Yield, Quality and Physiological Traits of Red Beet Under Different Magnesium Nutrition and Light Intensity Levels



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Experimental Design and Growing Condition
2.2. Crop Management
2.3. Aerial and Belowground Biomass Accumulation
2.4. Leaf Reflectance and Soil-Plant Analysis Development (SPAD) Readings
2.5. Analytical Determinations
2.5.1. Total Nitrogen and Magnesium Determination, Chlorophyll Content
2.5.2. Individual Carbohydrates
2.5.3. Total Polyphenols Content (TPC) and Betalains Assay
2.6. Statistical Analysis
3. Results
3.1. Storage Root and Leaves Yields
3.2. Nitrogen and Magnesium Accumulation and Chlorophyll Content
3.3. Sugars
3.4. Polyphenols and Betalains Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Akula, R.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Velasco, C.E.; Avila-Sosa, R.; Navarro-Cruz, A.R.; López-Malo, A.; Palou, E. Biotic and abiotic factors to increase bioactive compounds in fruits and vegetables. In Food Bioconversion Handbook of Food Bioengineering; Grumezescu, A.M., Holban, A.M., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 2, pp. 317–349. [Google Scholar]
- Stagnari, F.; Galieni, A.; D’Egidio, S.; Pagnani, G.; Pisante, M. Responses of radish (Raphanus sativus) to drought stress. Ann. Appl. Biol. 2018, 172, 170–186. [Google Scholar] [CrossRef]
- Stagnari, F.; Galieni, A.; D’Egidio, S.; Pagnani, G.; Ficcadenti, N.; Pisante, M. Defoliation and S nutrition on radish: Growth, polyphenols and antiradical activity. Hort. Bras. 2018, 36, 313–319. [Google Scholar] [CrossRef]
- Botella, M.Á.; Arévalo, L.; Mestre, T.C.; Rubio, F.; García-Sánchez, F.; Rivero, R.M.; Martínez, V. Potassium fertilization enhances pepper fruit quality. J. Plant Nutr. 2017, 40, 145–155. [Google Scholar] [CrossRef]
- Michalska, A.; Wojdyło, A.; Bogucka, B. The influence of nitrogen and potassium fertilisation on the content of polyphenolic compounds and antioxidant capacity of coloured potato. J. Food Compos. Anal. 2016, 47, 69–75. [Google Scholar] [CrossRef]
- Cakmak, I.; Yazici, A.M. Magnesium: A forgotten element in crop production. Better Crop. Plant Food 2010, 94, 23–25. [Google Scholar]
- Shaul, O. Magnesium transport and function in plants: The tip of the iceberg. BioMetals 2002, 15, 307–321. [Google Scholar] [CrossRef]
- Cakmak, I.; Kirkby, E.A. Role of magnesium in carbon partitioning and alleviating photooxidative damage. Physiol. Plant. 2008, 133, 692–704. [Google Scholar] [CrossRef]
- Cakmak, I. Magnesium in crop production, food quality and human health. Plant Soil. 2013, 368, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Verbruggen, N.; Hermans, C. Physiological and molecular responses to magnesium nutritional imbalance in plants. Plant Soil. 2013, 368, 87–99. [Google Scholar] [CrossRef]
- Hermans, C.; Bourgis, F.; Faucher, M.; Strasser, R.J.; Delrot, S.; Verbruggen, N. Magnesium deficiency in sugar beets alters sugar partitioning and phloem loading in young mature leaves. Planta 2005, 220, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, I.; Marschner, H. Magnesium deficiency and high light intensity enhance activities of superoxide dismutase, ascorbate peroxidase, and glutathione reductase in bean leaves. Plant Physiol. 1992, 98, 1222–1227. [Google Scholar] [CrossRef] [PubMed]
