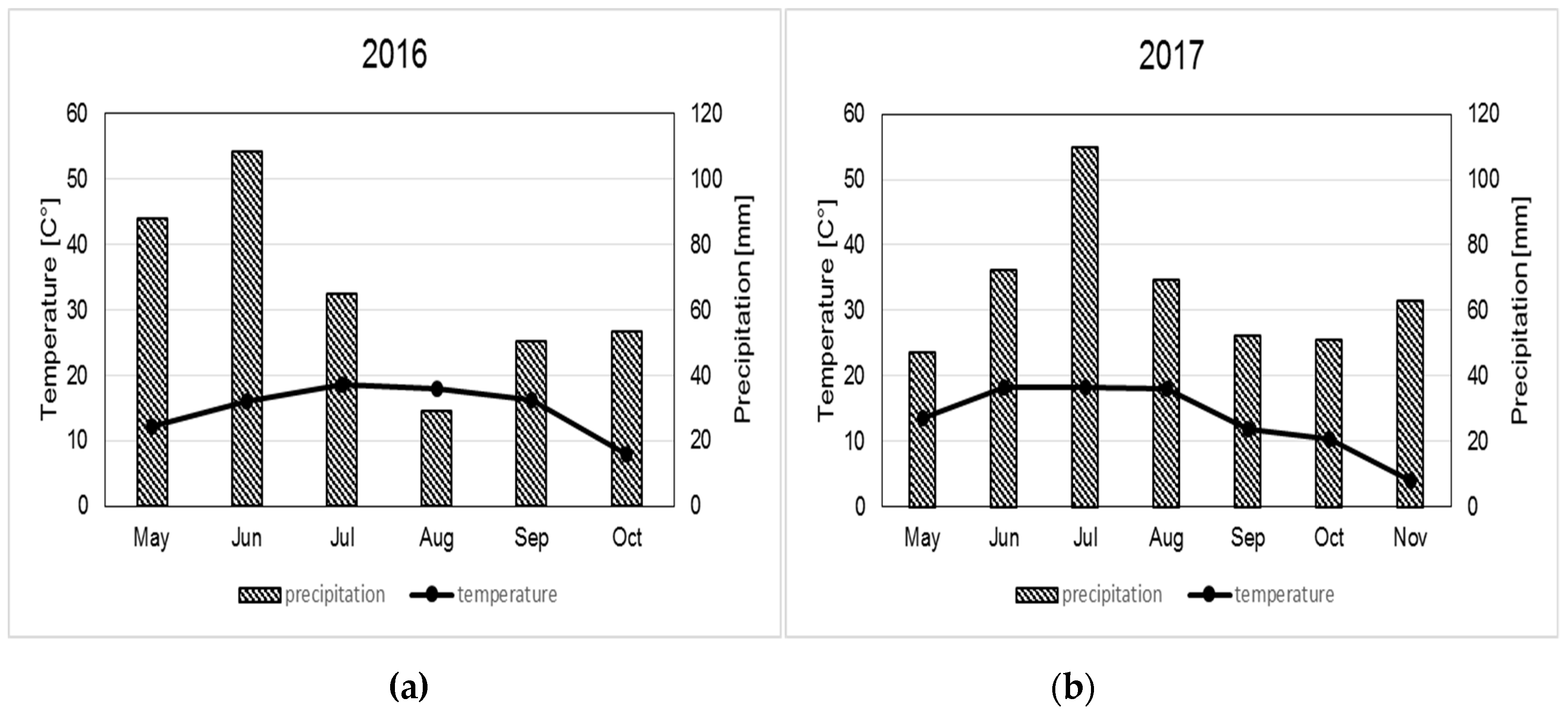
2.1. Field Site and Experimental Design
2.2. Treatments
2.3. Field Measurements and Sample Preparation
2.4. Data Collection
2.5. Statistical Analysis
3. Results
3.1. Tuber Yield
3.2. Sugar Composition
3.3.1. Nitrogen Concentration and Nitrogen Uptake
3.3.2. Indicators for Plant Efficiency
4. Discussion
4.1. Tuber Yield
4.2. Sugar Composition
4.3. Nitrogen Concentration and Uptake
4.4. Indicators for Plant Efficiency
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
| Genotype | Glucose [% DM] | Total Monosaccharides [% DM] | Total Sugar Content [% DM] | N Uptake Aboveground Biomass | N Uptake Total Plant [kg ha−1] | % N Rhizome | % N Tuber |
|---|---|---|---|---|---|---|---|
| B | 7.86a ± 0.4 | 16.31a ± 0.7 | 64.03a ± 0.8 | 126.39b ± 9.8 | 173.33b ± 12.0 | 1.52a ± 0.04 | 0.89a ± 0.02 |
| R | 2.91b ± 0.4 | 5.38b ± 0.7 | 41.35b ± 0.8 | 173.77a ± 9.8 | 236.58a ± 12.0 | 1.32b ± 0.04 | 0.76b ± 0.02 |
| Year × Genotype | Tuber Yield [t ha−1 FM] | Tuber Yield [t ha−1 DM) | Fructose [% DM] | N Uptake Tuber [kg ha−1] | % N Aboveground Biomass |
|---|---|---|---|---|---|
| 2016 × B | 20.42bA ± 1.4 | 2.26bB ± 0.2 | 1.87bA ± 0.4 | 22.55aA ± 3.9 | 3.47aA ± 0.06 |
| 2016 × R | 19.08bA ± 1.4 | 3.19bA ± 0.2 | n.d. | 33.80bA ± 5.0 | 3.19aB ± 0.07 |
| 2017 × B | 46.15aB ± 2.0 | 4.93aB ± 0.3 | 3.70aA ± 0.3 | 25.61aB ± 3.9 | 3.34aA ± 0.06 |
| 2017 × R | 63.23aA ± 2.0 | 10.75aA ± 0.3 | 0.13aB ± 0.3 | 69.11aA ± 5.0 | 2.19aA ± 0.07 |
| Fertilizer | N Uptake Aboveground Biomass [kg ha−1] | N Uptake Total Plant [kg ha−1] | % N Rhizome | PNUtES |
|---|---|---|---|---|
| 0 | 122.39b ± 12.0 | 167.62b ± 14.7 | 1.29b ± 0.05 | 13.77a ± 0.9 |
| 40 | 173.44a ± 12.0 | 227.54a ± 14.7 | 1.47a ± 0.05 | 10.26b ± 0.9 |
| 80 | 154.41ab ± 12.0 | 219.71a ± 14.7 | 1.52a ± 0.05 | 10.93b ± 0.9 |
| Year × Fertilizer | Tuber Yield [t ha−1 FM] | Tuber Yield [t ha−1 DM] | Total Sugar Content [% DM] | % N Aboveground Biomass |
|---|---|---|---|---|
| 2016*0 | 22.05bA ± 1.7 | 2.89bA ± 0.2 | 63.65aA ± 1.5 | 3.27aB ± 0.07 |
| 2016*40 | 20.77bAB ± 1.8 | 3.08bA ± 0.2 | 54.00aB ± 1.5 | 3.19aB ± 0.07 |
| 2016*80 | 16.44bB ± 1.7 | 2.21bB ± 0.2 | 53.57aB ± 1.5 | 3.77aA ± 0.07 |
| 2017*0 | 52.91aA ± 2.6 | 7.48aB ± 0.3 | 45.91bA ± 1.2 | 2.68bA ± 0.08 |
| 2017*40 | 52.37aA ± 2.6 | 7.50aB ± 0.3 | 49.36bA ± 1.2 | 2.74bA ± 0.08 |
| 2017*80 | 58.80aA ± 2.5 | 8.54aA ± 0.3 | 49.66aA ± 1.2 | 2.65bA ± 0.08 |
| Genotype × Fertilizer | Tuber DM | % N Aboveground Biomass |
|---|---|---|
| B0 | 11.21bA ± 0.41 | 3.28aB ± 0.08 |
| B40 | 10.41bA ± 0.41 | 3.12aB ± 0.08 |
