1. Introduction
Maize (
Zea mays L.) is an important global cereal crop, and its greater production is required to meet growing food demands of the ever-increasing world population [
1]. Globally, increasing planting density and nitrogen fertilizer application are very effective agronomic strategies to raise maize yield, but high density and excessive or inappropriate nitrogen application lead to an unreasonable canopy and weak stalks, which often increase lodging risk [
2,
3]. Generally, maize lodging occurs at both roots and stalks, and root lodging is caused by the failure of root anchorage which is affected by soil properties, root characteristics and pests [
4,
5]. Stalk lodging occurs most frequently at the basal internode above ground and bending or breaking mainly occurs at the 3rd to 5th basal elongation internodes [
6,
7,
8,
9]. Between 5 and 25% of annual maize yield loss is due to stalk lodging [
10]. Numerous studies have demonstrated that stalk strength is a reliable predictor of lodging risk [
11,
12,
13], and stalk strength is also one of the important agronomic traits linked to maize yield potential [
14]. Accordingly, improving maize stalk strength—especially the basal internodes bending resistance—is critical for reducing lodging risk and advancing maize production [
7,
9].
It has been proven that stalk bending strength is closely related to the geometrical and material properties of the stalks in both cereals [
3,
15,
16,
17,
18] and dicotyledonous crops [
19,
20]. For instance, lodging resistance is closely associated with basal internode length and stem diameter in barley and wheat [
21,
22,
23,
24,
25]. In maize, stalk lodging positively correlates with plant height, ear height, number of internodes under the ear and length of basal internodes [
26,
27], but negatively correlates with basal internode diameter [
6,
8]. Meanwhile, stalk strength has a significantly positive relationship with the amount or distribution of structural chemical materials in maize internode [
28,
29]. Likewise, plants with lower lignin content display an insufficient level of fitness to increase stalk lodging risk in various crops [
30]. Plants have high stem rigidity and lodging resistance through increasing lignin and cellulose content in rice internodes [
18,
31]. Moreover, the size of the secondary structure in the stem presents a strong relationship with stem lodging in barley (
Hordeum vulgare L.) [
21], wheat (
Triticum aestivum L.) [
23] and sunflower (
Helianthus annuus L.) [
20]. Similarly, Zuber et al. [
32] reported that about 50 to 80% of the strength of maize stalk comes from the rind. In addition, 79% of the penetration resistance strength in maize basal internodes depends on the number of mechanical cell layers, the thickness of mechanical tissue and the ratio of cortex to internode cross-section [
9].
Although the contribution of stem morphological and chemical traits to stem strength is disputable, it is generally recognized that stem strength is improved by genetic and agronomic strategies for modulating stem morphological and chemical traits in various plants. Based on the genetic diversity for stalk strength characteristics, the selection and breeding of lodging resistant varieties are very important and effective strategies to decrease the risk of lodging in crops [
13,
23,
25,
33]. For example, the selection and application of semi-dwarf traits have increased crop lodging resistance under higher plant density and fertilizer conditions, which contributed to the improvement of crop yield during the “Green Revolution” [
34]. Meanwhile, agronomic management practices including planting density, irrigation and fertilization can regulate the morphological and chemical traits of maize stalk affecting stalk strength [
27,
35,
36,
37]. Furthermore, plant growth regulators have been demonstrated as an effective strategy to reduce the risk of lodging by modulating stalk morphological and chemical characteristics in crops [
3,
38].
Ethephon (2-chloroethy-phosphonic acid) as an ethylene-releasing compound is widely used for reducing plant height to decrease the risk of stalk lodging in cereal production including maize [
39,
40], wheat [
38,
41] and barley [
42]. Mangieri et al. [
20] reported that ethephon increases the thickness of primary and secondary structures in sunflower stems for improving lodging resistance. In addition, lignification of the xylem cell wall is enhanced by ethephon in the
Pinus radiata D. Don stem [
43]. Ethephon also regulates the deposition of cellulose and hemicelluloses in the basal internodes of maize [
27]. Moreover, ethephon enhances the lodging resistance of maize stalk by increasing crushing strength and breaking force [
27,
44]. However, there is a weak relationship between lodging resistance and the cellulose and hemicellulose concentrations in ethephon-treated maize stalk [
27]. Recent simulation studies showed that maize stalk bending strength is highly dependent on stalk morphology [
45,
46,
47]. Above all, ethephon can regulate stalk strength involving the modulation of morphological and chemical traits, but it is still unclear how ethephon affects the mechanical characteristics of maize internode, and how ethephon-regulated internode morphological and chemical properties contribute to stalk strength. Furthermore, inappropriate application of ethephon often reduces maize grain yield [
39,
40,
48]. Therefore, it is essential to explore the roles of ethephon in regulating the morphological and chemical traits of maize stalk to enhance stalk strength for better application in maize production.