- Stagnari, F.; Galieni, A.; Pisante, M. Shading and nitrogen management affect quality, safety and yield of greenhouse-grown leaf lettuce. Sci. Hortic. 2015, 192, 70–79. [Google Scholar] [CrossRef]
- Stagnari, F.; Di Mattia, C.; Galieni, A.; Santarelli, V.; D’Egidio, S.; Pagnani, G.; Pisante, M. Light quantity and quality supplies Sharply affect growth, morphological, physiological and quality traits of basil. Ind. Crop. Prod. 2018, 122, 277–289. [Google Scholar] [CrossRef]
- Dai, Y.; Shen, Z.; Liu, Y.; Wang, L.; Hannaway, D.; Lu, H. Effects of Shade treatments on the photosynthetic capacity, chlorophyll fluorescence, and chlorophyll content of Tetrastigma hemsleyanum Diels et Gilg. Environ. Exp. Bot. 2009, 65, 177–182. [Google Scholar] [CrossRef]
- Stagnari, F.; Galieni, A.; Cafiero, G.; Pisante, M. Application of photo-selective films to manipulate wavelength of transmitted radiation and photosynthate composition in red beet (Beta vulgaris var. conditiva Alef.). J. Sci. Food Agric. 2014, 94, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Stagnari, F.; Galieni, A.; Speca, S.; Cafiero, G.; Pisante, M. Effect of light and water supply on morphological and physiological leaf traits of red beet. Agron. J. 2014, 106, 459–468. [Google Scholar] [CrossRef]
- Stagnari, F.; Galieni, A.; Speca, S.; Pisante, M. Water stress effects on growth, yield and quality traits of red beet. Sci. Hortic. 2014, 165, 13–22. [Google Scholar] [CrossRef]
- Galieni, A.; Stagnari, F.; Speca, S.; Pisante, M. Leaf traits as indicators of limiting growing conditions for lettuce (Lactuca sativa). Ann. Appl. Biol. 2016, 169, 342–356. [Google Scholar] [CrossRef]
- Sukhova, E.; Sukhov, V. Analysis of light-induced changes in the photochemical reflectance index (PRI) in leaves of pea, wheat, and pumpkin using pulses of green-yellow measuring light. Remote Sens. 2019, 11, 810. [Google Scholar] [CrossRef]
- Stylinski, C.; Gamon, J.; Oechel, W. Seasonal patterns of reflectance indices, carotenoid pigments and photosynthesis of evergreen chaparral species. Oecologia 2002, 131, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Gitelson, A.; Merzlyak, M.N. Spectral reflectance changes associated with autumn senescence of Aesculus hippocastanum L. and Acer platanoides L. leaves. Spectral features and relation to chlorophyll estimation. J. Plant Physiol. 1994, 143, 286–292. [Google Scholar] [CrossRef]
- Gamon, J.A.; Surfus, J.S. Assessing leaf pigment content and activity with a reflectometer. New Phytol. 1999, 143, 105–117. [Google Scholar] [CrossRef]
- Moreno, D.A.; García-Viguera, C.; Gil, J.I.; Gil-Izquierdo, A. Betalains in the era of global agri-food science, technology and nutritional health. Phytochem. Rev. 2008, 7, 261–280. [Google Scholar] [CrossRef]
- Orlovius, K.; McHoul, J. Effect of Two Magnesium Fertilizers on Leaf Magnesium Concentration, Yield, and Quality of Potato and Sugar Beet. J. Plant Nut. 2015, 38, 2044–2054. [Google Scholar] [CrossRef]
- Brewster, J.L.; Sutherland, R.A. The rapid determination in controlled environments of parameters for predicting seedling growth rates in natural conditions. Ann. Appl. Biol. 1993, 122, 123–133. [Google Scholar] [CrossRef]
- Rouse, J. Monitoring the Vernal Advancement of Retrogradation of Natural Vegetation; Type III Final Report; NASA/GSFG: Washington, DC, USA, 1974; p. 371.