| B80 | 11.30bA ± 0.41 | 3.59aA ± 0.08 |
| R0 | 16.04aB ± 0.41 | 2.67bA ± 0.08 |
| R40 | 17.34aA ± 0.41 | 2.81bA ± 0.08 |
| R80 | 16.63aAB ± 0.41 | 2.81bA ± 0.08 |
References
- Ojansivu, I.; Ferreira, C.L.; Salminen, S. Yacon, a new source of prebiotic oligosaccharides with a history of safe use. Trends Food Sci. Technol. 2011, 22, 40–46. [Google Scholar] [CrossRef]
- Bredemann, G. Über Polymnia sonchifolia Poepp. et Endl. (P. edulis Wedd.), die Yacon—Erdbirne. Bot. Oeconomie 1948, 1, 65–85. [Google Scholar]
- Fernández, E.C.; Viehmannova, I.; Bechyne, M.; Lachman, J.; Milella, L. The cultication and phenological growth stage of yacon [Smallanthus sonchifolius (Poepp. et Endl.) H. Robinson]. Agric. Trop. Subtrop. 2007, 40, 71–77. [Google Scholar]
- Zardini, E. Ethnobotanical notes on “Yacon” Polymnia sonchifolia (Asteraceae). Econ. Bot. 1991, 45, 72–85. [Google Scholar] [CrossRef]
- Delgado, G.T.C.; da Silva Cunha Tamashiro, W.M.; Marostica, M.R., Jr.; Pastore, G.M. Yacon (Smallanthus sonchifolius): A functional food. Plant Foods Hum. Nutr. 2013, 68, 222–228. [Google Scholar] [CrossRef]
- Douglas, J.A.; Follett, J.M.; Douglas, M.H.; Deo, B.; Scheffer, J.J.C.; Littler, R.A.; Manley-Harris, M. Effect of environment and time of planting on the production and quality of yacon (Smallanthus sonchifolius) storage roots. N. Z. J. Crop Hortic. Sci. 2007, 35, 107–116. [Google Scholar] [CrossRef]
- Valentova, K.; Lebeda, A.; Dolezalova, I.; Jirovsky, D.; Simonovska, B.; Vovk, I.; Kosina, P.; Gasmanova, N.; Dziechciarkova, M.; Ulrichova, J. The biological and chemical variability of yacon. J. Agric. Food Chem. 2006, 54, 1347–1352. [Google Scholar] [CrossRef]
- Graefe, S.; Hermann, M.; Manrique, I.; Golombek, S.; Buerkert, A. Effects of post-harvest treatments on the carbohydrate composition of yacon roots in the Peruvian Andes. Field Crop. Res. 2004, 86, 157–165. [Google Scholar] [CrossRef]
- Manrique, I.; Parraga, A.; Hermann, M. Yacon Syrup: Principles and Processing; Centro International de la Papa: Lima, Peru, 2005. [Google Scholar]
- Lachman, J.; Fernández, E.C.; Orsák, M. Yacon [Smallanthus sonchifolia (Poepp. et Endl.) H. Robinson] chemical composition and use—A review. Plant Soil Environ. 2003, 6, 283–290. [Google Scholar] [CrossRef]
- Goto, K.; Fukai, K.; Hikida, J.; Nanjo, F.; Hara, Y. Isolation and structural analysis of oligosaccharides from yacon (Polymnia sonchifolia). Biosci. Biotechnol. Biochem. 2014, 59, 2346–2347. [Google Scholar] [CrossRef]
- Mokrani, K.; Hamdi, K.; Tarchoun, N. Potato (Solanum Tuberosum L.) response to nitrogen, phosphorus and potassium fertilization rates. Commun. Soil Sci. Plant Anal. 2018, 49, 1314–1330. [Google Scholar] [CrossRef]
