Water Stress Permanently Alters Shoot Architecture in Common Bean Plants



Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Set-Up
2.2. Plant Architecture Data
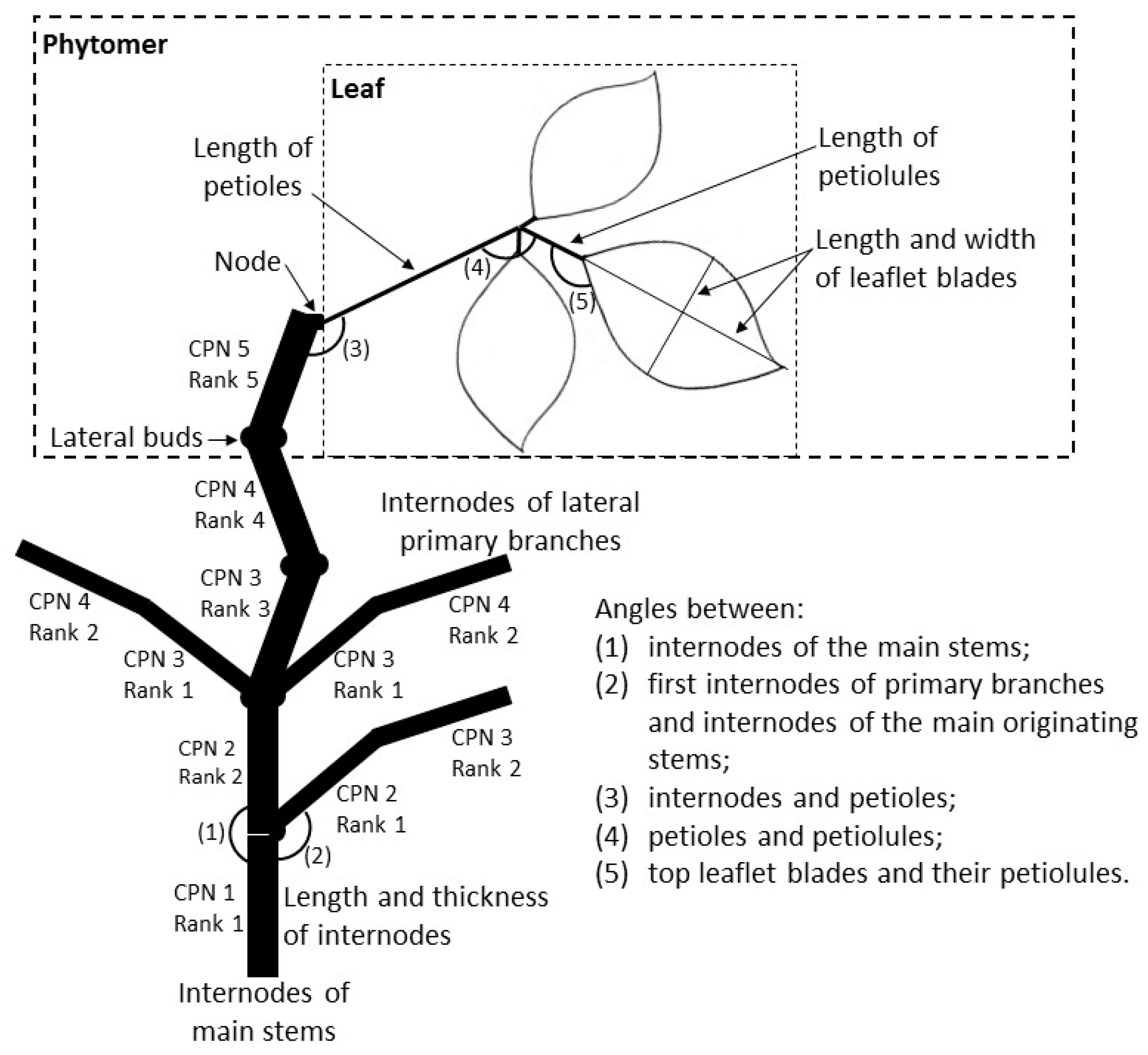
2.2.1. General Bean Plant Architecture
2.2.2. Air Temperature
2.2.3. Leaf Appearance
2.2.4. Final Organ Size, Number and Angles
2.3. Statistics
3. Results
3.1. Water Applied, Evapotranspiration and Yield Traits
