Impact of Different Shading Levels on Growth, Yield and Quality of Potato (Solanum tuberosum L.)

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
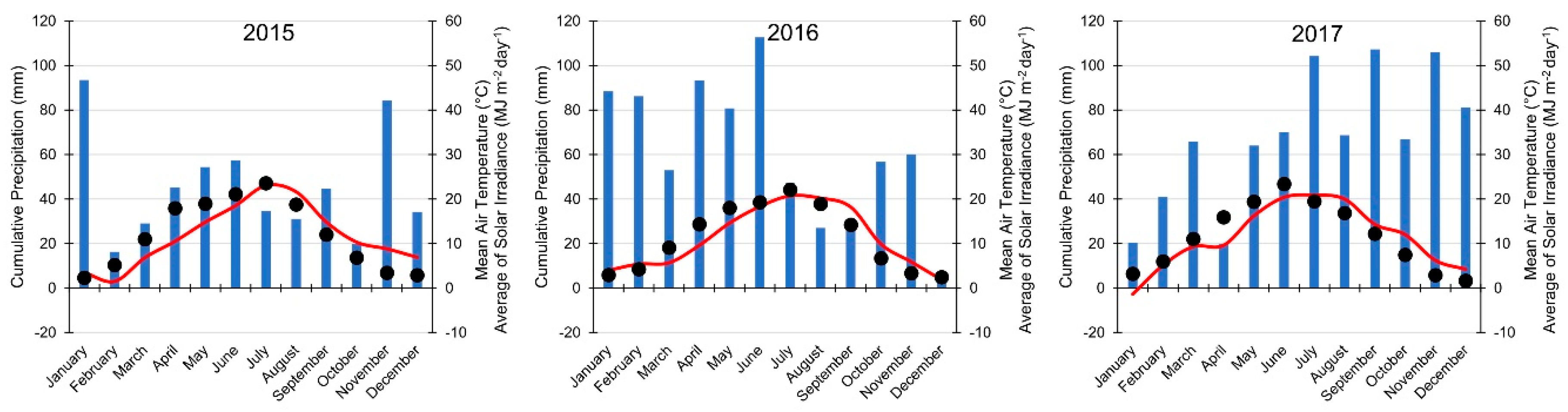
2.1. Site Conditions and Experimental Design
2.2. Shading Levels
2.3. Data Collection and Analysis
2.3.1. Growth Parameters
2.3.2. Yield Parameters
2.3.3. Quality Parameters
2.3.4. Data Analysis and Statistics
3. Results and Discussion
3.1. Growth and Development
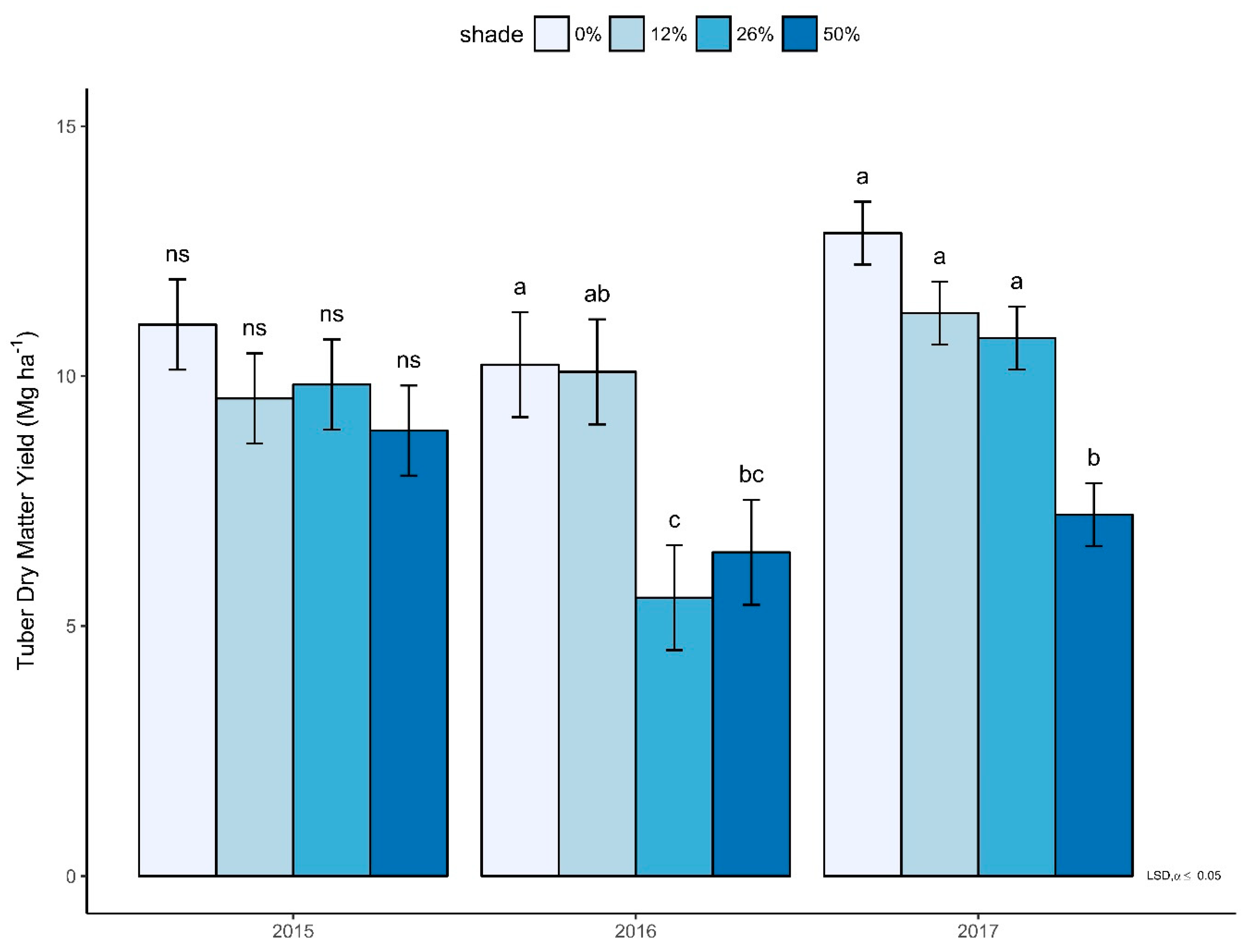
3.2. Yield Determining Parameters and Yield
3.3. Quality Parameters of Tubers
3.4. Prospects for AFS: Potential Total Solar Irradiance in the Temperate Zone of North-European Latitudes
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Product | Trade Name | Amount and Active Ingredient | MoA |
|---|---|---|---|---|
| 2015 | ||||
| 18 May | H | 2.0 kg ha−1Artist (Bayer AG) | 240 g kg−1 flufenacet, 175 g kg−1 metribuzin | K3 C1 |
| 10 June | F | 2.0 kg ha−1 Ridomil Gold (Syngenta AG) | 40 g kg−1 metalaxyl-M, 640 g kg−1 mancozeb | A1 M3 |
| 10 June | I | 0.3 L ha−1 Biscaya (Bayer AG) | 240 g L−1 thiacloprid | 4A |
| 25 June | F | 2 kg ha−1 Acrobat Plus WG (BASF SE) | 90 g kg−1 dimethomorph, 600 g kg−1 mancozeb | H5 M3 |
| 10 July | F | 2 kg ha−1 Acrobat Plus WG (BASF SE) | 90 g kg−1 dimethomorph, 600 g kg−1 mancozeb | H5 M3 |
| 10 July | I | 0.3 L ha−1 Biscaya (Bayer AG) | 240 g L−1 thiacloprid | 4A |
| 24 July | F | 2 kg ha−1 Acrobat Plus WG (BASF SE) | 90 g kg−1 dimethomorph, 600 g kg−1 mancozeb | H5 M3 |
| 6 August | F | 2 kg ha−1 Acrobat Plus WG (BASF SE) | 90 g kg−1 dimethomorph, 600 g kg−1 mancozeb | H5 M3 |
| 2016 | ||||
| 6 May | H | 3 L ha−1 Boxer (Syngenta AG) | 800 g L−1 prosulfocarb | N |
| 6 May | H | 0.3 kg ha−1 Sencor WG (Syngenta AG) | 700 g kg−1 metribuzin | C1 |
| 2 June | I | 0.3 L ha−1 Biscaya (Bayer AG) | 240 g L−1 thiacloprid | 4A |
| 20 June | F | 2 kg ha−1 Acrobat Plus WG (BASF SE) | 90 g kg−1 dimethomorph, 600 g kg−1 mancozeb | H5 M3 |
| 28 June | I | 0.3 L ha−1 Biscaya (Bayer AG) | 240 g L−1 thiacloprid | 4A |
| 28 June | F | 2 kg ha−1 Acrobat Plus WG (BASF SE) | 90 g kg−1 dimethomorph, 600 g kg−1 mancozeb | H5 M3 |
| 8 July | F | 1.5 L ha−1 Infinito (Bayer SE) | 62.5 g L−1 fluopicolide, 625,0 g L−1 propamocarb-HCl | B5 F4 |
| 15 July | F | 1.6 L ha−1 Infinito (Bayer SE) | 62.5 g L−1 fluopicolide, 625.0 g L−1 propamocarb-HCl | B5 F4 |
| 15 July | I | 0.3 L ha−1 Biscaya (Bayer AG) | 240 g L−1 thiacloprid | 4A |
| 3 August | H | 0.8 L ha−1 Quickdown (Ceminova Deutschland GmbH & Co. KG) | 24.2 g L−1 pyraflufen | E14 |
| 3 August | H | 2 L ha−1 Toil (Ceminova Deutschland GmbH & Co. KG) | 836 g L−1 rapeseed oil methyl ester | |
| 2017 | ||||
| 5 May | H | 2.0 kg ha−1Artist (Bayer AG) | 240 g kg−1 flufenacet, 175 g kg−1 metribuzin | K3 C1 |
| 2 June | F | 2.0 kg ha−1 Ridomil Gold (Syngenta AG) | 40 g kg−1 metalaxyl-M, 640 g kg−1 mancozeb | A1 M3 |