- Sumiyanto, J.; Dayan, F.E.; Cerdeira, A.L.; Wang, Y.-H.; Khan, I.A.; Moraes, R.M. Oligofructans content and yield of yacon (Smallanthus sonchifolius) cultivated in Mississippi. Sci. Hortic. 2012, 148, 83–88. [Google Scholar] [CrossRef]
- Doo, H.S. Changes in sugar contents and storability of yacon under different storage conditions. Korean J. Crop Sci. 2000, 45, 300–304. [Google Scholar]
- Doo, H.S. Changes of chemical composition in tuberous root of yacon by different curing conditions. Korean J. Crop Sci. 2000, 45, 79–82. [Google Scholar]
- Fernandes, A.M.; Campos, L.G.; Senna, M.S.; da Silva, C.L.; Assunção, N.S. Yield and nitrogen use efficiency of sweet potato in response to cover crop and nitrogen management. Agron. J. 2018, 110, 2004. [Google Scholar] [CrossRef]
- Märländer, B.; Hoffmann, C.; Koch, H.-J.; Ladewig, E.; Merkes, R.; Petersen, J.; Stockfisch, N. Environmental situation and yield performance of the sugar beet crop in Germany: Heading for sustainable development. J. Agron. Crop Sci. 2003, 11, 201–226. [Google Scholar] [CrossRef]
- Pocock, T.O.; Milford, G.F.J.; Armstrong, M.J. Storage root quality in sugarbeet in relation to nitrogen uptake. J. Agric. Sci. 1990, 115, 355. [Google Scholar] [CrossRef]
- Carter, J.N.; Traveller, D.J. Effect of time and amount of nitrogen uptake on sugarbeet growth and yield. Agron. J. 1981, 73, 665–671. [Google Scholar] [CrossRef]
- Sah, R.N.; Geng, S.; Puri, Y.P.; Rubatzky, V.E. Evaluation of four crops for nitrogen utilization and carbohydrate yield. Fertil. Res. 1987, 13, 55–70. [Google Scholar] [CrossRef]
- Maltas, A.; Dupuis, B.; Sinaj, S. Yield and quality response of two potato cultivars to nitrogen fertilization. Potato Res. 2018, 61, 97–114. [Google Scholar] [CrossRef]
- Gastal, F.; Lemaire, G. N uptake and distribution in crops: An agronomical and ecophysiological perspective. J. Exp. Bot. 2002, 53, 789–799. [Google Scholar] [CrossRef]
- Goulding, K. Nitrate leaching from arable and horticultural land. Soil Use Manag. 2000, 16, 145–151. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2006. Ein Rahmen für Internationale Klassifikation, Korrelation und Kommunikation. Erstes Update 2007. Deutsche Ausgabe; BGR: Hannover, Germany, 2008. [Google Scholar]
- VDLUFA. Methodenbuch Band II.1. Die Untersuchung von Bo den; VDLUFA-Verlag: Darmstadt, Germany, 2004; Available online: https://www.vdlufa.de/Methodenbuch/index.php?option=com_content&view=article&id=9&Itemid=115&lang=de (accessed on 15 January 2019).