3.2. Phyllochron
3.3. Number of Phytomers and Leaflet Blades
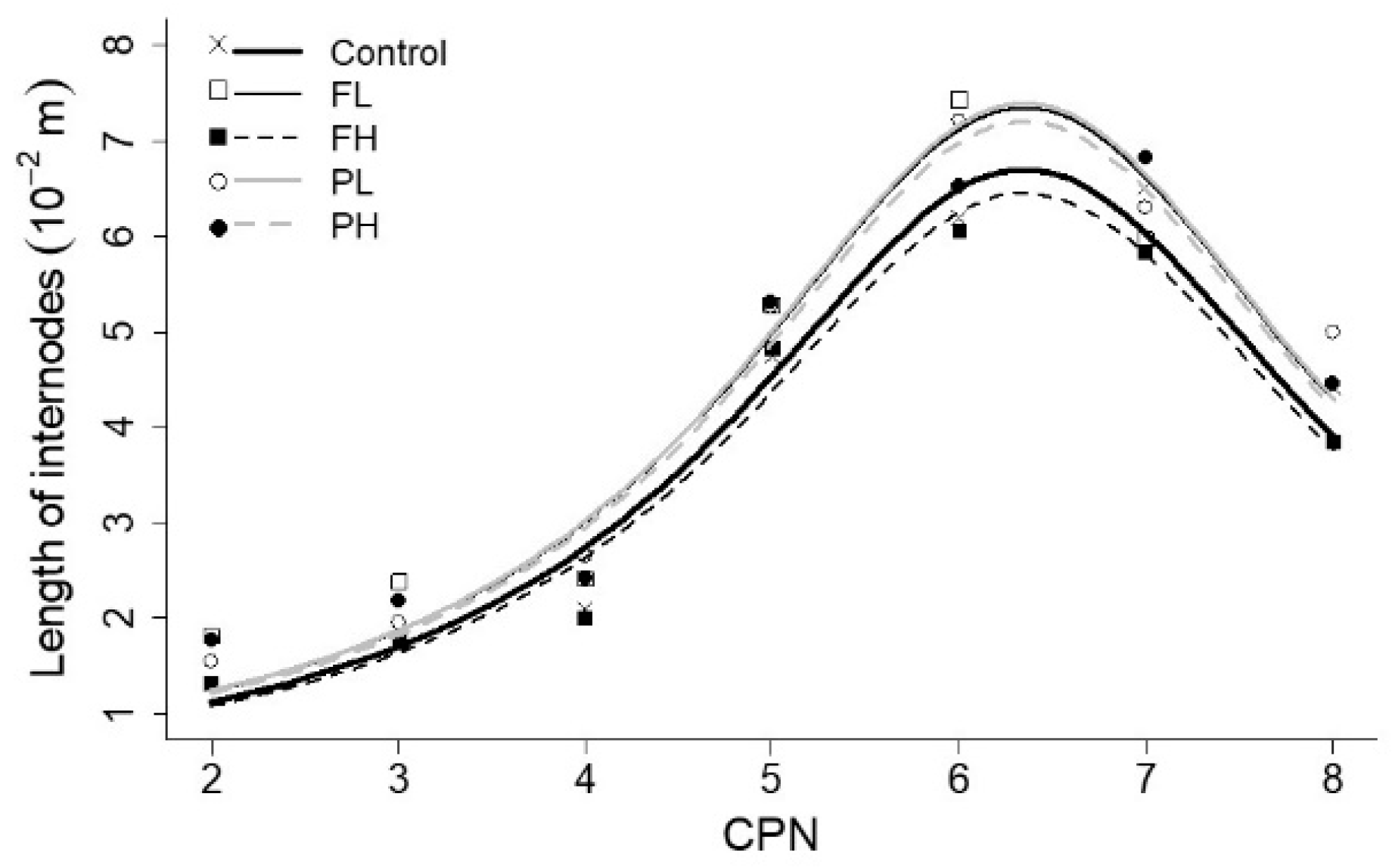
3.4. Size of Internodes
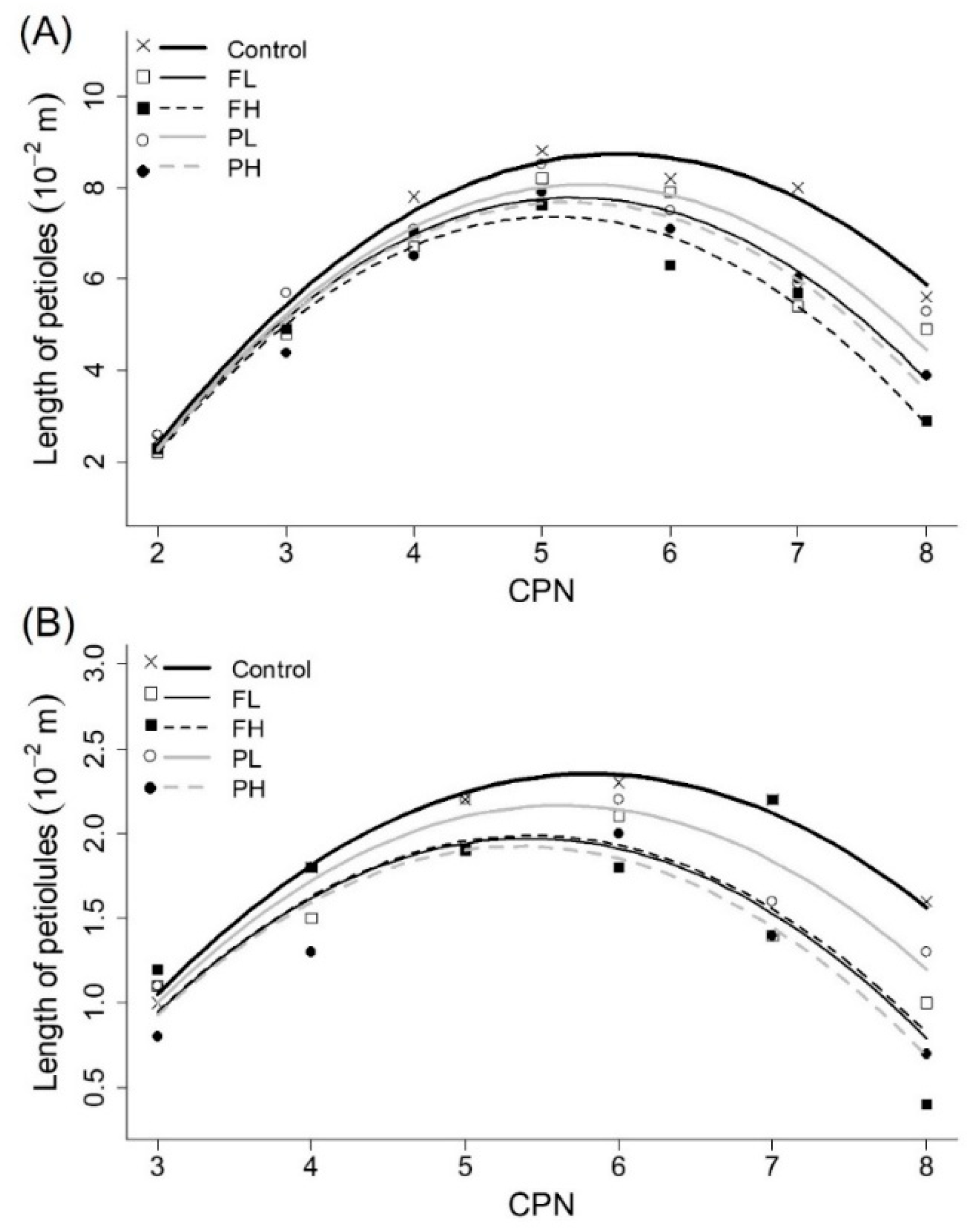
3.5. Lengths of Petioles and Petiolules
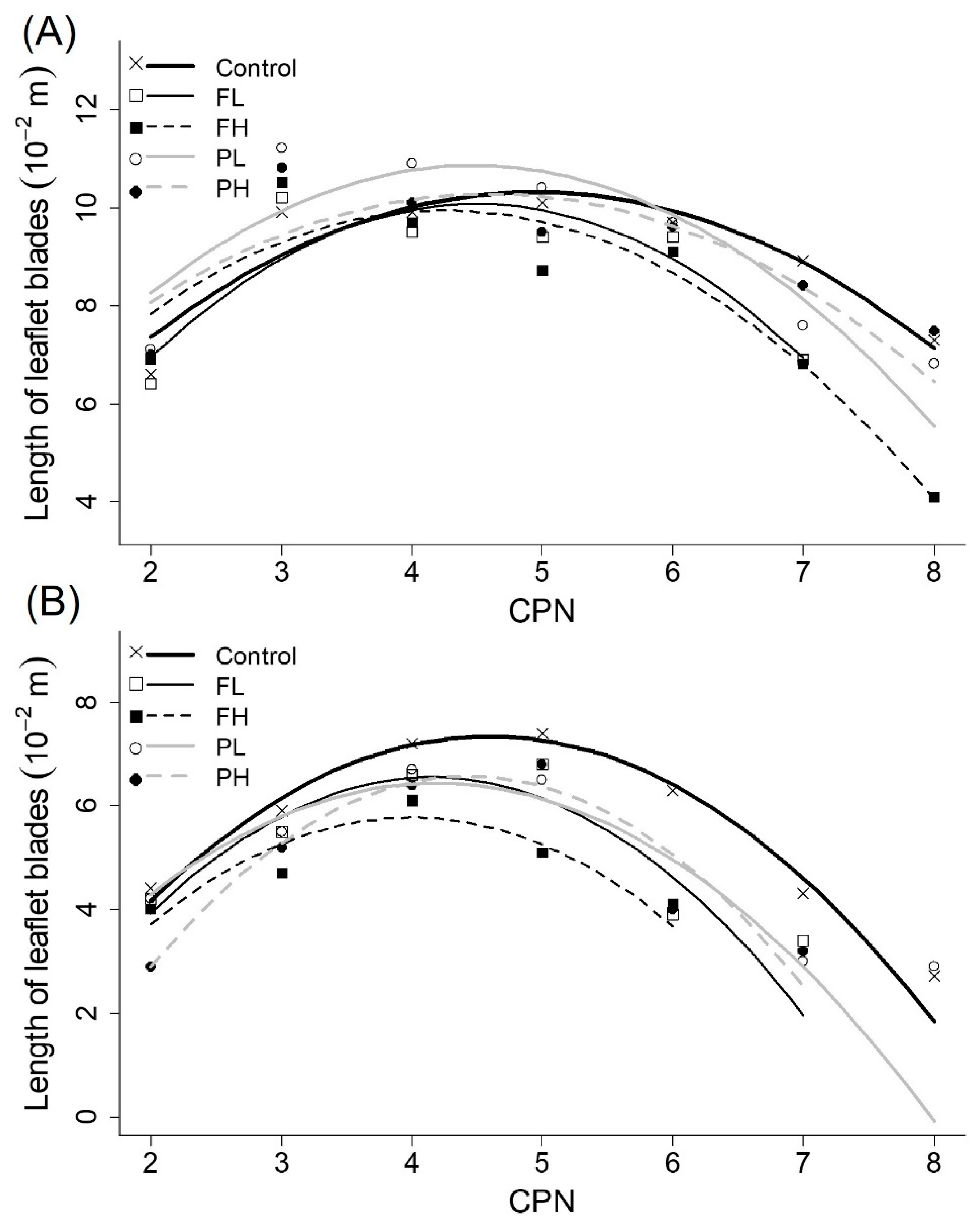
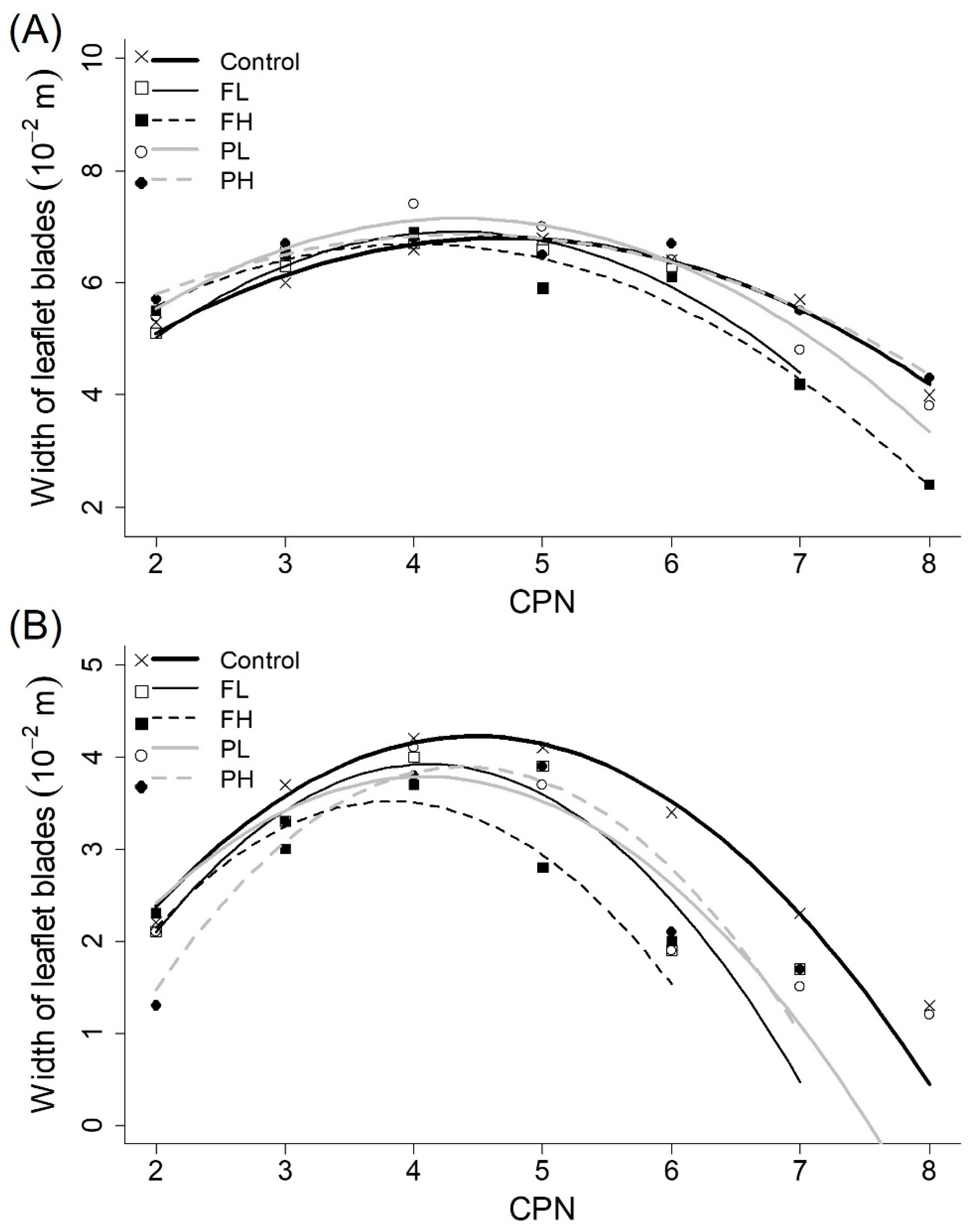
3.6. Size of Leaflet Blades
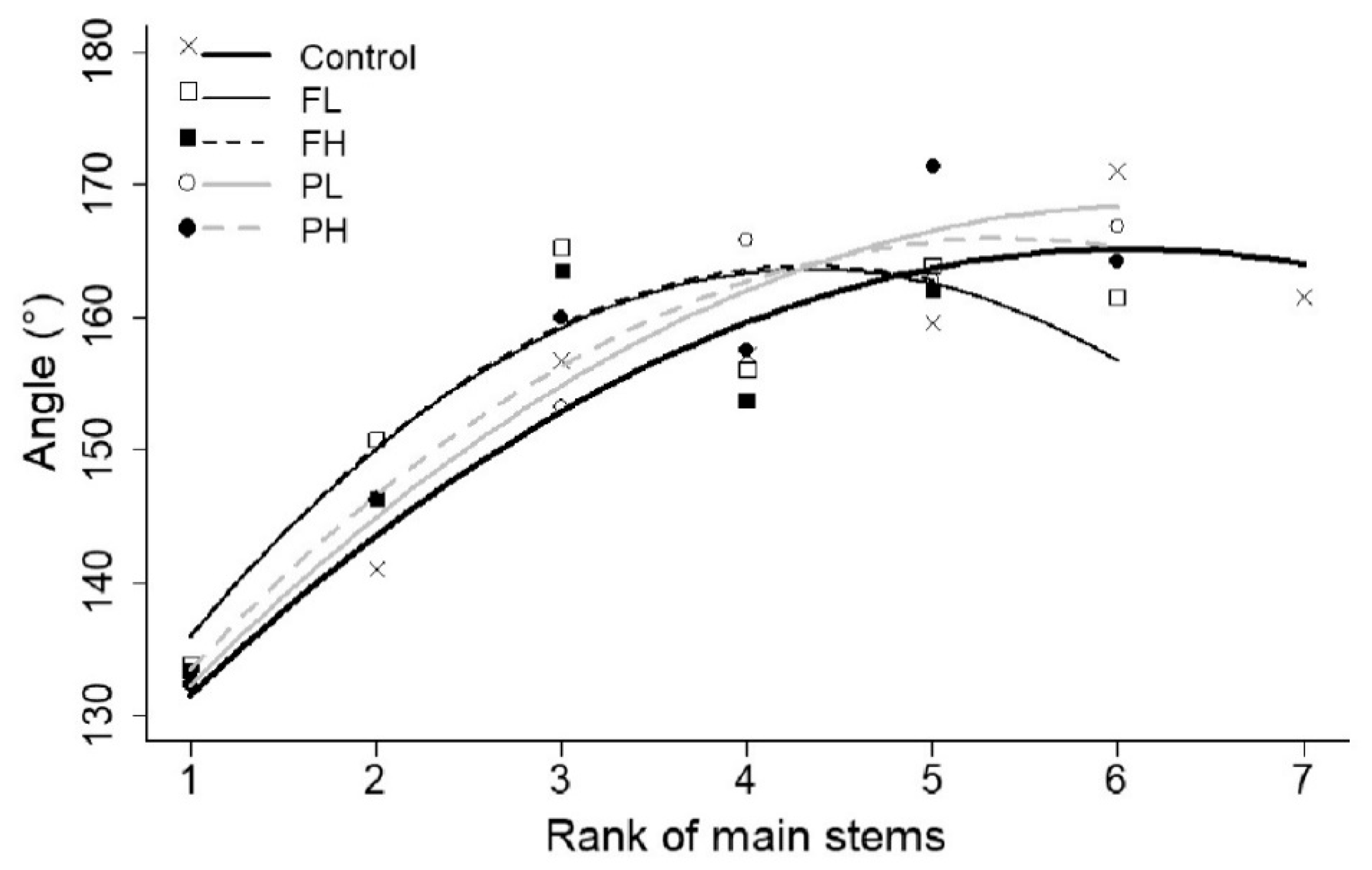
3.7. Angles
4. Discussion