The objective of the present study was to investigate the effects of ethephon on maize internode morphological, chemical and mechanical characteristics based on multiyear field experiments, and to further analyze the contribution of ethephon-modulating internode morphological and chemical traits to mechanical functionality for stalk strength. This research would be beneficial for evaluating the role of ethephon in improving maize stalk strength, which could promote efficient ethephon application in maize production.
4. Discussion
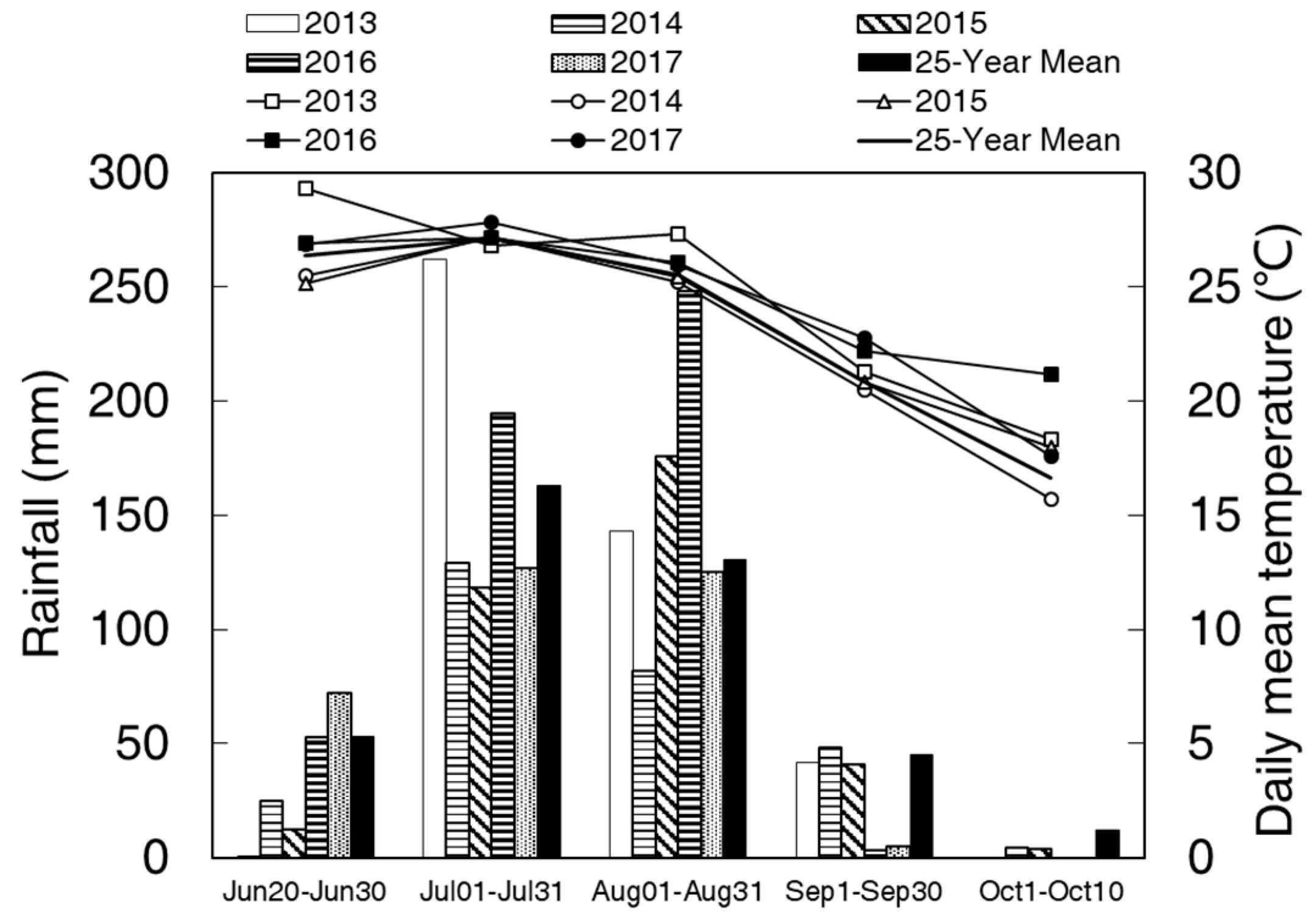
The North China Plain (NCP) produces almost one-third of maize output in China, and summer maize is the predominant maize planting model in this region. However, a typical temperate monsoon climate in NCP, with the summer maize growing in a high temperature and humid environment, often gives rise to keeping a high growth rate, while local farmers usually apply superfluous nitrogenous fertilizer and higher plant densities than is optimum in practice, which causes luxuriant growth leading to excessive plant height and weak mechanical features increasing the stalk’s susceptibility to lodging [
2,
54]. Moreover, stalk lodging is associated with morphological traits as well as environmental conditions and breaking usually arises from the mechanical failure of stalk tissue below the ear node before harvest [
55]. Based on multiyear (2013−2017) field experiments, ethephon could significantly decrease plant and ear heights and basal internode length but increase the maximum and minimum diameter of basal internodes (