- Campbell, J.M.; Bauer, L.L.; Fahey, G.C.; Hogarth, A.J.C.L.; Wolf, B.W.; Hunter, D.E. Selected fructooligosaccharide (1-kestose, nystose, and 1 F-β-fructofuranosylnystose) composition of foods and feeds. J. Agric. Food Chem. 1997, 45, 3076–3082. [Google Scholar] [CrossRef]
- Wolfinger, R. Covariance structure selection in general mixed models. Commun. Stat. Simul. Comput. 1993, 22, 1079–1106. [Google Scholar] [CrossRef]
- Piepho, H.P. An algorithm for a letter-based representation of all-pairwise comparisons. Biometrical J. 2004, 13, 456–466. [Google Scholar] [CrossRef]
- Fernández, E.C.; Viehmannová, I.; Bechyne, M.; Lachman, J.; Milella, L. Yacon [Smallanthus sonchifolius (Poeppig & Endlicher) H. Robinson]: A new crop in the Central Europe. Plant Soil Environ. 2006, 12, 564–570. [Google Scholar]
- Leemans, R.; Cramer, W.P. The IIASA database for mean monthly values of temperature, precipitation, and cloudiness on a global terrestrial grid. IIASA Res. Rep. 1991, 91. [Google Scholar]
- Douglas, J.A. Research and development of yacon production in New Zealand. Acta Hortic. 2005, 670, 79–85. [Google Scholar] [CrossRef]
- De Mastro, G.; Manolio, G.; Marzi, V. Jerusalem artichoke (Helianthus tuberosus L.) and chicory (Chicorium intybus L.): Potential crops for inulin production in the mediterranean area. Acta Hortic. 2004, 629, 365–374. [Google Scholar] [CrossRef]
- Meijer, W.J.M.; Mathijssen, E.W.J.M.; Borm, G.E.L. Crop characteristics and inulin production of Jerusalem artichoke and chicory. Stud. Plant Sci. 1993, 3, 29–38. [Google Scholar]
- Kai, G.; Tie-Xia, Z.; Qi-Bing, W. Nitrogen fertilization, irrigation, and harvest times affect biomass and energy value of Helianthus tuberosus L. J. Plant Nutr. 2016, 39, 1906–1914. [Google Scholar] [CrossRef]
- Losavio, N.; Lamascese, N.; Vonella, A.V. Water requirements and nitrogen fertilization in Jerusalem artichoke (Helianthus tuberosus L.) grown under mediterranean conditions. Acta Hortic. 1997, 1, 205–209. [Google Scholar] [CrossRef]
- Stahr, K.; Kandeler, E.; Herrmann, L.; Streck, T. Bodenkunde und Standortlehre, 2nd ed.; Eugen Ulmar KG: Stuttgart, Germany, 2012. [Google Scholar]
- Engels, T.; Kuhlmann, H. Effect of the rate of N fertilizer on apparent net mineralization of N during and after cultivation of cereal and sugar beet crops. J. Plant Nutr. Soil. Sci. 1993, 156, 149–154. [Google Scholar] [CrossRef]
- Manrique, I.; Hermann, M.; Bernet, T. Yacon Fact Sheet; International Potato Center (CIP): Lima, Peru, 2004. [Google Scholar]
- Douglas, J.A. Maximising fructo-oligosacharide production in yacon. Agron. N. Z. 2002, 32, 49–55. [Google Scholar]
- Hermann, M.; Freire, I.; Pazos, C. Compositional diversity of the yacon storage root. CIP Program Rep. 1997, 98, 425–432. [Google Scholar]
- Leidi, E.O.; Altamirano, A.M.; Mercado, G.; Rodriguez, J.P.; Ramos, A.; Alandia, G.; Sørensen, M.; Jacobsen, S.-E. Andean roots and tubers crops as sources of functional foods. J. Funct. Foods 2018, 51, 86–93. [Google Scholar] [CrossRef]
- Campos, D.; Betalleluz-Pallardel, I.; Chirinos, R.; Aguilar-Galvez, A.; Noratto, G.; Pedreschi, R. Prebiotic effects of yacon (Smallanthus sonchifolius Poepp. & Endl), a source of fructooligosaccharides and phenolic compounds with antioxidant activity. Food Chem. 2012, 135, 1592–1599. [Google Scholar] [CrossRef]
- Hermann, M. Andean Roots and Tubers: Ahipa, Arracacha, Maca and Yacon; International Plant Genetic Resources Institute: Rome, Italy, 1997; pp. 191–243. [Google Scholar]
- Sprague, H.B.; Farris, N.F.; Colby, W.G. Effect of soil conditions and treatment on yields of tubers and sugar from the American artichoke (Helianthus tuberosus). J. Am. Soc. Agron. 1935, 27, 392–399. [Google Scholar] [CrossRef]