4.1. Water Applied, Evapotranspiration and Yield Traits
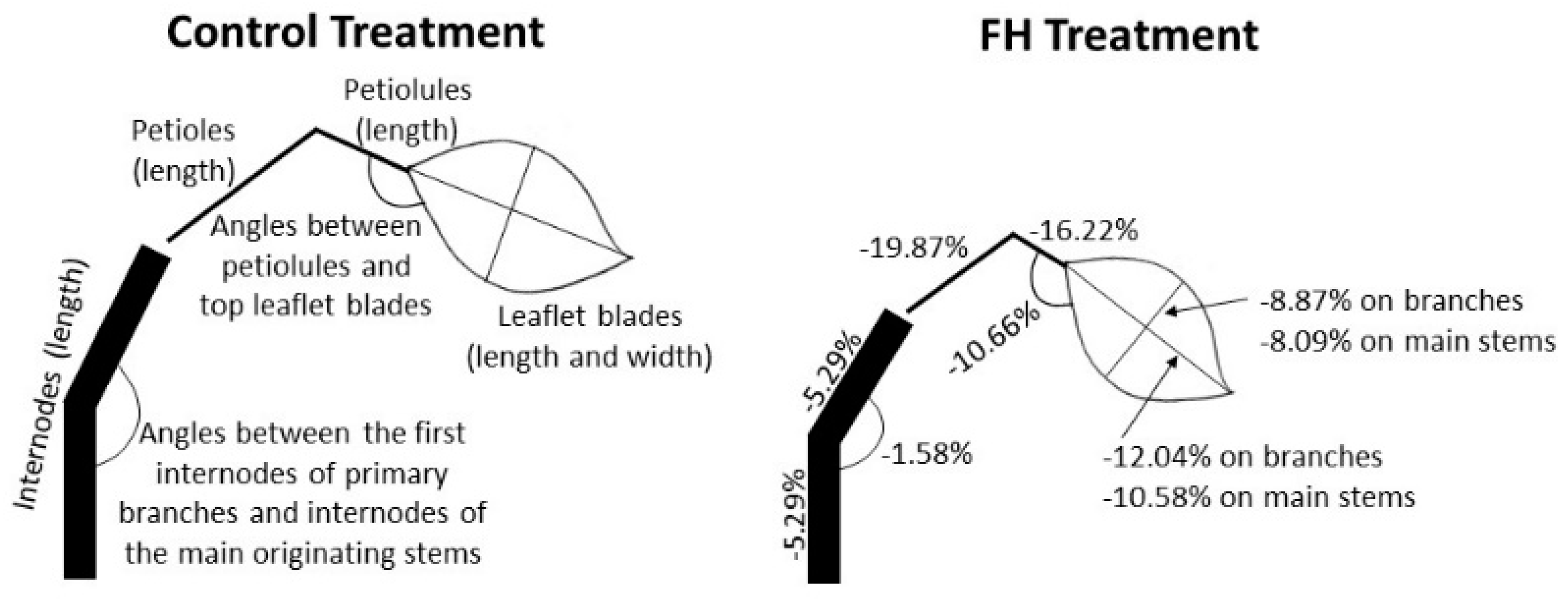
4.2. Development and Architecture
4.3. Outlook
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A. Dummy-Variable Regression Method
References
- Manschadi, A.M.; Christopher, J.; Voil, P.D.; Hammer, G.L. The role of root architectural traits in adaptation of wheat to water-limited environments. Func. Plant Biol. 2006, 33, 823–837. [Google Scholar] [CrossRef] [Green Version]
- Atta, B.M.; Mahmood, T.; Trethowan, R.M. Relationship between root morphology and grain yield of wheat in north-western NSW, Australia. Aust. J. Crop Sci. 2013, 7, 2108–2115. [Google Scholar]
- UNDP. Reducing Disaster Risk: A Challenge for Development. A Global Report. Available online: http://www.undp.org/cpr/whats_new/rdr_english.pdf (accessed on 14 January 2019).