| 2 June | I | 0.3 L ha−1 Biscaya (Bayer AG) | 240 g L−1 thiacloprid | 4A |
| 16 June | F | 2 kg ha−1 Acrobat Plus WG (BASF SE) | 90 g kg−1 dimethomorph, 600 g kg−1 mancozeb | H5 M3 |
| 16 June | I | 0.3 L ha−1 Biscaya (Bayer AG) | 240 g L−1 thiacloprid | 4A |
| 5 July | I | 0.06 L ha−1 Coragen (DuPont) | 200 g L−1 chlorantraniliprole | 28 |
| 5 July | F | 2 kg ha−1 Acrobat Plus WG (BASF SE) | 90 g kg−1 dimethomorph, 600 g kg−1 mancozeb | H5 M3 |
| 9 August | H | 2.5 L ha−1 Reglone (Syngenta AG) | 374 g L−1 diquat dibromide | D |
References
- Horwith, B. A role for intercropping in modern agriculture. BioScience 1985, 35, 286–291. [Google Scholar] [CrossRef]
- Power, J.F.; Follett, R.F. Monoculture. Sci. Am. 1987, 256, 78–87. [Google Scholar] [CrossRef]
- Gliessman, S.R. Multiple cropping systems: A basis for developing an alternative agriculture. In US Congress Off Ice of Technology Assessment. Innovative Biological Technologies for Lesser Developed Countries: Workshop Proceedings; Congress of the USA: Washington, DC, USA, 1985; pp. 69–83. [Google Scholar]
- Thevathasan, N.V.; Gordon, A.M.; Simpson, J.A.; Reynolds, P.E.; Price, G.; Zhang, P. Biophysical and ecological interactions in a temperate tree-based intercropping system. J. Crop Improv. 2004, 12, 339–363. [Google Scholar] [CrossRef]
- Kremen, C.; Miles, A. Ecosystem services in biologically diversified versus conventional farming systems: Benefits, externalities, and trade-offs. Ecol. Soc. 2012, 17. [Google Scholar] [CrossRef]
- Nair, P.R. The coming of age of agroforestry. J. Sci. Food Agric. 2007, 87, 1613–1619. [Google Scholar] [CrossRef]
- Nair, P.R. Classification of agroforestry systems. Agrofor. Syst. 1985, 3, 97–128. [Google Scholar] [CrossRef]
- Raintree, J.B. Agroforestry pathways: Land tenure, shifting cultivation and sustainable agriculture. Unasylva 1986, 38, 2–15. [Google Scholar]
- Nair, P.R. An Introduction to Agroforestry; Springer Science & Business Media: Berlin, Germany, 1993. [Google Scholar]
- Vandermeer, J.H. The Ecology of Intercropping; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- Montagnini, F.; Nair, P.K.R. Carbon sequestration: An underexploited environmental benefit of agroforestry systems. Agrofor. Syst. 2004, 61, 281–295. [Google Scholar]
- Rigueiro-Rodríguez, A.; Fernández-Núñez, E.; González-Hernández, P.; McAdam, J.H.; Mosquera-Losada, M.R. Agroforestry systems in Europe: Productive, ecological and social perspectives. In Agroforestry in Europe; Springer: Berlin/Heidelberg, Germany, 2009; pp. 43–65. [Google Scholar]
- Roces-Diaz, J.V.; Rolo, V.; Kay, S.; Moreno, G.; Szerencsits, E.; Fagerholm, N.; Plieninger, T.; Torralba, M.; Graves, A.; Giannitsopoulos, M.; et al. Exploring the Relationships among Bio-Physical and Socio-Cultural Ecosystem Services of Agroforestry Systems across Europe. In Proceedings of the Agroforestry as Sustainable Land Use, Nijmegen, The Netherlands, 28–30 May 2018; EURAF: Nijmegen, The Netherlands, 2018. [Google Scholar]