- Lachman, J.; Havrland, B.; Fernandez, E.C.; Dudjak, J. Saccharides of yacon [Smallanthus sonchifolius (Poepp. et Endl.) H. Robinson] tubers and rhizomes and factors affecting their content. Plant Soil Environ. 2004, 50, 383–390. [Google Scholar] [CrossRef]
- Ohyama, T.; Ito, O.; Yasuyoshi, S.; Ikarashi, T.; Minamisawa, K.; Kubota, M.; Tsukihashi, T.; Asami, T. Composition of storage carbohydrate in tubers of yacon (Polymnia sonchifolia). Soil Sci. Plant Nutr. 1990, 36, 167–171. [Google Scholar] [CrossRef]
- Delgado, G.T.C.; Thomé, R.; Gabriel, D.L.; Tamashiro, W.M.; Pastore, G.M. Yacon (Smallanthus sonchifolius)-derived fructooligosaccharides improves the immune parameters in the mouse. Nutr. Res. 2012, 32, 884–892. [Google Scholar] [CrossRef] [PubMed]
- Fukai, K.; Miyazaki, S.; Nanjo, F.; Hara, Y. Distribution of carbohydrates and related enzyme activities in yacon (polymnia sonchifolia). Soil Sci. Plant Nutr. 1993, 39, 567–571. [Google Scholar] [CrossRef]
- Khajehei, F.; Merkt, N.; Claupein, W.; Graeff-Hoenninger, S. Yacon (Smallanthus sonchifolius Poepp. & Endl.) as a novel source of health promoting compounds: Antioxidant activity, phytochemicals and sugar content in flesh, peel, and whole tubers of seven cultivars. Molecules 2018, 23, 278. [Google Scholar] [CrossRef]
- Minamisawa, K.; Ohyama, T. Fluctuations of oligofructan contents in tubers of yacon (Polymnia. sonchifolia) during growth and storage. Jpn. Soc. Soil Sci. Plant Nutr. 1991, 62, 621–627. [Google Scholar]
- Dorrell, D.G.; Chubey, B.B. Irrigation, fertilizer, harvest dates and storage effects on the reducing sugar and fructose concentrations of Jerusalem artichoke tubers. Can. J. Plant Sci. 1997, 57, 591–596. [Google Scholar] [CrossRef]
- Soja, G.; Dersch, G.; Praznik, W. Harvest dates, fertilizer and varietal effects on yield, concentration and molecular distribution of fructan in Jerusalem artichoke (Helianthus tuberosus L.). J. Agron. Crop Sci. 1990, 165, 181–189. [Google Scholar] [CrossRef]
- Ojala, J.C.; Stark, J.C.; Kleinkopf, G.E. Influence of irrigation and nitrogen management on potato yield and quality. Am. Potato J. 1990, 67, 29–43. [Google Scholar] [CrossRef]
- Richter, C. Agrikulturchemie und Pflanzenernährung; Margraf Publishers GmbH: Weikersheim, Germany, 2005. [Google Scholar]
- Lebeda, A.; Dolezalova, I.; Valentova, K.; Gasmanova, N.; Dziechciarkova, M.; Ulrichova, J. Yacon (Smallanthus sonchifolius)—A traditional crop of the Andean Indians as a challenge for the future—The news about biological variation and chemical substances content. Acta Hortic. 2008, 127–136. [Google Scholar] [CrossRef]
- Ana, R.; Snezana, C.; Marina, T.; Viorica, T.; Gabriel, I.; Demetra, S.; Carmen, G.; Slobodan, J. Comparative analysis of the chemical composition of Heliantus tuberosus L. growing in Serbia and Romania. Serb. J. Exp. Clin. Res. 2013, 14, 9–12. [Google Scholar] [CrossRef]
- Slimestad, R.; Seljaasen, R.; Meijer, K.; Skar, S.L. Norwegian-grown Jerusalem artichoke (Helianthus tuberosus L.): Morphology and content of sugars and fructo-oligosaccharides in stems and tubers. J. Sci. Food Agric. 2010, 90, 956–964. [Google Scholar] [CrossRef]
- Puangbut, D.; Jogloy, S.; Vorasoot, N.; Srijaranai, S.; Kesmala, T.; Holbrook, C.C.; Patanothai, A. Influence of planting date and temperature on inulin content in Jerusalem artichoke (Helianthus tuberosus L.). Aust. J. Crop Sci. 2012, 6, 1159–1165. [Google Scholar]
- Zotarelli, L.; Rens, L.R.; Cantliffe, D.J.; Stoffella, P.J.; Gergela, D.; Burhans, D. Rate and timing of nitrogen fertilizer application on potato ‘FL1867’. Part I: Plant nitrogen uptake and soil nitrogen availability. Field Crop. Res. 2015, 183, 246–256. [Google Scholar] [CrossRef]
- Märländer, B. Influence of nitrogen supply on yield and quality of sugar beet. J. Plant Soil Sci. 1990, 153, 327–332. [Google Scholar] [CrossRef]