Table 1), which leads to higher breaking resistance and bending strength of the internode compared to control (
Table 1). Similarly, numerous studies revealed that ethephon remarkably increases crops’ lodging resistance by reducing plant height and basal internode length while increasing internode diameters and structural dry matter deposition in the internode [
27,
39,
40,
41,
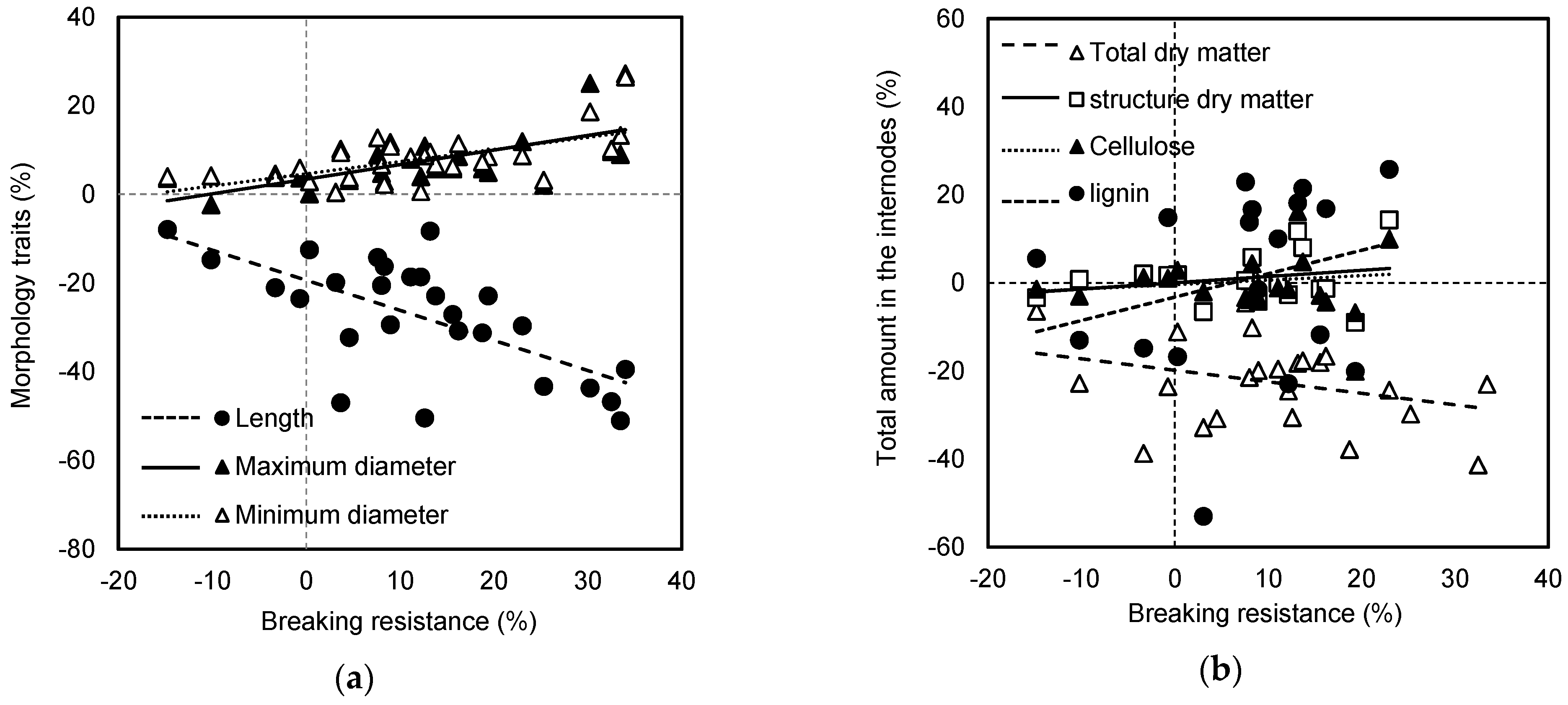
42]. However, internode breaking resistance and morphological parameters were also significantly affected by years, which led to weak correlations between internode breaking resistance and morphological parameters (
Figure S1). For highlighting the roles of ethephon in internode properties regulation and eliminating weather effects, the relative change ratio of each parameter ((ET-Ct)/Ct) was used for further analysis. As expected, a strong correlation was observed between internode breaking resistance and internode length and diameters. These results indicated that the stable effects of ethephon altered internode properties was the reason for improved stalk strength among different growing seasons. Similar effects of the ethephon application on increasing crop stalk strength and lodging resistance are also approved as relatively stable in various species [
20,
38,
41,
42]. Thus, the modification of internode morphology played an important role in ethephon’s improvement of the stalk strength of maize.