- Elevitch, C.; Mazaroli, D.; Ragone, D. Agroforestry standards for regenerative agriculture. Sustainability 2018, 10, 3337. [Google Scholar] [CrossRef]
- Gao, L.; Xu, H.; Bi, H.; Xi, W.; Bao, B.; Wang, X.; Bi, C.; Chang, Y. Intercropping competition between apple trees and crops in agroforestry systems on the Loess Plateau of China. PLoS ONE 2013, 8, e70739. [Google Scholar] [CrossRef]
- Zhang, W.; Ahanbieke, P.; Wang, B.J.; Xu, W.L.; Li, L.H.; Christie, P.; Li, L. Root distribution and interactions in jujube tree/wheat agroforestry system. Agrofor. Syst. 2013, 87, 929–939. [Google Scholar] [CrossRef]
- Newman, S.M.; Bennett, K.; Wu, Y. Performance of maize, beans and ginger as intercrops in Paulownia plantations in China. Agrofor. Syst. 1997, 39, 23–30. [Google Scholar] [CrossRef]
- Khybri, M.L.; Gupta, R.K.; Ram, S.; Tomar, H.P.S. Crop yields of rice and wheat grown in rotation as intercrops with three tree species in the outer hills of Western Himalaya. Agrofor. Syst. 1992, 17, 193–204. [Google Scholar] [CrossRef]
- Ceccon, E. Production of bioenergy on small farms: A two-year agroforestry experiment using Eucalyptus urophylla intercropped with rice and beans in Minas Gerais, Brazil. New For. 2008, 35, 285–298. [Google Scholar] [CrossRef]
- Friday, J.B.; Fownes, J.H. Competition for light between hedgerows and maize in an alley cropping system in Hawaii, USA. Agrofor. Syst. 2002, 55, 125–137. [Google Scholar] [CrossRef]
- Rahman, M.; Bari, M.; Rahman, M.; Ginnah, M.; Rahman, M. Screening of Potato Varieties under Litchi Based Agroforestry System. Am. J. Exp. Agric. 2016, 14, 1–10. [Google Scholar] [CrossRef]
- Mariana, M.; Hamdani, J.S. Growth and yield of Solanum tuberosum at medium plain with application of paclobutrazol and paranet shade. Agric. Agric. Sci. Procedia 2016, 9, 26–30. [Google Scholar] [CrossRef]
- Pleijel, H.; Danielsson, H.; Vandermeiren, K.; Blum, C.; Colls, J.; Ojanperä, K. Stomatal conductance and ozone exposure in relation to potato tuber yield—Results from the European CHIP Programme. Eur. J. Agron. 2002, 17, 303–317. [Google Scholar] [CrossRef]
- Kareem, I.A. Rattle Tree (ALBIZIA LEBBECK) Effects on Soil Properties and Productivity of Irish Potato (SOLANUM TOBEROSUM) on The Jos Plateau, Nigeria. Ph.D. Thesis, Environmental Resource Planning, University of Jos, Jos, Nigeria, 2007. [Google Scholar]
- Nadir, S.W.; Ng’etich, W.K.; Kebeney, S.J. Performance of crops under Eucalyptus tree-crop mixtures and its potential for adoption in agroforestry systems. Aust. J. Crop Sci. 2018, 12, 1231. [Google Scholar] [CrossRef]
- Li, L.; Ljung, K.; Breton, G.; Schmitz, R.J.; Pruneda-Paz, J.; Cowing-Zitron, C.; Cole, B.J.; Ivans, L.J.; Pedmale, U.V.; Jung, H.-S.; et al. Linking photoreceptor excitation to changes in plant architecture. Genes Dev. 2012, 26, 785–790. [Google Scholar] [CrossRef] [Green Version]