- Helmer, M.; Hilhorst, D. Natural disasters and climate change. Disasters 2006, 30, 1–4. [Google Scholar] [CrossRef]
- Li, Y.; Ye, W.; Wang, M.; Yan, X. Climate change and drought: A risk assessment of crop-yield impacts. Climate Res. 2009, 39, 31–46. [Google Scholar] [CrossRef]
- IPCC. Summary for policymakers. In Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Field, C.B., Barros, V.R., Dokken, D.J., Mach, K.J., Mastrandrea, M.D., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., et al., Eds.; Cambridge University Press: Cambridge, UK, 2014; pp. 1–32. [Google Scholar]
- Dai, A.; Trenberth, K.E.; Qian, T. A global data set of Palmer Drought Severity Index for 1870–2002: Relationship with soil moisture and effects of surface warming. J. Hydrometeorol. 2004, 5, 1117–1130. [Google Scholar] [CrossRef]
- Rosegrant, M.W.; Cline, S.A. Global food security: Challenges and policies. Science 2003, 302, 1917–1919. [Google Scholar] [CrossRef]
- Ericksen, P.J. Conceptualizing food systems for global environmental change research. Glob. Environ. Chang. 2008, 18, 234–245. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology, 5th ed.; Sinauer Associates: Sunderland, UK, 2010; 782p. [Google Scholar]
- Monclus, R.; Dreyer, E.; Villar, M.; Delmotte, F.M.; Delay, D.; Pettit, J.M.; Barbaroux, C.; Thiec, D.; Brechet, C.; Brignolas, F. Impact of drought on productivity and water use efficiency in 29 genotypes of Populus deltoides x Populus nigra. New Phytol. 2006, 169, 765–777. [Google Scholar] [CrossRef]
- Haldimann, P.; Galle, A.; Feller, U. Impact of an exceptionally hot dry summer on photosynthetic traits in oak (Quercus pubescens) leaves. Tree Physiol. 2008, 28, 785–795. [Google Scholar] [CrossRef]
- Ruiz-Ramos, M.; Mínguez, M.I. ALAMEDA, a structural-functional model for faba bean crops: Morphological parameterization and verification. Ann. Bot. 2006, 97, 377–388. [Google Scholar] [CrossRef]
- Roujean, J.-L. A tractable physical model of shortwave radiation interception by vegetative canopies. J. Geophys. Res. 1996, 101, 9523–9532. [Google Scholar] [CrossRef]
- Archontoulis, S.V.; Vos, J.; Yin, X.; Bastiaans, L.; Danalatos, N.G.; Struik, P.C. Temporal dynamics of light and nitrogen distributions in canopies of sunflower, kenaf and cynara. Field Crops Res. 2011, 122, 186–198. [Google Scholar] [CrossRef]
- Atti, S.; Bonnel, R.; Prasher, S.; Smith, D.L. Response of soybean {Glycine max (L.) merr.} under chronic water deficit to LCO application during flowering and pod filling. Irrig. Drain. 2005, 54, 15–30. [Google Scholar] [CrossRef]
- Graham, P.H.; Vance, C.P. Legumes: Importance and constraints to greater use. Plant Physiol. 2003, 131, 872–877. [Google Scholar] [CrossRef]
- Mitchell, D.C.; Lawrence, F.R.; Hartman, T.J.; Curran, J.M. Consumption of dry beans, peas, and lentils could improve diet quality in the US population. J. Am. Diet. Ass. 2009, 109, 909–913. [Google Scholar] [CrossRef]
- Pachico, D. Trends in world common bean production. In Bean Production Problems in the Tropics; Schwartz, H.F., Pastor-Corrales, M.A., Eds.; International Center for Tropical Agriculture (CIAT): Cali, Colombia, 1989; pp. 1–8. [Google Scholar]
- Ramirez-Vallejo, P.; Kelly, I.D. Traits related to drought resistance in common bean. Euphytica 1998, 99, 127–136. [Google Scholar] [CrossRef]
- Broughton, W.J.; Hernandez, G.; Blair, M.; Beebe, S.; Gepts, P.; Vanderleyden, J. Beans (Phaseolus spp.)-model food legumes. Plant Soil 2003, 252, 55–128. [Google Scholar] [CrossRef]
- Petry, N.; Boy, E.; Worth, J.P.; Hurrel, R. The potential of the Common Bean (Phaseolus vulgaris) as a vehicle for iron biofortification. Nutrients 2015, 7, 1144–1173. [Google Scholar] [CrossRef] [PubMed]
- van Schoonhoven, A.; Voysest, O. Common beans in Latin America and their constraints. In Bean Production Problems in the Tropics, 2nd ed.; Schwartz, H.F., Pastor-Corrales, M.A., Eds.; International Center for Tropical Agriculture (CIAT): Cali, Colombia, 1989; pp. 33–37. [Google Scholar]
- Shellie-Dessert, K.; Bliss, F. Genetic improvement of food quality factors. In Common Beans: Research for Crop Improvement; van Schoonhoven, A., Voyses, O., Eds.; CAB International in association with International Center for Tropical Agriculture (CIAT): Cali, Colombia, 1991; pp. 649–679. [Google Scholar]
- Back, A.J. Irrigation of bean culture in the South of Santa Catarina state–Brazil. Rev. Tec. Amb. 2001, 7, 35–44. [Google Scholar]
- Wallace, D.H. Adaptation of Phaseolus to different environments. In Advances in Legume Science; Summerfield, R.J., Bunting, A.H., Eds.; Royal Botanic Gardens: Kew, UK, 1980; pp. 349–357. [Google Scholar]
- Halterlein, A.J. Bean. In Crop Water Relations; Teare, I.D., Peet, M.M., Eds.; Willey & Sons: New York, NY, USA, 1983; pp. 157–185. [Google Scholar]
- Rosales-Serna, R.; Kohashi-Shibata, J.; Acosta-Gallegos, J.A.; Trejo-Lopez, C.; Ortiz-Cereceres, J.; Kelly, J.D. Biomass distribution, maturity acceleration and yield in drought stressed common bean cultivars. Field Crops Res. 2004, 85, 203–211. [Google Scholar] [CrossRef]
- Boutraa, T.; Sanders, F.E. Effects of interactions of moisture regime and nutrient addition on nodulation and carbon partitioning in two cultivars of bean (Phaseolus vulgaris L.). J. Agron. Crop Sci. 2001, 186, 229–237. [Google Scholar] [CrossRef]
- Miller, D.E.; Burke, D.W. Response of dry beans to daily deficit sprinkler irrigation. Agron. J. 1983, 75, 775–778. [Google Scholar] [CrossRef]
- Silveira, P.M.; Steinmetz, S.; Guimarães, C.M.; Aidar, H.; Carvalho, J.R.P. Frequency and levels of irrigation in dry beans during winter season. Pesq. Agropec. Bras. 1984, 19, 219–223. [Google Scholar]