In general, the strength of any structure rests with both material and morphology [
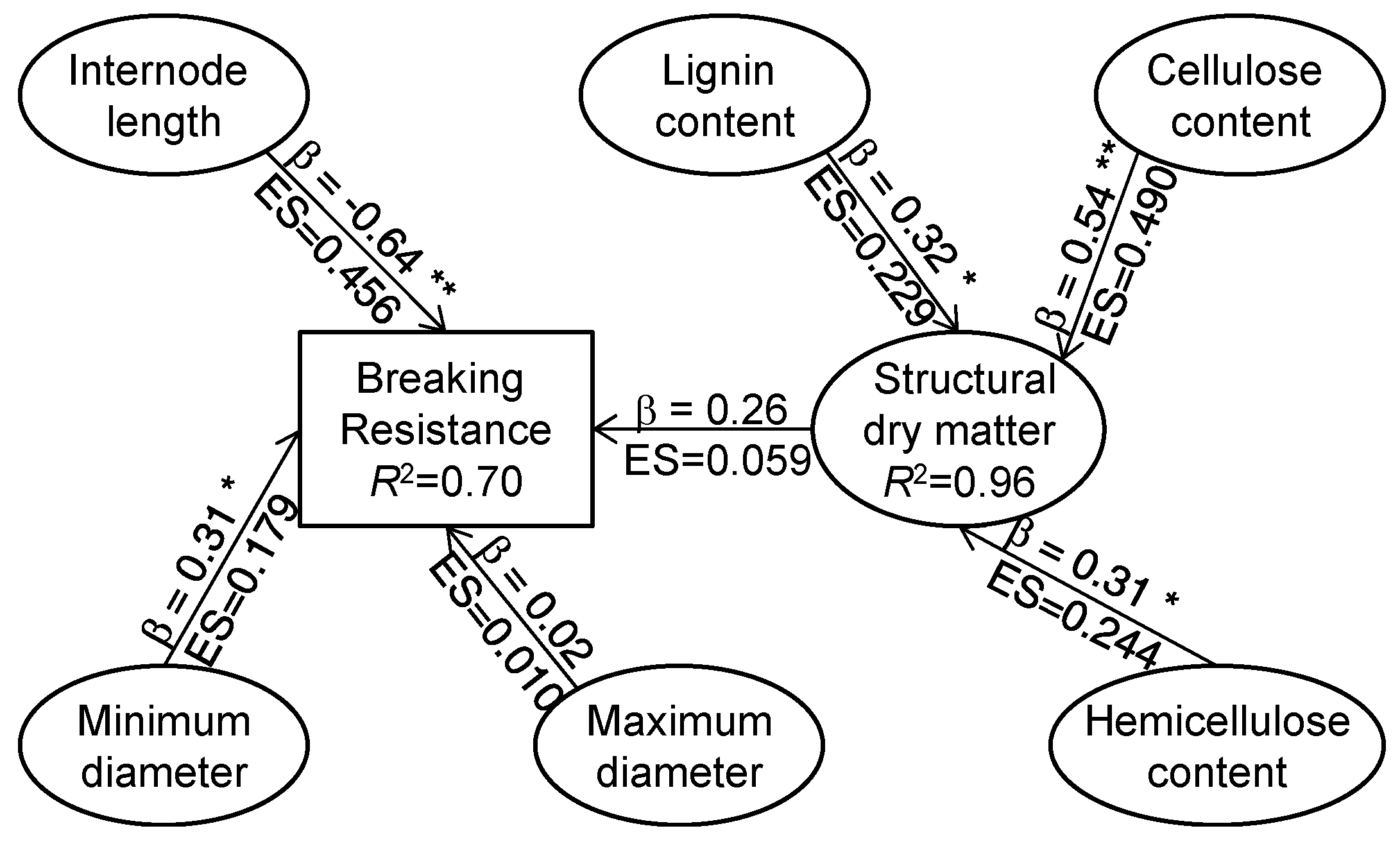
3]. Several studies suggested that the size of the secondary structure and the content of structural dry matter may be important factors for improving stalk breaking resistance in maize [
28,
29], wheat [
23], barley [
21], sunflower [
20] and rice [
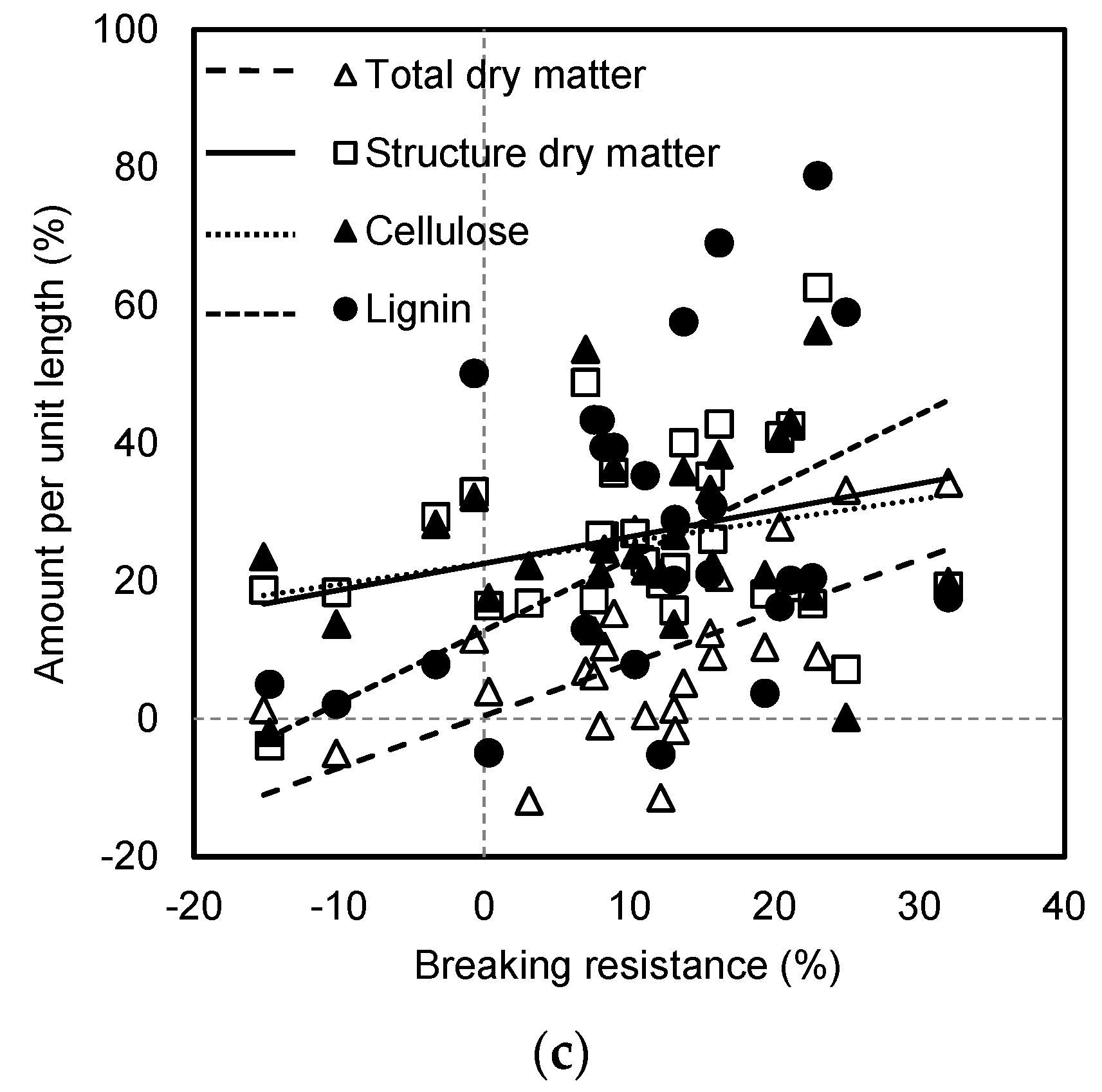
31]. In this study, weak positive correlations were observed between internode breaking resistance and the total dry matter and structural dry matter amount in the internode (
Figure S2). Similarly, Kong et al. [
56] suggested that there is a weak relationship between lodging resistance and cellulose and hemicellulose amount in the whole stalk. Based on the morphological traits, many researchers define the total weight per unit length as stalk density for predicting stalk strength [
28,
56,
57]. Here, there were strong correlations between the breaking resistance and amount per unit length or volume of total dry matter and structural dry matter (
Figure S2). Similar results were observed by Appenzeller et al. [
28], who suggested that dry matter per unit length explained more than 50% variation in mechanical strength.
Ethylene can enhance lignin accumulation by regulating the lignin biosynthesis enzymes, such as phenylalanine ammonialyase, cinnamylalcohol dehydrogenase and peroxidase in bamboo shoot [
58], transgenic tobacco plant [
59] and mungbean roots [
60]. Meanwhile, ethephon can improve breaking resistance by increasing the thickness of secondary structure and secondary tissue area in sunflowers [
20], and increasing the deposition or allocation of hemicellulose, cellulose and lignin in maize [
27]. In this study, ethephon significantly increased the ratio of structural dry matter to total dry matter in the internode and enhanced the amount per unit length and unit volume of structural dry matter including hemicellulose, cellulose and lignin (
Table 3 and
Table S2). Furthermore, the correlation coefficient between relative change ratio of internode morphological properties and breaking resistance were higher than the coefficient between the relative change ratio of breaking resistance and internode chemical properties (
Figure 3 and
Table 5). The path analysis also revealed that the alteration of morphological traits by ethephon contributed almost twice as much variation in internode breaking resistance as the alteration of chemical composition (
Figure 4). These results indicated that ethephon enhanced stalk strength mainly by altering the morphological properties of basal internodes, and partly through regulating the deposition of structural dry matter. Our results also indicated that controlling the plant type could be more effective for decreasing crop lodging.