- Abdrabbo, M.A.; Farag, A.A.; Abul-Soud, M. The intercropping effect on potato under net house as adaption procedure of climate change impacts. Appl. Res. 2013, 5, 48–60. [Google Scholar]
- Gawronska, H.; Dwelle, R.B.; Pavek, J.J. Partitioning of photoassimilates by potato plants (Solanum tuberosum L.) as influenced by irradiance: II. Partitioning patterns by four clones grown under high and low irradiance1. Am. J. Potato Res. 1990, 67, 163–176. [Google Scholar] [CrossRef]
- Kooman, P.L.; Fahem, M.; Tegera, P.; Haverkort, A.J. Effects of climate on different potato genotypes 2. Dry matter allocation and duration of the growth cycle. Eur. J. Agron. 1996, 5, 207–217. [Google Scholar] [CrossRef]
- NASA Atmospheric Science Data Center. Available online: https://eosweb.larc.nasa.gov/sse/global/text/lat_tilt_radiation (accessed on 24 December 2017).
- Smith, J.; Pearce, B.D.; Wolfe, M.S. Reconciling productivity with protection of the environment: Is temperate agroforestry the answer? Renew. Agric. Food Syst. 2013, 28, 80–92. [Google Scholar] [CrossRef]
- Barbeau, C.D.; Wilton, M.J.; Oelbermann, M.; Karagatzides, J.D.; Tsuji, L.J. Local food production in a subarctic Indigenous community: The use of willow (Salix spp.) windbreaks to increase the yield of intercropped potatoes (Solanum tuberosum) and bush beans (Phaseolus vulgaris). Int. J. Agric. Sustain. 2018, 16, 29–39. [Google Scholar] [CrossRef]
- Wilton, M.J.; Karagatzides, J.D.; Tsuji, L.J. Nutrient Concentrations of Bush Bean (Phaseolus vulgaris L.) and Potato (Solanum tuberosum L.) Cultivated in Subarctic Soils Managed with Intercropping and Willow (Salix spp.) Agroforestry. Sustainability 2017, 9, 2294. [Google Scholar] [CrossRef]
- BSV Saaten–Blühmischung M2 FAKT Maßnahme E2.1 und E2.2. Available online: https://bsv-saaten.de/bluehmischungen-und-bienenweiden/foerderprogramme-1/fakt-1/naturplus-fakt-m2-zr-100-bluehmischung-m2-fuer-fakt-massnahme-e2-1-und-e2-2.html (accessed on 24 November 2018).
- Röhrig, M.; Sander, R.; eV Geschäftsstelle, I. Interaktive Online-Beratung mit dem Informationssystem Integrierte Pflanzenproduktion (ISIP). In Proceedings of the GIL Jahrestagung, Potsdam, Germany, 6–8 March 2006; pp. 221–224. [Google Scholar]
- Bundesministerium für Ernährung. Landwirtschaft und Verbraucherschutz (BMELV). Gute fachliche Praxis im Pflanzenschutz-Grundsätze für die Durchführung; BMELV: Bonn, Germany, 2010.
- Janssen, W. Online irrigation service for fruit and vegetable crops at farmers site. In Proceedings of the 9th EMS Annual Meeting, 9th European Conference on Applications of Meteorology (ECAM) Abstracts, Toulouse, France, 28 September–2 October 2009; Available online: http://meetings.copernicus.org/ems2009/ (accessed on 5 September 2017).