- Gamon, J.A.; Filella, I.; Peñuelas, J. The Dynamic 531-Nanometer Δ Reflectance Signal: A Survey of Twenty Angiosperm Species. In Photosynthetic Responses to the Environment, American Society of Plant Physiologists; Yamamoto, H.Y., Smhh, C.M., Eds.; American Society of Plant Physiologists: Derwood, MD, USA, 1993; pp. 172–177. [Google Scholar]
- Penuelas, J.; Filella, I.; Gamon, J.A. Assessment of photosynthetic radiation-use efficiency with spectral reflectance. New Phytol. 1995, 131, 291–296. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1. [Google Scholar] [CrossRef]
- Karkacier, M.; Erbas, M.; Uslu, M.K.; Aksu, M. Comparison of different extraction and detection methods for sugars using amino-bonded phase HPLC. J. Chromatogr. Sci. 2003, 41, 331–333. [Google Scholar] [CrossRef]
- Nurmi, K.; Ossipov, V.; Haukioja, E.; Pihlaja, K. Variation of total phenolic content and individual low-molecular-weight phenolics in foliage of mountain birch trees (Betula pubescens ssp. tortuosa). J. Chem. Ecol. 1996, 22, 2023–2040. [Google Scholar] [CrossRef]
- Cardoso-Ugarte, G.A.; Sosa-Morales, M.E.; Ballard, T.; Liceaga, A.; San Martín-González, M.F. Microwave-assisted extraction of betalains from red beet (Beta vulgaris). LWT-Food Sci. Technol. 2014, 59, 276–282. [Google Scholar] [CrossRef]
- Cowan, J.A. Structural and catalytic chemistry of magnesium-dependent enzymes. BioMetals 2002, 15, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Mengel, K.; Kirkby, E.A. Principles of Plant Nutrition; International PotaLR Institute: Worblaufen-Bern, Switzerland, 1987; pp. 481–492. [Google Scholar]
- Guo, W.; Nazim, H.; Liang, Z.; Yang, D. Magnesium deficiency in plants: An urgent problem. Crop J. 2016, 4, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Farhat, N.; Rabhi, M.; Krol, M.; Barhoumi, Z.; Ivanov, A.G.; McCarthy, A.; Abdelly, C.; Smaoui, A.; Hüner, N.P.A. Starch and sugar accumulation in Sulla carnosa leaves upon Mg2+ starvation. Acta Physiol. Plant. 2014, 36, 2157–2165. [Google Scholar] [CrossRef]
- Hermans, C.; Verbruggen, N. Physiological characterization of Mg deficiency in Arabidopsis thaliana. J. Exp. Bot. 2005, 56, 2153–2161. [Google Scholar] [CrossRef] [PubMed]