- Doorenbos, J.; Pruitt, W.O. Crop Water Requirements. Irrigation and Drainage Paper; n. 24; Food and Agriculture Organization of The United Nations: Rome, Italy, 1976; 189p. [Google Scholar]
- Costa-Franca, M.G.; Thi, A.T.; Pimentel, C.; Pereyra, R.O.; Zuily-Fodil, Y.; Laffray, D. Differences in growth and water relations among Phaseolus vulgaris cultivars in response to induced drought stress. Environ. Exp. Bot. 2000, 43, 227–237. [Google Scholar] [CrossRef]
- Acosta-Gallegos, J.A.; Adams, M.W. Plant traits and yield stability of dry bean (Phaseolus vulgaris L.) cultivars under drought stress. J. Agric. Sci. 1991, 117, 213–219. [Google Scholar] [CrossRef]
- Adams, M.W.; Coyne, D.P.; Davis, J.H.C.; Graham, P.H.; Francis, C.A. Common bean (Phaseolus vulgaris L.). In Grain Legume Crops; Summerfield, R.J., Roberts, E.H., Eds.; Collins: London, UK, 1985; pp. 433–476. [Google Scholar]
- Xia, M.Z. Effect of soil drought during the generative development phase on seed yield and nutrient uptake of faba bean (Vicia faba L.). Aust. J. Agric. Res. 1997, 48, 447–451. [Google Scholar] [CrossRef]
- Loss, S.P.; Siddique, K.H.M.; Tennant, D. Adaptation of faba bean (Vicia faba L.) to dryland Mediterranean-type environments I. Seed yield and yield components. Field Crops Res. 1997, 52, 17–28. [Google Scholar] [CrossRef]
- Mwanamwenge, J.; Loss, S.P.; Siddique, K.H.M.; Cocks, P.S. Effect of water stress during floral initiation, flowering and podding on the growth and yield of faba bean (Vicia faba L.). Eur. J. Agron. 1999, 11, 1–11. [Google Scholar] [CrossRef]
- Doorenbos, J.; Kassam, A.H. Yield Response to Water. Irrigation and Drainage Paper; n. 33; Food and Agriculture Organization of The United Nations: Rome, Italy, 1979; 306p. [Google Scholar]
- Pardossi, A.; Vernieri, P.; Tognoni, T. Involvement of abscisic acid in regulating water status in Phaseolus vulgaris L. during chilling. Plant Physiol. 1992, 100, 1243–1250. [Google Scholar] [CrossRef] [PubMed]
- Comstock, J.; Ehleringer, J. Stomatal response to humidity in common bean (Phaseolus vulgaris): Implications for maximum transpiration rate, water-use efficiency and productivity. Aust. J. Plant Physiol. 1993, 20, 669–691. [Google Scholar] [CrossRef]
- Njoka, E.M.; Muraya, M.M.; Okumu, M. The influences of plant density on yield and yield components of common beans (Phaseolus vulgaris L.). Agric. Trop. Subtrop. 2005, 38, 22–29. [Google Scholar]
- Barrios, A.N.; Hoogenboom, G.; Nesmith, D.S. Drought stress and the distribution of vegetative and reproductive traits of a bean cultivar. Scientia Agric. 2005, 62, 18–22. [Google Scholar] [CrossRef]
- Silva, A.O.; Lima, E.A.; Fidelis Filho, J.; Nóbrega, J.Q. Dry matter accumulation of beans grown in different densities in Lagoa Seca-PB. In Proceedings of the XIV Brazilian Conference of Meteorology, Florianópolis, Brazil, 27 November–1 December 2006. [Google Scholar]
- Steeves, T.A.; Sussex, I.M. Patterns in Plant Development, 2nd ed.; Cambridge University Press: New York, NY, USA, 1989; 408p. [Google Scholar]
- McMaster, G.S. Phytomers, phyllochrons, phenology and temperate cereal development. J. Agric. Sci. 2005, 143, 137–150. [Google Scholar] [CrossRef]
- Brossinger, G. Segments (Phytomers). In Handbook of Plant Science; Roberts, K., Ed.; Willey & Sons: Chichester, UK, 2007; pp. 200–202. [Google Scholar]
- Barbosa Machado Neto, N.; Prioli, M.R.; Gatti, A.B.; Mendes Cardoso, V.J. Temperature effects on seed germination in races of common beans (Phaseolus vulgaris L.). Acta Scient. Agron. 2006, 28, 155–164. [Google Scholar]
- Abramoff, M.D.; Magalhaes, P.J.; Ram, S.J. Image processing with ImageJ. Biophotonics Intern. 2004, 11, 36–42. [Google Scholar]
- Buck-Sorlin, G.H. L-system model of the vegetative growth of winter barley. In Proceedings of the Fifth German Workshop on Artificial Life, Lübek, Germany, 18–20 March 2002; Polani, D., Kim, J., Martinez, T., Eds.; pp. 53–64. [Google Scholar]
- R Development and Core Team. An Introduction to R. Notes on R: A Programming Environment for Data Analysis and Graphics Version 3.1.1; R Foundation for Statistical Computing: Vienna, Austria, 2014; 105p. [Google Scholar]
- Quinn, G.; Keough, M. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002; 553p. [Google Scholar]
- Carvalho, I.R.; Korcelski, C.; Pelissari, G.; Hanus, A.D.; Rosa, G.M. Water demand of crop agronomic interest. Enciclop. Biosfera 2013, 9, 969–985. [Google Scholar]
- Bergamaschi, G.; Vieira, H.J.; Libardi, P.L.; Ometto, J.C.; Angelocci, L.R. Water deficit in common bean. III. Maximum crop evapotranspiration and its relationship with evapotranspiration by the Penman method and “Class A” Pan evaporation. Pesq. Agropec. Bras. 1989, 24, 387–392. [Google Scholar]
- Santos, R.Z.; André, R.G.B. Water use for bean crop in different growth stages. Pesq. Agropec. Bras. 1992, 27, 543–548. [Google Scholar]
- Saxena, C.M.; Singh, B.K. Effect of supplementary irrigation during reproductive growth on winter and spring chickpea (Cicer arietinum) in a Mediterranean environment. J. Agric. Sci. 1990, 114, 285–293. [Google Scholar] [CrossRef]
- Acosta-Gallegos, J.A.; Shibata, J.K. Effect of water stress on growth and yield of indeterminate dry-bean (Phaseolus vulgaris) cultivars. Field Crops Res. 1989, 20, 81–93. [Google Scholar] [CrossRef]
- Castellanos, J.Z.; Pena-Cabriales, J.J.; Acosta-Gallegos, J.A. 15N-determined dinitrogen fixation capacity of common bean (Phaseolus vulgaris L.) cultivars under water stress. J. Agric. Sci. 1996, 126, 327–333. [Google Scholar] [CrossRef]
- Lopez, F.B.; Johansen, C.; Chauhan, Y.S. Effect of timing of drought stress on phenology, yield and yield components of a short-duration pigeon pea. J. Agron. Crop Sci. 1996, 177, 311–320. [Google Scholar] [CrossRef]