- Hack, H.; Gall, H.; Klemke, T.H.; Klose, R.; Meier, U.; Stauss, R.; Witzenberger, A. The BBCH scale for phenological growth stages of potato (Solanum tuberosum L.). In Proceedings of the 12th Annual Congress of the European Association for Potato Research, Paris, France, 1993; pp. 153–154. [Google Scholar]
- Schulz, V.S.; Munz, S.; Stolzenburg, K.; Hartung, J.; Weisenburger, S.; Mastel, K.; Möller, K.; Claupein, W.; Graeff-Hönninger, S. Biomass and Biogas Yield of Maize (Zea mays L.) Grown under Artificial Shading. Agriculture 2018, 8, 178. [Google Scholar] [CrossRef]
- Mushagalusa, G.N.; Ledent, J.-F.; Draye, X. Shoot and root competition in potato/maize intercropping: Effects on growth and yield. Environ. Exp. Bot. 2008, 64, 180–188. [Google Scholar] [CrossRef]
- Bodlaender, K.B.A. Influence of Temperature, Radiation and Photoperiod on Development and Yield; Wageningen University: Gelderland, The Netherlands, 1963. [Google Scholar]
- Cao, W.; Tibbitts, T.W. Leaf emergence on potato stems in relation to thermal time. Agron. J. 1995, 87, 474–477. [Google Scholar] [CrossRef]
- Sprenger, H.; Rudack, K.; Schudoma, C.; Neumann, A.; Seddig, S.; Peters, R.; Zuther, E.; Kopka, J.; Hincha, D.K.; Walther, D.; et al. Assessment of drought tolerance and its potential yield penalty in potato. Funct. Plant Biol. 2015, 42, 655–667. [Google Scholar] [CrossRef]
- Fernández, S.D.M.; Aguilar, R.M.; Moreno, Y.S.; Pérez, J.E.R.; León, M.T.C.; Saldaña, H.L. Growth and sugar content of potato tubers in four maturity stages under greenhouse conditions. Rev. Chapingo Ser. Hortic. 2018, 24, 53–67. [Google Scholar] [CrossRef]
- McMaster, G.S.; Wilhelm, W.W. Growing degree-days: One equation, two interpretations. Agric. For. Meteorol. 1997, 87, 291–300. [Google Scholar] [CrossRef]
- Bundessortenamt. Richtlinien für die Durchführung von Landwirtschaftlichen Sortenversuchen; Bundessortenamt: Hannover, Germany, 2000. [Google Scholar]
- Bundessortenamt. Beschreibende Sortenliste Kartoffel—2017; Bundessortenamt: Hannover, Germany, 2017. [Google Scholar]
- Hoffmann, G. VDLUFA-Methodenbuch Band I: Die Untersuchung von Boeden; Loseblattsammlung; VDLUFA-Verlag: Darmstadt, Germany, 1991; ISBN 3-922712-42-8. [Google Scholar]
- Janßen, E. VDLUFA-Methodenbuch Band VII: Umweltanalytik, 3rd ed.; VDLUFA: Darmstadt, Germany, 2003. [Google Scholar]
- Bassler, R. VDLUFA-Methodenbuch, Band III: Die Chemische Untersuchung von Futtermitteln; 3. Aufl., 2. Ergänzungslieferung 1988 und 3. Ergänzungslieferung 1993; VDLUFA-Verlag: Darmstadt, Germany, 1988. [Google Scholar]
- Amtsblatt, D.E.U. Verordnung (EG) Nr. 152/2009 der Kommission vom 27.01. 2009 zur Festlegung der Probenahmeverfahren und Analysemethoden für die amtliche Untersuchung von Futtermitteln. L 2009, 54, 130. [Google Scholar]
- Heinecke, A. Beitrag zur Ermittlung der Biochemischen Ursachen der Schwarzfleckigkeit bei Kartoffeln; Georg-August-Universität Göttingen: Gottingen, Germany, 2007. [Google Scholar]
- Baty, F.; Ritz, C.; Charles, S.; Brutsche, M.; Flandrois, J.-P.; Delignette-Muller, M.-L. A toolbox for nonlinear regression in R: The package nlstools. J. Stat. Softw. 2015, 66, 1–21. [Google Scholar] [CrossRef]
- Fox, J. Nonlinear Regression and Nonlinear Least Squares; Wiley: New York, NY, USA, 2002. [Google Scholar]
- Demagante, A.L.; Zaag, P.V. The response of potato (Solanum spp.) to photoperiod and light intensity under high temperatures. Potato Res. 1988, 31, 73–83. [Google Scholar] [CrossRef]
- Sale, P.J.M. Productivity of vegetable crops in a region of high solar input. II. Yields and efficiencies of water use and energy. Aust. J. Agric. Res. 1973, 24, 751–762. [Google Scholar] [CrossRef]
- Kuruppuarachchi, D.S.P. Intercropped potato (Solanum spp.): Effect of shade on growth and tuber yield in the northwestern regosol belt of Sri Lanka. Field Crops Res. 1990, 25, 61–72. [Google Scholar] [CrossRef]
- Ebwongu, M.; Adipala, E.; Ssekabembe, C.K.; Kyamanywa, S.; Bhagsari, A.S. Effect of intercropping maize and Solanum potato on yield of the component crops in central Uganda. Afr. Crop Sci. J. 2001, 9, 83–96. [Google Scholar] [CrossRef]