- Jezek, M.; Geilfus, C.M.; Bayer, K.H. Mühling. Photosynthetic capacity, nutrient status, and growth of maize (Zea mays L.) upon a MgSO4 leaf-application. Front. Plant Sci. 2015, 5, 781. [Google Scholar] [CrossRef] [PubMed]
- Hermans, C.; Johnson, G.N.; Strasser, R.J.; Verbruggen, N. Physiological characterisation of magnesium deficiency in sugar beet: Acclimation to low magnesium differentially affects photosystems I and II. Planta 2004, 220, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H.; Cakmak, I. High light intensity enhances chlorosis and necrosis in leaves of zinc, potassium, and magnesium deficient bean (Phaseolus vulgaris) plants. J. Plant Physiol. 1989, 134, 308–315. [Google Scholar] [CrossRef]
- Cakmak, I.; Hengeler, C.; Marschner, H. Partitioning of shoot and root dry matter and carbohydrates in bean plants suffering from phosphorus, potassium and magnesium deficiency. J. Exp. Bot. 1994, 45, 1245–1250. [Google Scholar] [CrossRef]
- Ding, Y.; Luo, W.; Xu, G. Characterisation of magnesium nutrition and interaction of magnesium and potassium in rice. Ann. Appl. Biol. 2006, 149, 111–123. [Google Scholar] [CrossRef]
- Casa, R.; Castaldi, F.; Pascucci, S.; Pignatti, S. Chlorophyll estimation in field crops: An assessment of handheld leaf meters and spectral reflectance measurements. J. Agric. Sci. 2015, 153, 876–890. [Google Scholar] [CrossRef]
- Haboudane, D.; Miller, J.R.; Tremblay, N.; Zarco-Tejada, P.J.; Dextraze, L. Integrated narrow-band vegetation indices for prediction of crop chlorophyll content for application to precision agriculture. Remote Sens. Environ. 2002, 81, 416–426. [Google Scholar] [CrossRef]
- Montes-Lora, S.; Rodríguez-Pulido, F.J.; Cejudo-Bastante, M.J.; Heredia, F.J. Implications of the red beet ripening on the colour and betalain composition relationships. Plant Foods Hum. Nutr. 2018, 73, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Grzebisz, W. Crop response to magnesium fertilization as affected by nitrogen supply. Plant Soil. 2013, 368, 23–39. [Google Scholar] [CrossRef]
- Yin, S.; Ze, Y.G.; Liu, C.; Li, N.; Zhou, M.; Duan, Y.M.; Hong, F.S. Cerium relieves the inhibition of nitrogen metabolism of spinach caused by magnesium deficiency. Biol. Trace Elem. Res. 2009, 132, 247–258. [Google Scholar] [CrossRef] [PubMed]