- Landwirtschaftliches Technologiezentrum Augustenberg. Merkblätter für die Umweltgerechte Landbewirtschaftung: Kartoffeln, Anbau, Düngung, Pflanzenschutz; Landwirtschaftliches Technologiezentrum Augustenberg: Karlsruhe, Germany, 2011; pp. 1–12. [Google Scholar]
- Battilani, A.; Plauborg, F.L.; Hansen, S.; Dolezal, F.; Mazurczyk, W.; Bizik, J.; Coutinho, J. Nitrogen uptake and nitrogen use efficiency of fertigated potatoes. Acta Hortic. 2008, 61–67. [Google Scholar] [CrossRef]
- Bélanger, G.; Walsh, J.R.; Richards, J.E.; Milburn, P.H.; Ziadi, N. Nitrogen fertilization and irrigation affects tuber characteristics of two potato cultivars. Am. J. Potato Res. 2002, 79, 269–279. [Google Scholar] [CrossRef]
- Seiler, G.J. Protein and mineral concentrations in tubers of selected genotypes of wild and cultivated Jerusalem-artichoke (Helianthus tuberosus, asteraceae). Econ. Bot. 1990, 44, 322–335. [Google Scholar] [CrossRef]
- Joern, B.C.; Vitosh, M.L. Influence of applied nitrogen on potato part 1: Yield, quality, and nitrogen uptake. Am. Potato J. 1995, 72, 51–63. [Google Scholar] [CrossRef]
- Kleijn, D.; Treier, U.A.; Müller-Schärer, H. The importance of nitrogen and carbohydrate storage for plant growth of the alpine herb Veratrum album. New Phytol. 2005, 166, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Laufer, D.; Koch, H.-J. Growth and yield formation of sugar beet (Beta vulgaris L.) under strip tillage compared to full width tillage on silt loam soil in Central Europe. Eur. J. Agron. 2017, 82, 182–189. [Google Scholar] [CrossRef]
- Houba, V.J.G.; Huijbregts, A.W.M.; Wilting, P.; Novozamsky, I.; Gort, G. Sugar yield, nitrogen uptake by sugar beet and optimal nitrogen fertilization in relation to nitrogen soil analyses and several additional factors. Biol. Fertil. Soils 1995, 19, 55–59. [Google Scholar] [CrossRef]
- Laufer, D.; Nielsen, O.; Wilting, P.; Koch, H.-J.; Märländer, B. Yield and nitrogen use efficiency of fodder and sugar beet (Beta vulgaris L.) in contrasting environments of northwestern Europe. Eur. J. Agron. 2016, 73, 124–132. [Google Scholar] [CrossRef]
- Mengel, K. Turnover of organic nitrogen in soils and its availability to crops. Plant Soil 1996, 181, 83–93. [Google Scholar] [CrossRef]

| Abbreviation Treatment | Genotype | Fertilization (kg N ha−1) |
|---|---|---|
| B0 | brown-shelled | 0 |
| B40 | brown-shelled | 40 |
| B80 | brown-shelled | 80 |
| R0 | red-shelled | 0 |
| R40 | red-shelled | 40 |
| R80 | red-shelled | 80 |
| Year | Transplanting Date | Date of Measurement (DAP) | Harvest | |||
|---|---|---|---|---|---|---|
| 1. | 2. | 3. | 4. | |||
| 2016 | 18.05 | 27.07. (70) | 30.08. (104) | 27.09. (132) | 26.10. (161) | 28.10. (163) |
| 2017 | 11.05 | 27.07. (77) | 24.08. (105) | 21.09. (133) | 23.10. (165) | 02.11. (175) |
| Treatment | Tuber Yield | |||
| Fresh Matter [t ha−1 FM] | DM [%] | Dry Matter [t ha−1 DM] | ||
| 2016 | ||||
| B0 | 24.06 ± 2.45 | 11.83 ± 0.67 | 2.81 ± 0.3 | |
| B40 | 19.57 ± 2.45 | 10.36 ± 0.67 | 2.04 ± 0.3 | |
| B80 | 17.62 ± 2.45 | 11.28 ± 0.67 | 1.95 ± 0.3 | |
| R0 | 20.04 ± 2.45 | 15.03 ± 0.67 | 3.01 ± 0.3 | |
| R40 | 21.97 ± 2.45 | 17.68 ± 0.67 | 3.90 ± 0.3 | |
| R80 | 15.26 ± 2.45 | 16.25 ± 0.67 | 2.47 ± 0.3 | |
| 2017 | ||||
| B0 | 45.38 ± 3.55 | 10.58 ± 0.47 | 4.72 ± 0.5 | |
| B40 | 42.81 ± 3.67 | 10.45 ± 0.47 | 4.51 ± 0.5 | |
| B80 | 50.25 ± 3.55 | 11.31 ± 0.47 | 5.67 ± 0.5 | |
| R0 | 60.44 ± 3.66 | 17.04 ± 0.47 | 10.35 ± 0.5 | |
| R40 | 61.93 ± 3.55 | 17.00 ± 0.47 | 10.50 ± 0.5 | |
| R80 | 67.32 ± 3.55 | 17.00 ± 0.47 | 11.40 ± 0.5 | |
| Results of statistical analysis | ||||
| Factor | DF | p-Value | ||
| Year × Rep | 4 | <0.0001 | 0.1060 | 0.0048 |
| Year | 1 | <0.0001 | 0.6375 | <0.0001 |
| G | 1 | <0.0001 | <0.0001 | <0.0001 |
| Year × G | 1 | <0.0001 | 0.1272 | <0.0001 |