- Menzel, C.M. Tuberization in potato at high temperatures: Interaction between temperature and irradiance. Ann. Bot. 1985, 55, 35–39. [Google Scholar] [CrossRef]
- Struik, P.C.; Vreugdenhil, D.; Haverkort, A.J.; Bus, C.B.; Dankert, R. Possible mechanisms of size hierarchy among tubers on one stem of a potato (Solanum tuberosum L.) plant. Potato Res. 1991, 34, 187–203. [Google Scholar] [CrossRef]
- Abu-Zinada, I.A.; Mousa, W.A. Growth and productivity of different potato varieties under Gaza Strip conditions. Int. J. Agric. Crop Sci. 2015, 8, 433. [Google Scholar]
- Stamm, P.; Kumar, P.P. The phytohormone signal network regulating elongation growth during shade avoidance. J. Exp. Bot. 2010, 61, 2889–2903. [Google Scholar] [CrossRef] [PubMed]
- Arsenault, W.J.; LeBlanc, D.A.; Tai, G.C.; Boswall, P. Effects of nitrogen application and seedpiece spacing on yield and tuber size distribution in eight potato cultivars. Am. J. Potato Res. 2001, 78, 301–309. [Google Scholar] [CrossRef]
- Putz, B. Kartoffeln: Züchtung, Anbau, Verwertung; Behr’s Verlag: Hamburg, Germany, 1989. [Google Scholar]
- Struik, P.C.; Haverkort, A.J.; Vreugdenhil, D.; Bus, C.B.; Dankert, R. Manipulation of tuber-size distribution of a potato crop. Potato Res. 1990, 33, 417–432. [Google Scholar] [CrossRef]
- Delaplace, P.; Brostaux, Y.; Fauconnier, M.-L.; du Jardin, P. Potato (Solanum tuberosum L.) tuber physiological age index is a valid reference frame in postharvest ageing studies. Postharvest Biol. Technol. 2008, 50, 103–106. [Google Scholar] [CrossRef]
- O’Brien, P.J.; Firman, D.M.; Allen, E.J. Effects of shading and seed tuber spacing on initiation and number of tubers in potato crops (Solanum tuberosum). J. Agric. Sci. 1998, 130, 431–449. [Google Scholar] [CrossRef] [Green Version]
- de Lucas, M.; Davière, J.-M.; Rodríguez-Falcón, M.; Pontin, M.; Iglesias-Pedraz, J.M.; Lorrain, S.; Fankhauser, C.; Blázquez, M.A.; Titarenko, E.; Prat, S. A molecular framework for light and gibberellin control of cell elongation. Nature 2008, 451, 480. [Google Scholar] [CrossRef]
- Sun, T. Gibberellin-GID1-DELLA: A Pivotal Regulatory Module for Plant Growth and Development. Plant Physiol. 2010, 154, 567–570. [Google Scholar] [CrossRef]
- García-Martinez, J.L.; Gil, J. Light regulation of gibberellin biosynthesis and mode of action. J. Plant Growth Regul. 2001, 20, 354–368. [Google Scholar] [CrossRef]
- Kamiya, Y.; García-Martínez, J.L. Regulation of gibberellin biosynthesis by light. Curr. Opin. Plant Biol. 1999, 2, 398–403. [Google Scholar] [CrossRef]
- Wurr, D.C.E.; Hole, C.C.; Fellows, J.R.; Milling, J.; Lynn, J.R.; O’Brien, P.J. The effect of some environmental factors on potato tuber numbers. Potato Res. 1997, 40, 297–306. [Google Scholar] [CrossRef]
- Kolbe, H.; Stephan-Beckmann, S. Development, growth and chemical composition of the potato crop (Solanum tuberosum L.). II. Tuber and whole plant. Potato Res. 1997, 40, 135–153. [Google Scholar] [CrossRef]
- Ewing, E.E.; Wareing, P.F. Shoot, Stolon, and Tuber Formation on Potato (Solanum tuberosum L.) Cuttings in Response to Photoperiod. Plant Physiol. 1978, 61, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Pohjakalli, O. On the effect of the intensity of light and length of day on the energy economy of certain cultivated plants. Acta Agric. Scand. 1950, 1, 153–175. [Google Scholar] [CrossRef]
- Saha, R.R.; Sarker, A.Z.; Talukder, A.H.M.M.R.; Akter, S.; Golder, P.C. Variability in Growth and Yield of Potato Varieties at Different Locations of Bangladesh. Bangladesh Soc. Hortic. Sci. 2016, 2, 101–110. [Google Scholar]
- Hébert, Y.; Guingo, E.; Loudet, O. The Response of Root/Shoot Partitioning and Root Morphology to Light Reduction in Maize Genotypes. Crop Sci. 2001, 41, 363–371. [Google Scholar] [CrossRef]
- Geigenberger, P.; Kolbe, A.; Tiessen, A. Redox regulation of carbon storage and partitioning in response to light and sugars. J. Exp. Bot. 2005, 56, 1469–1479. [Google Scholar] [CrossRef]
- Manrique, L.A.; Kinry, J.R.; Hodges, T.; Axness, D.S. Dry Matter Production and Radiation Interception of Potato. Crop Sci. 1991, 31, 1044–1049. [Google Scholar] [CrossRef]