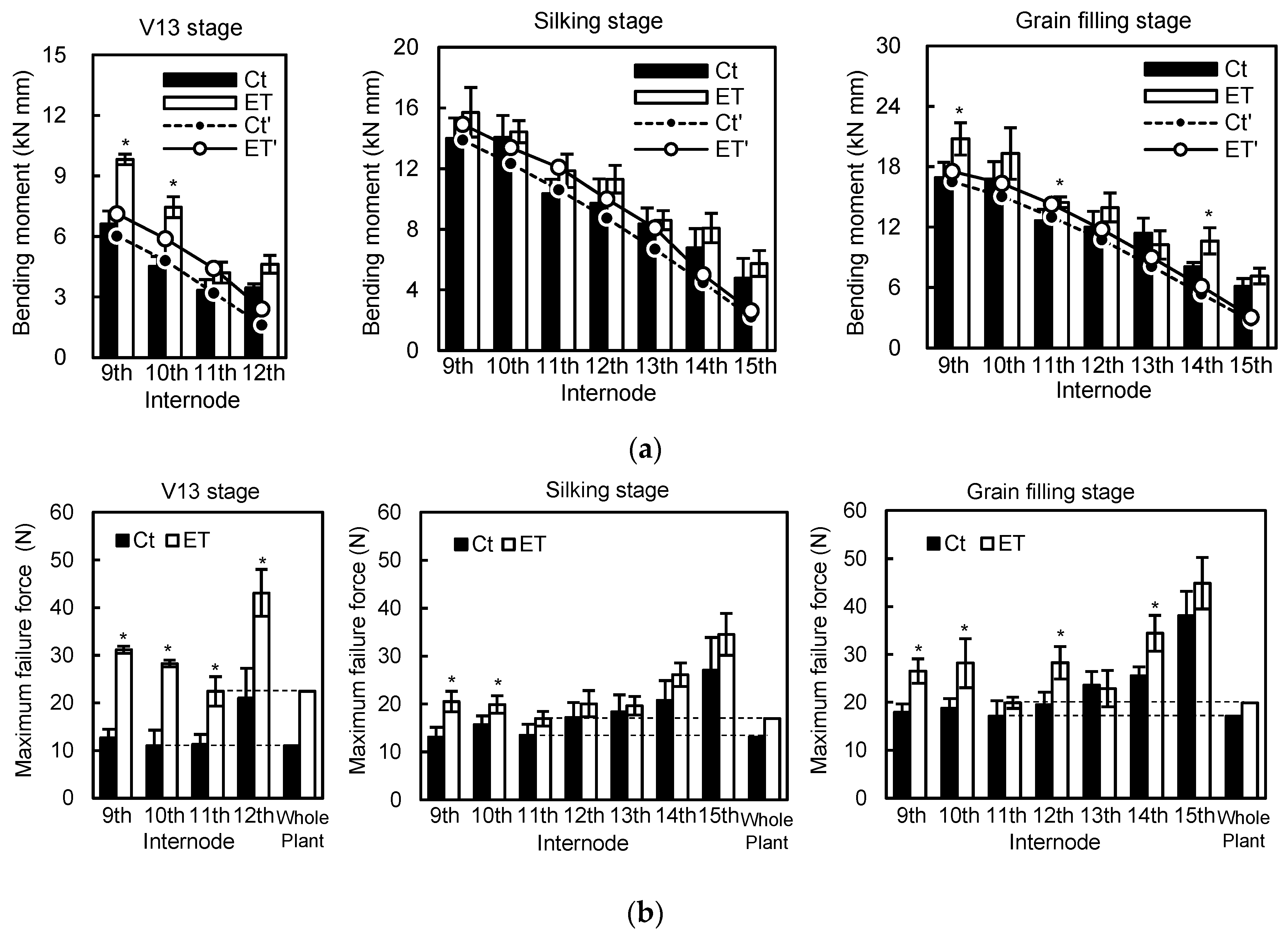
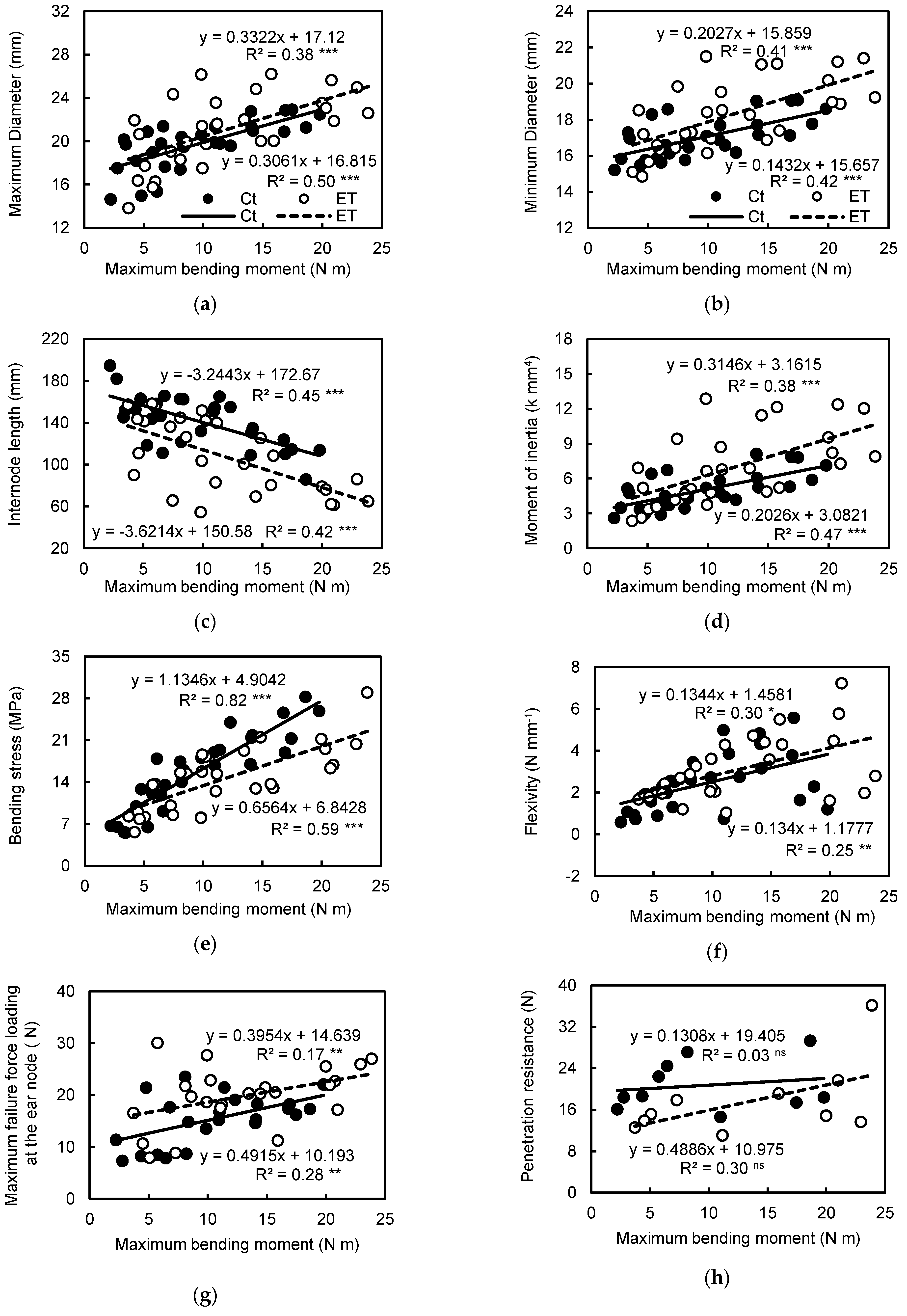
Mechanical parameters are approved and accepted as stalk strength predictors and the bending test has been widely used in plant biomechanics [
3,
24,
46,
61]. Although maize stalk sustains both bending and shear-compressive force under field condition, the bending stresses are many orders of magnitude greater than shear or compressional stresses [
45]. Meanwhile, the bending test estimates not only the bending mechanical properties but also material properties including elasticity modulus (Young’s modulus), flexural stiffness (predicted by flexibility) by using data from bending experiments [
11,
12,
25,
62]. In the present study, ethephon could significantly increase the maximum bending moment, moment of inertia, maximum failure force, flexibility of internode and the maximum failure force when loading at the ear, but decrease the maximum bending stress, Young’s modulus and penetration resistance (
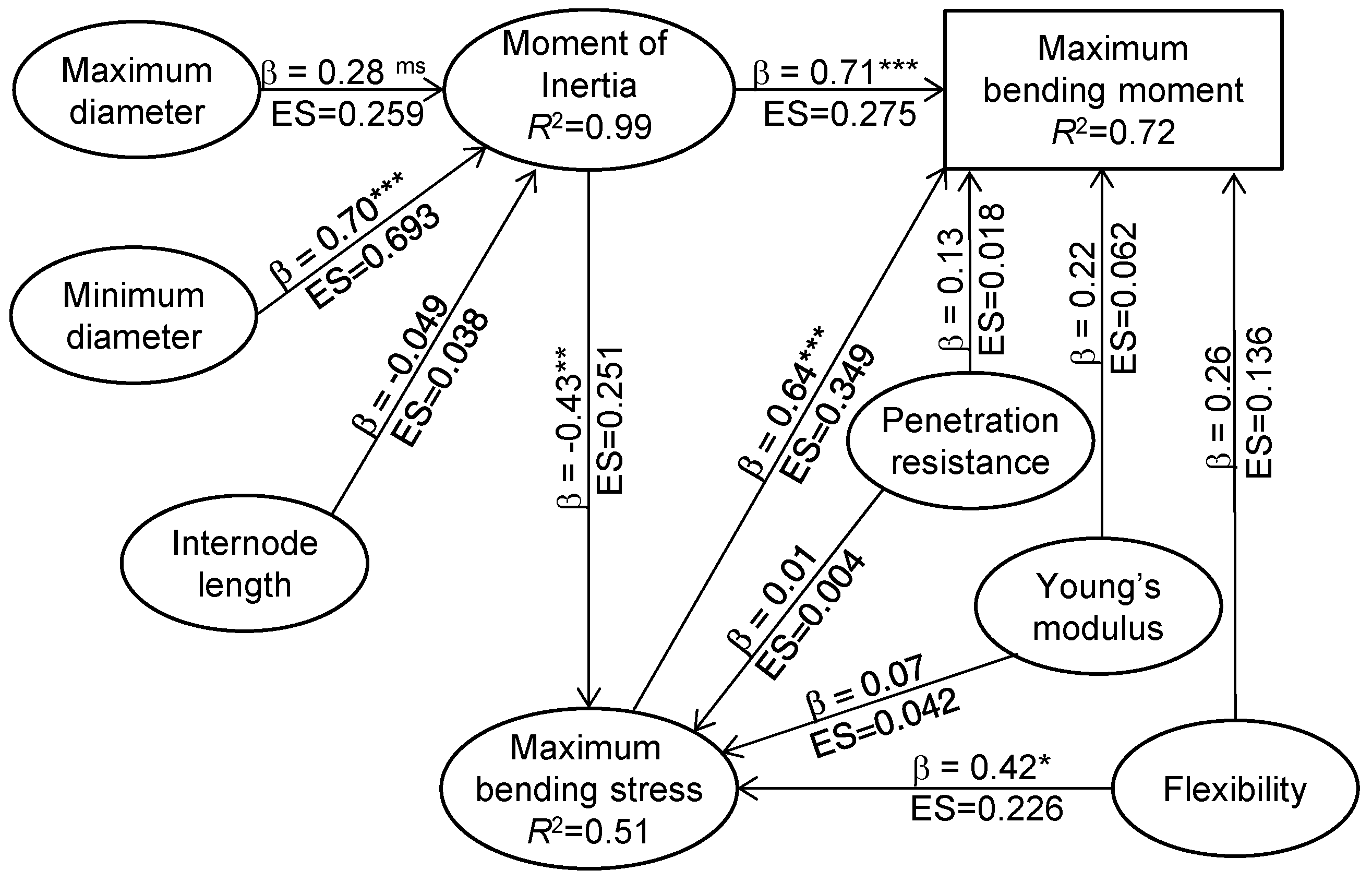
Table 3). Generally, the maximum bending moment governs the internode bending failure as all structures and is usually defined as stalk bending strength [
61]. Here, the maximum bending moment had a strong positive correlation with bending stress, and medium strong correlation with flexibility and geometric parameters including internode diameter and length, while having a weak correlation with Young’s modulus and penetration resistance (