- Board, J.E.; Harville, B.G. Late-planted soybean yield response to reproductive source/sink stress. Crop Sci. 1998, 38, 763–771. [Google Scholar] [CrossRef]
- Linkemer, G.; Board, J.E.; Musgrave, M.E. Water logging effects on growth and yield components in late planted soybeans. Crop Sci. 1998, 38, 1576–1584. [Google Scholar] [CrossRef] [PubMed]
- Scully, B.T.; Wallace, D.H. Variation in and relationship of biomass, growth rate, harvest index, and phenology to yield of common bean. J. Am. Soc. Hortic. Sci. 1990, 115, 218–225. [Google Scholar] [CrossRef]
- Araújo, A.P.; Teixeira, M.G. Ontogenetic variations on absorption and utilization of phosphorus in common bean cultivars under biological nitrogen fixation. Plant Soil 2000, 225, 1–10. [Google Scholar] [CrossRef]
- Foster, E.F.; Pajarito, A.J.A.; Acosta, G. Moisture stress impact on N partitioning, N remobilization and N-use efficiency in beans (Phaseolus vulgaris L.). J. Agric. Sci. 1995, 124, 27–37. [Google Scholar] [CrossRef]
- Acosta-Díaz, E.; Acosta-Gallegos, J.A.; Trejo-López, C.; Padilla-Ramírez, J.S.; Amador-Ramírez, M.D. Adaptation traits in dry bean cultivars grown under drought stress. Agric. Téc. Méx. 2009, 4, 416–425. [Google Scholar]
- Gholinezhad, E.; Aynaband, A.; Ghorthapeh, A.H.; Noormohamadi, G.; Bernousi, I. Study of the effect of drought stress on grain yield, quality traits, phyllochron and leaf appearance rate of sunflower hybrid Iroflor at different levels of nitrogen and plant population. Am.-Eurasian J. Agric. 2012, 12, 306–314. [Google Scholar]
- Albert, D.G.; Carberry, P.S. The influence of water deficit prior to tassel initiation on maize growth, development and yield. Field Crops Res. 1993, 31, 55–59. [Google Scholar] [CrossRef]
- White, J.W.; Izquierdo, J. Physiology of yield potential and stress tolerance. In Common Beans: Research for Crop Improvement; Schoonhoven, A., Voysest, O., Eds.; C. A. B. International: Wallingford, UK, 1991; pp. 287–382. [Google Scholar]
- Robins, J.S.; Domingo, C.E. Moisture deficits in relation to the growth and development of dry beans. Agron. J. 1956, 48, 67–70. [Google Scholar] [CrossRef]
- Kohashi-Shibata, J.; Galván, M.T.; García, A.E.; Yánez, P.J.; Martínez, E.V.; Ruiz, L.P. Water stress effect on the growth of phytomers in common bean (Phaseolus vulgaris L.). Agric. Téc. Méx. 2002, 28, 65–75. [Google Scholar]
- Ku, Y.S.; Au-Yeung, W.K.; Yung, Y.L.; Li, M.W.; Wen, C.Q.; Liu, X.; Lam, H.M. Drought stress and tolerance in soybean. In A Comprehensive Survey of International Soybean Research—Genetics, Physiology, Agronomy and Nitrogen Relationships; Board, J.E., Ed.; InTech: Rijeka, Croatia, 2013; pp. 209–237. [Google Scholar]
- Du, N.; Guo, W.; Zhang, X. Morphological and physiological responses of Vitex negundo L. var. heterophylla (Franch.) Rehd. to drought stress. Acta Physiol. Plant. 2010, 32, 839–848. [Google Scholar] [CrossRef]
- Oosterhuis, D.M.; Walker, S.; Eastham, J. Soybean leaflet movements as an indicator of crop water stress. Crop Sci. 1985, 25, 1101–1106. [Google Scholar] [CrossRef]
- Yu, F.; Berg, V.A. Control of paraheliotropism in two Phaseolus species. Plant Physiol. 1994, 106, 1567–1573. [Google Scholar] [CrossRef]
- Pastenes, C.; Pimentel, P.; Lillo, J. Leaf movements and photoinhibition in relation to water stress in field-grown beans. J. Exp. Bot. 2004, 56, 425–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, B.; Sathitsuksanoh, N.; Tang, Y.H.; Udvardi, M.K.; Zhang, J.Y.; Shen, Z.X.; Balota, M.; Harich, K.; Zhang, Y.H.P.; Zhao, B.Y. Overexpression of AtLov1 in switchgrass alters plant architecture, lignin content, and flowering time. PLoS ONE 2012, 7, e47399. [Google Scholar] [CrossRef] [PubMed]
- Chávez, R.O.; Clevers, J.G.P.W.; Herold, M.; Ortiz, M.; Acevedo, E. Modelling the spectral response of the desert tree Prosopis tamarugo to water stress. Int. J. Appl. Earth Obs. Geoinf. 2013, 21, 53–65. [Google Scholar] [CrossRef]
- Farooq, M.; Gogoi, N.; Barthakur, S.; Baroowa, B.; Bharadwaj, N.; Alghamdi, S.S.; Siddique, K.H.M. Drought stress in grain legumes during reproduction and grain filling. J. Agron. Crop Sci. 2017, 203, 81–102. [Google Scholar] [CrossRef]
- Desclaux, D.; Huynh, T.T.; Roumet, P. Identification of soybean plant characteristics that indicate the timing of drought stress. Crop Sci. 2000, 40, 716–722. [Google Scholar] [CrossRef]
- Ueda, A.; Kathiresan, A.; Inada, M.; Narita, Y.; Nakamura, T.; Shi, W.; Takabe, T.; Benett, J. Osmotic stress in barley regulates expression of a different set of genes than salt stress does. J. Exp. Bot. 2009, 55, 2213–2218. [Google Scholar] [CrossRef] [PubMed]
- Vos, J.; Evers, J.B.; Buck-Sorlin, G.H.; Andrieu, B.; Chelle, M.; de Visser, P.H.B. Functional-structural plant modeling: A new versatile tool in crop science. J. Exp. Bot. 2010, 61, 2101–2115. [Google Scholar] [CrossRef] [PubMed]
- Evers, J.B. Simulating crop growth and development using functional-structural plant modeling. In Canopy Photosynthesis: From Basic to Applications; Hikosaka, K., Niinemets, U., Anten, N.P.R., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 219–236. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Treatment | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | FL | FH | PL | PH | ||||||
| Water applied (mm) | 383 | 308 | 259 | 316 | 272 | |||||
| Evapotranspiration (mm d−1) | 4.89 ± 0.22 1 | a | 3.99 ± 0.12 | b | 3.33 ± 0.11 | c | 4.11 ± 0.14 | b | 3.54 ± 0.13 | c |
| Grain yield (g plant−1) | 12.89 ± 0.34 | a | 9.31 ± 0.25 | c | 7.16 ± 0.31 | d | 11.09 ± 0.31 | b | 8.16 ± 0.40 | cd |
| Shoot biomass (g plant−1) | 24.95 ± 1.23 | a | 16.84 ± 0.80 | c | 13.18 ± 0.67 | d | 19.42 ± 1.17 | b | 15.52 ± 0.47 | cd |
| Harvest index (HI) | 0.52 ± 0.03 | a | 0.56 ± 0.03 | a | 0.55 ± 0.03 | a | 0.58 ± 0.04 | a | 0.53 ± 0.04 | a |