- Dam, J.V.; Kooman, P.L.; Struik, P.C. Effects of temperature and photoperiod on early growth and final number of tubers in potato (Solanum tuberosum L.). Potato Res. 1996, 39, 51–62. [Google Scholar]
- Streck, N.A.; de Paula, F.L.M.; Bisognin, D.A.; Heldwein, A.B.; Dellai, J. Simulating the development of field grown potato (Solanum tuberosum L.). Agric. For. Meteorol. 2007, 142, 1–11. [Google Scholar] [CrossRef]
- Sale, P.J.M. Effect of shading at different times on the growth and yield of the potato. Aust. J. Agric. Res. 1976, 27, 557–566. [Google Scholar] [CrossRef]
- Loewe, V.; Delard, C. Stone pine (Pinus pinea L.): An interesting species for agroforestry in Chile. Agrofor. Syst. 2019, 93, 703–713. [Google Scholar] [CrossRef]
- Tekalign, T.; Hammes, P.S. Growth and productivity of potato as influenced by cultivar and reproductive growth—I. Stomatal conductance, rate of transpiration, net photosynthesis, and dry matter production and allocation. Sci. Hortic. 2005, 105, 13–27. [Google Scholar] [CrossRef]
- Tekalign, T.; Hammes, P.S. Growth and productivity of potato as influenced by cultivar and reproductive growth—II. Growth analysis, tuber yield and quality. Sci. Hortic. 2005, 105, 29–44. [Google Scholar] [CrossRef]
- Knowles, N.R.; Knowles, L.O. Manipulating stem number, tuber set, and yield relationships for northern-and southern-grown potato seed lots. Crop Sci. 2006, 46, 284–296. [Google Scholar] [CrossRef]
- Kolbe, H.; Stephan-Beckmann, S. Development, growth and chemical composition of the potato crop (Solanum tuberosum L.). I. leaf and stem. Potato Res. 1997, 40, 111–129. [Google Scholar] [CrossRef]
- Kläring, H.-P.; Krumbein, A. The effect of constraining the intensity of solar radiation on the photosynthesis, growth, yield and product quality of tomato. J. Agron. Crop Sci. 2013, 199, 351–359. [Google Scholar] [CrossRef]
- Gautier, H.; Diakou-Verdin, V.; Bénard, C.; Reich, M.; Buret, M.; Bourgaud, F.; Poëssel, J.L.; Caris-Veyrat, C.; Génard, M. How does tomato quality (sugar, acid, and nutritional quality) vary with ripening stage, temperature, and irradiance? J. Agric. Food Chem. 2008, 56, 1241–1250. [Google Scholar] [CrossRef] [PubMed]
- Burlingame, B.; Mouillé, B.; Charrondière, R. Nutrients, bioactive non-nutrients and anti-nutrients in potatoes. J. Food Compos. Anal. 2009, 22, 494–502. [Google Scholar] [CrossRef]
- NASA POWER Data Access Viewer. Available online: https://power.larc.nasa.gov/data-access-viewer/ (accessed on 1 November 2018).




| Date | Fertilizer | Active Ingredient | Pure Nutrient |
|---|---|---|---|
| 16 April 2015 (DOY 106) | 130 kg ha−1 lime-nitrogen | 20% N, 50% CaO | 26 kg N, 46 kg Ca |
| 300 kg ha−1 ALZON46 | 46% N | 138 kg N | |
| 600 kg ha−1 potassium sulfate with magnesium | 23% P2O5, 9% S | 60 kg P, 54 kg S | |
| 200 kg ha−1 superphosphate 18 | 18% P2O5, 12% S | 16 kg P, 24 kg S | |
| 11 April 2016 (DOY 102) | 350 kg ha−1 lime-nitrogen | 20% N, 50% CaO | 70 kg N, 125 kg Ca |
| 12 April 2016 (DOY 103) | 260 kg ha−1 calcium ammonium nitrate | 27% N | 70 kg N |
| 450 kg ha−1 superphosphate 18 | 18% P2O5, 12% S | 35 kg P, 54 kg S | |
| 1110 kg ha−1 sulphate of potash containing magnesium salt | 30% K2O, 10% MgO, 17% S | 276 kg K, 67 kg Mg, 189 kg S | |
| 13 April 2017 (DOY 103) | 260 kg ha−1 ALZON46 | 46% N | 120 kg N |
| 970 kg ha−1 sulphate of potash containing magnesium salt | 30% K2O, 10% MgO, 17% S | 242 kg K, 58 kg Mg, 165 kg S | |
| 27 April 2017 (DOY 117) | 390 kg ha−1 superphosphate 18 | 18% P2O5, 12% S | 31 kg P 47 kg S |
| Total Solar Irradiance (MJ m−2 day−1) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Year | 2015 | 2016 | 2017 | |||||||
| Time Period | -S (26) | +S (112) | GP (138) | -S (26) | +S (121) | GP (147) | -S (32) | +S (115) | GP (147) | |
| Shading level | 0% | 18.52 | 20.22 | 19.90 | 17.70 | 19.15 | 18.90 | 14.87 | 20.13 | 18.96 |
| 12% ‡ | 17.80 | 17.93 | 16.86 | 17.00 | 17.72 | 17.08 | ||||
| 26% ‡ | 14.97 | 15.64 | 14.17 | 14.80 | 14.90 | 14.89 | ||||
| 50% ‡ | 10.11 | 11.70 | 9.58 | 10.01 | 10.07 | 11.14 | ||||
| Duration of Growing Phases | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| -S | +S | ||||||||
| Year | Shade | P-E | E-F | F-S | S-H | ||||