| F | 2 | 0.8731 | 0.6895 | 0.8622 |
| Year × F | 2 | 0.0137 | 0.6356 | 0.0057 |
| G × F | 2 | 0.4782 | 0.0486 | 0.2412 |
| Year × G × F | 2 | 0.9438 | 0.0698 | 0.5333 |
| Treatment | Fructose | Glucose | Sucrose | GF2 | GF3 | GF4 | Total FOS | Total Sugar | |
|---|---|---|---|---|---|---|---|---|---|
| 2016 | |||||||||
| B0 | 1.38 ± 0.6 | 4.67 ± 1.2 | 6.15aA ± 0.4 | 23.16aA ± 1.0 | 19.04aA ± 0.6 | 12.22bAB ± 0.5 | 54.42aA ± 1.8 | 66.62aB ± 2.16 | |
| B40 | 2.63 ± 0.6 | 7.87 ± 1.2 | 7.09aA ± 0.4 | 25.07aA ± 1.0 | 19.30aA ± 0.6 | 11.77aB ± 0.5 | 56.15aA ± 1.8 | 73.74aA ± 2.16 | |
| B80 | 1.60 ± 0.6 | 5.44 ± 1.2 | 6.58aA ± 0.4 | 23.85aA ± 1.0 | 20.85aA ± 0.6 | 13.52aA ± 0.5 | 58.22aA ± 1.8 | 71.84aAB ± 2.16 | |
| R0 | n.d. | 2.48 ± 1.2 | 4.22bA ± 0.4 | 14.59bA ± 1.0 | 20.75aA ± 0.6 | 18.64aA ± 0.5 | 53.99aA ± 1.8 | 60.69aA ± 2.16 | |
| R40 | n.d. | 1.68 ± 1.2 | 1.84bB ± 0.4 | 8.20bB ± 1.0 | 12.05bB ± 0.6 | 10.50aB ± 0.5 | 30.75bB ± 1.8 | 34.27bB ± 2.16 | |
| R80 | n.d. | 2.01 ± 1.2 | 1.94bB ± 0.4 | 7.38bB ± 1.0 | 12.43bB ± 0.6 | 11.55bB ± 0.5 | 31.36bB ± 1.8 | 35.31bB ± 2.16 | |
| 2017 | |||||||||
| B0 | 4.01 ± 0.4 | 10.66 ± 0.9 | 4.78aA ± 0.2 | 12.89aA ± 1.0 | 13.45aB ± 0.6 | 9.86bB ± 0.4 | 36.21aB ± 1.2 | 55.65aA ± 1.70 | |
| B40 | 3.92 ± 0.4 | 10.50 ± 0.9 | 5.02aA ± 0.2 | 14.22aA ± 1.0 | 14.49aAB ± 0.6 | 10.64bAB ± 0.4 | 39.35aAB ± 1.2 | 58.80aA ± 1.70 | |
| B80 | 3.18 ± 0.4 | 8.00 ± 0.9 | 4.37aA ± 0.2 | 15.09aA ± 1.0 | 15.54aA ± 0.6 | 11.39bA ± 0.4 | 42.02aA ± 1.2 | 57.56aA ± 1.70 | |
| R0 | 0.08 ± 0.4 | 3.28 ± 0.9 | 2.32bA ± 0.2 | 6.05bA ± 1.0 | 12.23aB ± 0.6 | 12.22aB ± 0.4 | 30.50bB ± 1.2 | 36.18bB ± 1.70 | |
| R40 | 0.17 ± 0.4 | 3.85 ± 0.9 | 2.68bA ± 0.2 | 6.87bA ± 1.0 | 13.12bAB ± 0.6 | 13.24aAB ± 0.4 | 33.23bAB ± 1.2 | 39.92bAB ± 1.70 | |
| R80 | 0.15 ± 0.4 | 4.16 ± 0.9 | 2.63bA ± 0.2 | 7.10bA ± 1.0 | 13.72bA ± 0.6 | 13.99aA ± 0.4 | 34.81bA ± 1.2 | 41.75bA ± 1.70 | |
| Results of statistical analysis | |||||||||
| Factor | DF | p-Value | |||||||
| Year × Rep | 4 | 0.2147 | 0.3612 | 0.0594 | 0.1646 | 0.1028 | 0.0843 | 0.1656 | 0.2596 |
| Year | 1 | 0.0064 | 0.0003 | <0.0001 | <0.0001 | <0.0001 | 0.0003 | <0.0001 | <0.0001 |
| G | 1 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Year × G | 1 | 0.0156 | 0.1196 | 0.0003 | <0.0001 | <0.0001 | 0.0104 | <0.0001 | 0.0006 |
| F | 2 | 0.5143 | 0.3753 | 0.1348 | 0.3982 | 0.0010 | 0.0001 | 0.0085 | 0.0523 |
| Year × F | 2 | 0.7009 | 0.7453 | 0.0834 | 0.0025 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| G × F | 2 | 0.5364 | 0.2099 | 0.0100 | 0.0019 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Year × G × F | 2 | 0.6348 | 0.2230 | 0.0021 | 0.0095 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Treatment | % N in Different Plant Parts | |||
| Aboveground Biomass | Rhizome | Tuber | ||
| 2016 | ||||
| B0 | 3.35 ± 0.11 | 1.46 ± 0.15 | 0.95 ± 0.07 | |
| B40 | 3.06 ± 0.11 | 1.43 ± 0.15 | 0.96 ± 0.07 | |
| B80 | 4.01 ± 0.11 | 1.75 ± 0.15 | 1.07 ±0.07 | |
| R0 | 3.19 ± 0.11 | 1.25 ± 0.15 | 0.82 ± 0.07 | |
| R40 | 3.31 ± 0.11 | 1.64 ± 0.15 | 0.73 ± 0.07 | |
| R80 | 3.52 ± 0.11 | 1.47 ± 0.15 | 0.91 ± 0.07 | |
| 2017 | ||||
| B0 | 3.21 ± 0.12 | 1.45 ± 0.05 | 0.91 ± 0.03 | |
| B40 | 3.18 ± 0.12 | 1.52 ± 0.05 | 0.75 ± 0.03 | |
| B80 | 3.19 ± 0.12 | 1.50 ± 0.05 | 0.78 ± 0.03 | |
| R0 | 2.14 ± 0.12 | 0.98 ± 0.05 | 0.61 ± 0.03 | |
| R40 | 2.31 ± 0.12 | 1.27 ± 0.05 | 0.77 ± 0.03 | |
| R80 | 2.10 ± 0.12 | 1.34 ± 0.05 | 0.71 ± 0.03 | |
| Results of statistical analysis | ||||
| Factor | DF | p-Value | ||
| Year × Rep | 4 | 0.0275 | 0.3944 | 0.2193 |
| Year | 1 | <0.0001 | 0.0297 | 0.0002 |
| G | 1 | <0.0001 | 0.0088 | 0.0018 |
| Year × G | 1 | <0.0001 | 0.1362 | 0.2133 |
| F | 2 | 0.0097 | 0.0262 | 0.1710 |
| Year × F | 2 | 0.0010 | 0.9263 | 0.1677 |
| G × F | 2 | 0.0233 | 0.1500 | 0.7892 |