| Number of pods per plant | 39.41 ± 1.23 | a | 41.24 ± 1.81 | a | 33.04 ± 2.97 | a | 37.49 ± 2.03 | a | 40.87 ± 1.56 | a |
| Length of pods (10−2 m) | 9.86 ± 1.91 | a | 8.12 ± 2.18 | b | 7.74 ± 2.26 | c | 8.98 ± 2.14 | b | 7.35 ± 2.21 | c |
| Phyllochron on branch 2.1 (°Cd) | 67.91 ± 4.15 | a | 91.81 ± 1.23 | b | 91.44 ± 1.02 | b | 68.94 ± 1.22 | a | 71.91 ± 1.43 | a |
| Phyllochron on branch 2.2 (°Cd) | 66.77 ± 6.56 | a | 91.86 ± 0.85 | b | 91.34 ± 1.25 | b | 76.62 ± 3.64 | a | 76.37 ± 4.16 | a |
| Final number of phytomers | 21.03 ± 1.08 | a | 17.84 ± 1.25 | ab | 14.21 ± 1.66 | b | 20.49 ± 1.07 | a | 21.45 ± 0.79 | a |
| Final number of leaflet blades | 55.03 ± 3.71 | a | 44.44 ± 4.85 | ab | 33.21 ± 2.82 | b | 52.81 ± 4.67 | a | 52.08 ± 1.23 | a |
| Angles—petiolules and top leaflet blades (°) | 135.22 ± 1.72 | a | 134.07 ± 2.86 | a | 120.89 ± 4.01 | b | 130.63 ± 2.29 | a | 126.62 ± 1.72 | b |
| Variable | Fitted Coefficients | Treatment | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | FL | FH | PL | PH | |||||||
| Length of internodes | Lm (10−2 m) | 6.71 ± 0.14 1 | bc | 7.35 ± 0.22 | ab | 6.47 ± 0.18 | c | 7.40 ± 0.18 | a | 7.21 ± 0.17 | abc |
| xo | 6.35 ± 0.04 | ||||||||||
| α | 1.95 ± 0.07 | ||||||||||
| Length of petioles | a (10−3 m) | −4.91 ± 0.05 | a | −5.24 ± 0.07 | b | −5.39 ± 0.09 | b | −5.14 ± 0.06 | ab | −5.28 ± 0.07 | b |
| b (10−2 m) | 5.50 ± 0.30 | ||||||||||
| c (10−2 m) | −6.63 ± 0.7 | ||||||||||
| Length of petiolules | a (10−3 m) | −1.64 ± 0.02 | a | −1.76 ± 0.02 | b | −1.76 ± 0.03 | b | −1.70 ± 0.02 | ab | −1.78 ± 0.02 | b |
| b (10−2 m) | 1.91 ± 0.18 | ||||||||||
| c (10−2 m) | −3.20 ± 0.49 | ||||||||||
| Length of leaflet blades (main stems) | a (10−3 m) | −3.41 ± 0.20 | a | −5.01 ± 0.37 | b | −4.18 ± 0.33 | ab | −4.26 ± 030 | ab | −3.30 ± 0.41 | a |
| b (10−2 m) | 3.37 ± 0.20 | b | 4.50 ± 0.34 | a | 3.55 ± 0.31 | ab | 3.81 ± 030 | ab | 3.03 ± 0.39 | b | |
| c (10−2 m) | 1.98 ± 0.47 | ab | −0.05 ± 0.73 | b | 2.41 ± 0.69 | ab | 2.34 ± 0.68 | ab | 3.32 ± 0.85 | a | |
| Length of leaflet blades (branches) | a (10−3 m) | −4.74 ± 0.35 | a | −5.66 ± 0.43 | ab | −5.20 ± 1.00 | ab | −4.48 ± 0.41 | a | −6.19 ± 0.38 | b |
| b (10−2 m) | 4.35 ± 0.32 | ab | 4.70 ± 0.37 | ab | 4.15 ± 0.76 | ab | 3.75 ± 0.38 | b | 5.49 ± 0.33 | a | |
| c (10−2 m) | −2.64 ± 0.73 | a | −3.22 ± 0.77 | ab | −2.50 ± 1.37 | ab | −1.42 ± 0.83 | a | −5.62 ± 0.68 | b | |
| Width of leaflet blades (main stems) | a (10−3 m) | −2.36 ± 0.13 | ab | −3.49 ± 0.21 | b | −2.72 ± 0.21 | ab | −2.88 ± 0.19 | b | −1.90 ± 0.27 | a |
| b (10−2 m) | 2.21 ± 0.13 | b | 3.02 ± 0.19 | a | 2.19 ± 0.21 | b | 2.51 ± 0.19 | ab | 1.67 ± 0.26 | b | |
| c (10−2 m) | 1.62 ± 0.31 | ab | 0.39 ± 0.41 | b | 2.30 ± 0.45 | a | 1.66 ± 0.43 | ab | 3.22 ± 0.57 | a | |
| Width of leaflet blades (branches) | a (10−3 m) | −3.03 ± 0.26 | a | −4.12 ± 0.32 | ab | −4.16 ± 0.72 | ab | −3.17 ± 0.30 | ab | −4.25 ± 0.29 | b |
| b (10−2 m) | 2.71 ± 0.24 | b | 3.38 ± 0.28 | ab | 3.17 ± 0.55 | ab | 2.59 ± 0.28 | b | 3.73 ± 0.26 | a | |
| c (10−2 m) | −1.83 ± 0.55 | a | −3.02 ± 0.58 | ab | −2.53 ± 0.99 | ab | −1.50 ± 0.61 | a | −4.29 ± 0.52 | b | |
| Angles—first internodes and internodes of the main originating stems | d (°) | −1.32 ± 0.22 | a | −2.49 ± 0.35 | a | −2.50 ± 0.35 | a | −1.36 ± 0.29 | a | −1.70 ± 0.27 | a |
| e (°) | 15.95 ± 1.15 | b | 21.58 ± 1.53 | a | 21.67 ± 1.45 | a | 16.73 ± 1.41 | ab | 18.23 ± 1.25 | ab | |
| f (°) | 116.90±2.83 | ||||||||||
| Variable | Treatment | ||||
|---|---|---|---|---|---|
| Control | FL | FH | PL | PH | |
| Coefficient of Determination (r2) | |||||
| Length of internodes | 0.64 | 0.51 | 0.61 | 0.61 | 0.56 |
| Length of petioles | 0.55 | 0.51 | 0.38 | 0.36 | 0.36 |
| Length of petiolules | 0.28 | 0.21 | 0.13 | 0.16 | 0.31 |
| Length of leaflet blades (main stems) | 0.61 | 0.54 | 0.59 | 0.58 | 0.28 |
| Length of leaflet blades (branches) | 0.29 | 0.33 | 0.11 | 0.24 | 0.38 |
| Width of leaflet blades (main stems) | 0.67 | 0.67 | 0.65 | 0.63 | 0.25 |
| Width of leaflet blades (branches) | 0.23 | 0.32 | 0.11 | 0.24 | 0.33 |
| Angles—first internodes and internodes of the main originating stems | 0.59 | 0.41 | 0.63 | 0.61 | 0.62 |
| Root mean squared error (RMSE) | |||||
| Length of internodes (10−2 m) | 1.3 | 1.8 | 1.4 | 1.6 | 1.6 |
| Length of petioles (10−2 m) | 1.8 | 1.9 | 2.3 | 2.2 | 2.3 |
| Length of petiolules (10−2 m) | 0.6 | 0.6 | 0.8 | 0.8 | 0.7 |
| Length of leaflet blades (main stems) (10−2 m) | 0.9 | 1.1 | 1.2 | 1.2 | 1.4 |
| Length of leaflet blades (branches) (10−2 m) | 1.6 | 1.5 | 1.8 | 1.8 | 1.7 |
| Width of leaflet blades (main stems) (10−2 m) | 0.8 | 0.6 | 0.8 | 0.8 | 1.0 |
| Width of leaflet blades (branches) (10−2 m) | 1.2 | 1.2 | 1.3 | 1.4 | 1.3 |
| Angles—first internodes and internodes of the main originating stems (°) | 22.35 | 30.37 | 26.93 | 20.05 | 22.34 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Durigon, A.; Evers, J.; Metselaar, K.; de Jong van Lier, Q. Water Stress Permanently Alters Shoot Architecture in Common Bean Plants. Agronomy 2019, 9, 160. https://doi.org/10.3390/agronomy9030160
Durigon A, Evers J, Metselaar K, de Jong van Lier Q. Water Stress Permanently Alters Shoot Architecture in Common Bean Plants. Agronomy. 2019; 9(3):160. https://doi.org/10.3390/agronomy9030160
Chicago/Turabian StyleDurigon, Angelica, Jochem Evers, Klaas Metselaar, and Quirijn de Jong van Lier. 2019. "Water Stress Permanently Alters Shoot Architecture in Common Bean Plants" Agronomy 9, no. 3: 160. https://doi.org/10.3390/agronomy9030160
APA StyleDurigon, A., Evers, J., Metselaar, K., & de Jong van Lier, Q. (2019). Water Stress Permanently Alters Shoot Architecture in Common Bean Plants. Agronomy, 9(3), 160. https://doi.org/10.3390/agronomy9030160