Figure 4). Path coefficient analysis suggested that ethephon-improved maximum bending moment was mainly attributed to the change of maximum bending stress and the moment of inertia. There were some indirect effects of moment of inertia on maximum bending stress in the effect size of maximum bending stress on maximum bending strength (
Figure 6). Ethephon increased both maize stalk flexibility and structural dry matter, while these two properties relate with material rigidity [
18,
31]. Although alteration of material properties had a weak contribution to internode strength improvement, internode flexibility had a larger effect size than other material properties (
Figure 5). Similarly, Robertson et al. [
12] suggested that the predictive potential of internode flexibility for stalk strength has higher robustness than the predictive potential of rind penetration resistance.
In addition, maximum bending stress is defined as tissue strength, which also depends on both geometric and material parameters [
45,
47]. Cell wall lignification positively correlated with stem bending stress in rice [
31]. In this study, internode bending stress was decreased by ethephon, although flexibility of internode increased. SEM of mechanical properties revealed that negative effects of the moment of inertia on maximum bending stress contributed more than the positive effects of flexibility (
Figure 6). Ethephon-enhanced moment of inertia contributed more to bending stress than the improved flexibility, which could be the reason for the lower bending stress in ethephon-treated internode (
Table 4 and
Table 5). Von Forell et al. [
45] observed similar results, that the modulation of maize internode bending stress is much more sensitive to changes in dimensions of the stalk cross-section than changes in material properties of stalk components. Above all, ethephon-induced geometric parameter changes influenced internode bending strength greater than alteration of structure material properties.
Although ethephon dramatically improved internode strength, ethephon decreased grain yield by 3.3% compared to the control in the present case in which no lodging occurred during the 2013 to 2017 growing seasons. Similar results were observed in previous studies on maize [
39,
40,
63] and other cereal crops [
41,
42]. Nevertheless, our previous studies demonstrated that 90 g ha
-1 ethephon application before V9 stage does not significantly affect grain yield [
48]. Moreover, recent studies reported that novel plant growth regulators using ethephon as the main ingredient have positive effects on grain yield regardless of whether lodging occurs [
36,
44,
64]. Considering the potential loss of 5–25% of yield caused by lodging [
10], ethephon is still an effective way to reduce maize lodging risk under high N application and planting density conditions. Further studies should be conducted to explore the physiological and molecular mechanisms of ethephon on regulating ear and grain development for improving ethephon application in maize production.

 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}