| Year × G × F | 2 | 0.2954 | 0.2032 | 0.1834 |
| Treatment | N Uptake (kg ha−1) of Different Plant Parts | ||||
| Aboveground Biomass | Rhizome | Tuber | Total Plant | ||
| 2016 | |||||
| B0 | 88.60 ± 28.6 | 13.31 ± 4.8 | 19.06 ± 6.7 | 120.98 ± 32.3 | |
| B40 | 169.42 ± 28.6 | 22.35 ± 4.8 | 18.77 ± 6.7 | 210.56 ± 32.3 | |
| B80 | 110.19 ± 28.6 | 12.20 ± 4.8 | 29.80 ± 6.7 | 152.18 ± 32.3 | |
| R0 | 118.81 ± 28.6 | 10.84 ± 4.8 | 31.41 ± 6.7 | 161.07 ± 32.3 | |
| R40 | 200.64 ± 28.6 | 20.46 ± 4.8 | 15.33 ± 6.7 | 236.42 ± 32.3 | |
| R80 | 152.62 ± 28.6 | 17.03 ± 4.8 | 30.08 ± 6.7 | 199.73 ± 32.3 | |
| 2017 | |||||
| B0 | 131.00 ± 18.3 | 12.74 ± 6.6 | 33.36 ± 8.7 | 177.10 ± 26.3 | |
| B40 | 130.30 ± 18.3 | 22.66 ± 6.6 | 29.52 ± 8.7 | 182.48 ± 26.3 | |
| B80 | 128.81 ± 18.3 | 29.32 ± 6.6 | 38.53 ± 8.7 | 196.66 ± 26.3 | |
| R0 | 151.14 ± 18.3 | 9.30 ± 6.6 | 50.88 ± 8.7 | 211.31 ± 26.3 | |
| R40 | 193.38 ± 18.3 | 15.07 ± 6.6 | 72.25 ± 8.7 | 280.71 ± 26.3 | |
| R80 | 226.03 ± 18.3 | 20.03 ± 6.6 | 84.19 ± 8.7 | 330.25 ± 26.3 | |
| Results of statistical analysis | |||||
| Factor | DF | p-Value | |||
| Year × Rep | 4 | 0.4321 | 0.4120 | 0.0149 | 0.1587 |
| Year | 1 | 0.1666 | 0.5236 | <0.0001 | 0.0087 |
| G | 1 | 0.0033 | 0.3313 | 0.0004 | 0.0014 |
| Year × G | 1 | 0.3708 | 0.3095 | 0.0019 | 0.1509 |
| F | 2 | 0.0252 | 0.0856 | 0.0683 | 0.0191 |
| Year × F | 2 | 0.1153 | 0.2629 | 0.2708 | 0.1873 |
| G × F | 2 | 0.4400 | 0.9508 | 0.7648 | 0.4534 |
| Year × G × F | 2 | 0.6343 | 0.7186 | 0.1287 | 0.5043 |
| Treatment | TNUtED | PNUtES | TNUtES | |
| 2016 | ||||
| B0 | 80.33 ± 40.3 | 13.38 ± 1.7 | 96.32 ± 20.9 | |
| B40 | 114.69 ± 40.3 | 5.68 ± 1.7 | 75.14 ± 20.9 | |
| B80 | 35.29 ± 40.3 | 6.74 ± 1.7 | 73.74 ± 20.9 | |
| R0 | 105.48 ± 40.3 | 9.56 ± 1.7 | 50.96 ± 20.9 | |
| R40 | 184.95 ± 40.3 | 7.25 ± 1.7 | 95.19 ± 20.9 | |
| R80 | 103.48 ± 40.3 | 6.11 ± 1.7 | 37.77 ± 20.9 | |
| 2017 | ||||
| B0 | 83.96 ± 7.0 | 14.88 ± 1.8 | 78.54 ± 3.6 | |
| B40 | 134.16 ± 7.0 | 13.70 ± 1.8 | 84.41 ± 3.6 | |
| B80 | 129.35 ± 7.0 | 15.71 ± 1.8 | 80.68 ± 3.6 | |
| R0 | 164.51 ± 7.0 | 17.26 ± 1.8 | 72.29 ± 3.6 | |
| R40 | 131.23 ± 7.0 | 14.41 ± 1.8 | 56.32 ± 3.6 | |
| R80 | 85.43 ± 7.0 | 15.17 ± 1.8 | 60.59 ± 3.6 | |
| Results of statistical analysis | ||||
| Factor | DF | p-Value | ||
| Year × Rep | 4 | 0.2726 | 0.9535 | 0.0789 |
| Year | 1 | 0.3445 | <0.0001 | 0.9472 |
| G | 1 | 0.0912 | 0.9606 | 0.0605 |
| Year × G | 1 | 0.2456 | 0.3926 | 0.9026 |
| F | 2 | 0.1022 | 0.0281 | 0.4142 |
| Year × F | 2 | 0.4356 | 0.2328 | 0.4548 |
| G × F | 2 | 0.6241 | 0.7200 | 0.5396 |
| Year × G × F | 2 | 0.1571 | 0.3422 | 0.1979 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamp, L.; Hartung, J.; Mast, B.; Graeff-Hönninger, S. Impact of Nitrogen Fertilization on Tuber Yield, Sugar Composition and Nitrogen Uptake of Two Yacon (Smallanthus sonchifolius Poepp. & Endl.) Genotypes. Agronomy 2019, 9, 151. https://doi.org/10.3390/agronomy9030151
Kamp L, Hartung J, Mast B, Graeff-Hönninger S. Impact of Nitrogen Fertilization on Tuber Yield, Sugar Composition and Nitrogen Uptake of Two Yacon (Smallanthus sonchifolius Poepp. & Endl.) Genotypes. Agronomy. 2019; 9(3):151. https://doi.org/10.3390/agronomy9030151
Chicago/Turabian StyleKamp, Larissa, Jens Hartung, Benjamin Mast, and Simone Graeff-Hönninger. 2019. "Impact of Nitrogen Fertilization on Tuber Yield, Sugar Composition and Nitrogen Uptake of Two Yacon (Smallanthus sonchifolius Poepp. & Endl.) Genotypes" Agronomy 9, no. 3: 151. https://doi.org/10.3390/agronomy9030151
APA StyleKamp, L., Hartung, J., Mast, B., & Graeff-Hönninger, S. (2019). Impact of Nitrogen Fertilization on Tuber Yield, Sugar Composition and Nitrogen Uptake of Two Yacon (Smallanthus sonchifolius Poepp. & Endl.) Genotypes. Agronomy, 9(3), 151. https://doi.org/10.3390/agronomy9030151