| GDD | days | GDD | day | GDD | days | GDD | days | ||
| 2016 | 0% | 132 | 26 | 559 c † | 42 | 973 b | 30 | 1689 | 48 |
| 12% | 132 | 26 | 573 b | 43 | 973 b | 29 | 1685 | 48 | |
| 26% | 132 | 26 | 580 b | 44 | 1211 a | 43 | 1685 | 33 | |
| 50% | 137 | 26 | 598 a | 45 | 1211 a | 42 | 1685 | 33 | |
| SEM | 2.24 | 3.62 | 0.00 ‡ | 2.03 | |||||
| p-values $ | |||||||||
| Replicate | 0.422 | 0.422 | 1.000 | 0.422 | |||||
| Shade | 0.455 | 0.002 | <0.0001 | 0.455 | |||||
| 2017 | 0% | 169 | 32 | 440 b | 21 | 1003 | 39 | 1755 | 54 |
| 12% | 173 | 32 | 447 b | 21 | 1016 | 39 | 1768 | 54 | |
| 26% | 173 | 32 | 444 b | 21 | 1011 | 39 | 1764 | 54 | |
| 50% | 169 | 32 | 467 a | 24 | 1007 | 36 | 1760 | 54 | |
| SEM | 2.93 | 2.79 | 4.17 | 4.02 | |||||
| p-values $ | |||||||||
| Replicate | 0.670 | 1.000 | 0.823 | 0.708 | |||||
| Shade | 0.654 | 0.002 | 0.249 | 0.243 | |||||
| Shade | Number of Stems per Plant | Number of Tubers per Plant | Tuber Mass per Plant | Foliage Mass per Plant | Foliage:Tuber Mass Ratio | Total Mass per Plant | HI |
|---|---|---|---|---|---|---|---|
| Year | 2016 | ||||||
| 0% | 2.50 | 10.50 | 44.47 | 48.21 | 1.32 | 92.68 | 0.45 |
| 12% | 3.67 | 12.44 ┴ | 56.70 | 57.06 | 1.25 | 113.76 | 0.48 |
| 26% | 4.17 | 12.83 | 49.81 | 75.45 | 2.45 | 125.25 | 0.36 |
| 50% | 4.17 | 13.67 | 36.28 | 57.05 | 1.81 | 93.33 | 0.39 |
| SEM | 0.85 | 2.79 | 10.97 | 9.98 | 0.53 | 19.66 | 0.04 |
| p-values $ | |||||||
| Replicate | 0.248 | 0.144 | 0.104 | 0.509 | 0.092 | 0.488 | 0.002 |
| Shade | 0.479 | 0.873 | 0.612 | 0.300 | 0.417 | 0.585 | 0.213 |
| Year | 2017 | ||||||
| 0% | 4.83 | 19.00 a † | 103.60 a | 79.37 | 0.95 b | 182.97 a | 0.54 |
| 12% | 3.50 | 17.83 a | 51.28 b | 64.70 | 1.58 b | 115.98 b | 0.43 |
| 26% | 3.17 | 13.17 ab | 66.32 ab | 62.15 | 1.00 b | 128.47 ab | 0.52 |
| 50% | 3.33 | 9.00 b | 32.68 b ║ | 60.07 | 4.67 a ∇ | 86.12 b | 0.25 |
| SEM | 0.51 | 2.47 | 14.05 | 10.59 | 0.92 | 21.15 | 0.06 |
| p-values $ | |||||||
| Replicate | 0.835 | 0.605 | 0.510 | 0.259 | 0.573 | 0.473 | 0.788 |
| Shade | 0.186 | 0.038 | 0.020 | 0.575 | 0.051 | 0.032 | 0.064 |
| Starch | Fraction O | BSB | N | P | K | Ca | Mg | S | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Shade | Undersized | Table | Oversized | ||||||||
| 0% | 70.45 | 4.39 | 74.83 | 19.18 | 16.80 | 1.30 | 0.21 | 2.62 | 0.03 | 0.13 | 0.19 |
| 12% | 71.04 | 4.69 | 77.27 | 17.06 | 19.81 | 1.31 | 0.22 | 2.65 | 0.03 | 0.13 | 0.19 |
| 26% | 70.06 | 7.84 | 76.48 | 12.62 | 19.70 □ | 1.36 | 0.22 | 2.70 | 0.03 | 0.13 | 0.18 |
| 50% | 68.43 | 5.62 | 83.90 | 8.86 | 26.65 | 1.42 | 0.23 | 2.69 | 0.03 | 0.13 | 0.19 |
| SEM | 0.67 | 2.60 | 4.997 | 4.02 | 3.82 □ | 0.05 | 0.01 | 0.07 | 0.00 ‡ | 0.00 ‡ | 0.01 |
| p-values $ | |||||||||||
| Year | 0.043 | 0.034 | 0.071 | 0.011 | 0.157 | 0.066 | <0.0001 | 0.011 | <0.0001 | 0.026 | 0.055 |
| Shade | 0.063 | 0.806 | 0.642 | 0.415 | 0.386 | 0.339 | 0.448 | 0.864 | 0.808 | 0.921 | 0.853 |
| Year x Replicate | 0.424 | 0.705 | 0.234 | 0.028 | 0.011 | 0.206 | 0.299 | 0.287 | 0.104 | 0.043 | 0.138 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schulz, V.S.; Munz, S.; Stolzenburg, K.; Hartung, J.; Weisenburger, S.; Graeff-Hönninger, S. Impact of Different Shading Levels on Growth, Yield and Quality of Potato (Solanum tuberosum L.). Agronomy 2019, 9, 330. https://doi.org/10.3390/agronomy9060330
Schulz VS, Munz S, Stolzenburg K, Hartung J, Weisenburger S, Graeff-Hönninger S. Impact of Different Shading Levels on Growth, Yield and Quality of Potato (Solanum tuberosum L.). Agronomy. 2019; 9(6):330. https://doi.org/10.3390/agronomy9060330
Chicago/Turabian StyleSchulz, Vanessa S., Sebastian Munz, Kerstin Stolzenburg, Jens Hartung, Sebastian Weisenburger, and Simone Graeff-Hönninger. 2019. "Impact of Different Shading Levels on Growth, Yield and Quality of Potato (Solanum tuberosum L.)" Agronomy 9, no. 6: 330. https://doi.org/10.3390/agronomy9060330
APA StyleSchulz, V. S., Munz, S., Stolzenburg, K., Hartung, J., Weisenburger, S., & Graeff-Hönninger, S. (2019). Impact of Different Shading Levels on Growth, Yield and Quality of Potato (Solanum tuberosum L.). Agronomy, 9(6), 330. https://doi.org/10.3390/agronomy9060330