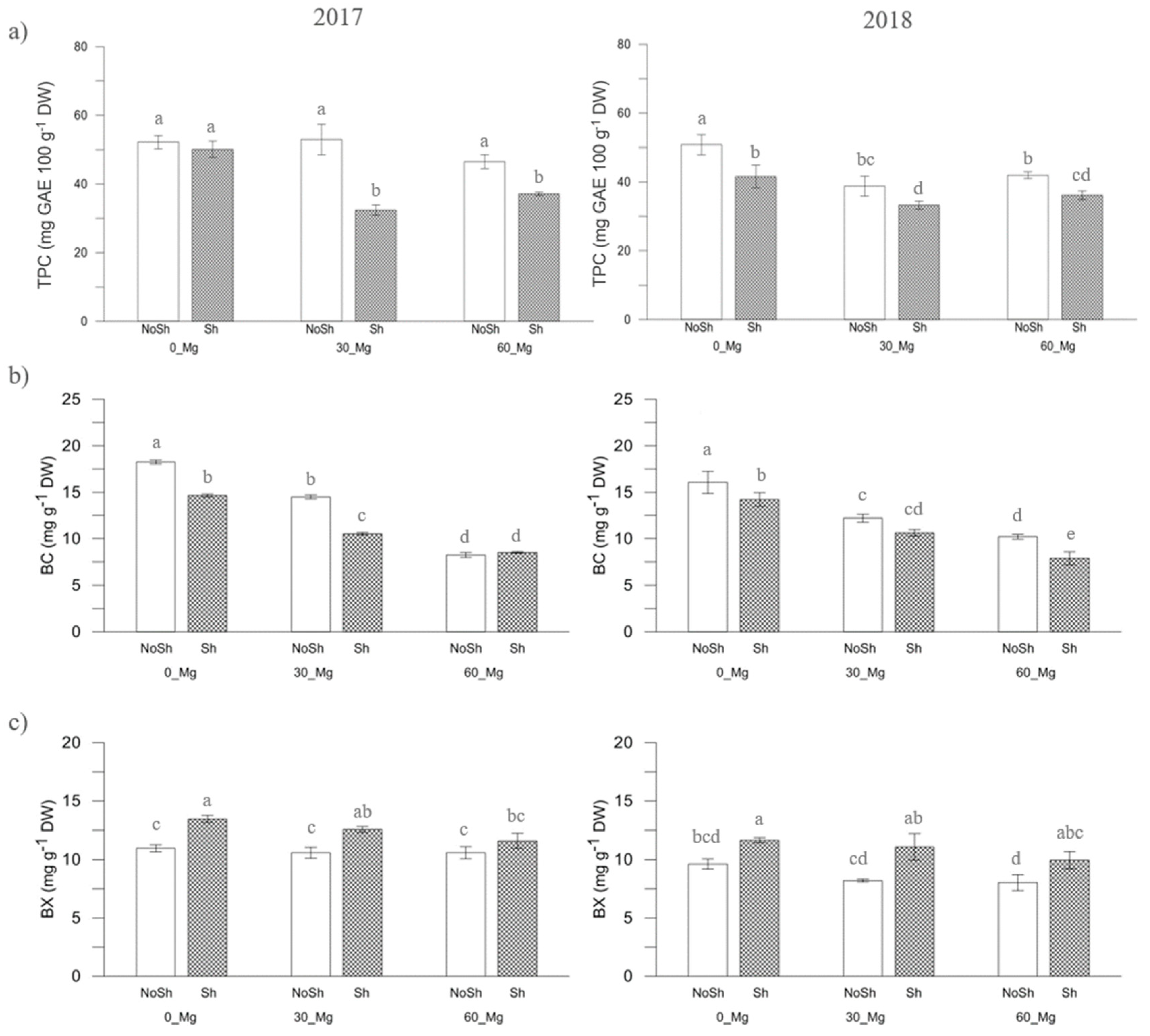{kind=link}
| 2017 | 2018 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatments | Leaves (g plant−1) | Storage Root (g plant−1) | Leaves (g plant−1) | Storage Root (g plant−1) | ||||||||
| FL | LR | o.m.† | FL | LR | o.m. | FL | LR | o.m. | FL | LR | o.m. | |
| Leaves | ||||||||||||
| MG_0 | 0.68 | 0.72 | 0.70 C | 1.56 | 1.60 | 1.58 C | 0.88 | 0.86 | 0.87 B | 1.31 c | 1.17 c | 1.24 |
| MG_30 | 0.95 | 1.18 | 1.07 B | 6.05 | 6.15 | 6.09 B | 1.57 | 1.83 | 1.70 A | 4.90 b | 7.61 a | 6.25 |
| MG_60 | 1.52 | 1.63 | 1.58 A | 7.67 | 7.84 | 7.75 A | 1.57 | 2.13 | 1.84 A | 6.86 a | 7.42 a | 7.13 |
| o.m. | 1.05 | 1.18 | 5.09 | 5.19 | 1.34 B | 1.60 A | 4.35 | 5.40 | ||||
| PAR availability | n.s. | n.s. | **(0.050) | *(0.266) | ||||||||
| Mg rates | **(0.034) | **(0.111) | **(0.146) | **(0.267) | ||||||||
| PAR × Mg | n.s. | n.s. | n.s. | ** (0.378) | ||||||||
| 2017 | 2018 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatments | N (mg g−1 DW) | Mg (mg g−1 DW) | N (mg g−1 DW) | Mg (mg g−1 DW) | ||||||||
| FL | LR | o.m.† | FL | LR | o.m. | FL | LR | o.m. | FL | LR | o.m. | |
| Leaves | ||||||||||||
| MG_0 | 15.54 | 15.83 | 15.69 B | 4.06 e | 5.14 d | 4.60 | 13.85 | 12.59 | 13.22 B | 7.35 c | 6.50 d | 6.92 |
| MG_30 | 17.85 | 19.03 | 18.44 AB | 5.05 d | 10.41 b | 7.73 | 16.75 | 16.32 | 16.54 A | 13.05 c | 14.65 b | 13.85 |
| MG_60 | 20.15 | 19.65 | 19.90 A | 8.64 c | 11.82 a | 10.23 | 16.19 | 18.45 | 17.32 A | 17.22 a | 21.72 a | 19.47 |
| o.m. | 17.85 | 18.17 | 5.92 | 9.12 | 15.60 | 15.78 | 12.54 | 1.429 | ||||
| PAR availability | n.s. | **(0.013) | n.s. | n.s. | ||||||||
| Mg rates | **(0.909) | **(0.026) | **(0.980) | **(0.048) | ||||||||
| PAR × Mg | n.s. | **(0.036) | n.s. | ** (0.068) | ||||||||
| Storage root | ||||||||||||
| MG_0 | 10.49 | 11.53 | 11.01 B | 2.16 d | 1.85 e | 2.01 | 10.47 | 10.18 | 10.32 B | 2.06 d | 1.94 d | 2.00 |
| MG_30 | 12.50 | 12.09 | 12.29 A | 3.04 b | 3.92 b | 3.48 | 12.09 | 10.76 | 11.43 A | 2.89 b | 3.69 a | 3.29 |
| MG_60 | 13.08 | 12.61 | 12.85 A | 2.78 a | 3.11 c | 2.94 | 12.12 | 10.95 | 11.54 A | 2.53 c | 3.20 b | 2.86 |
| o.m. | 12.02 | 12.07 | 2.66 | 2.96 | 11.56 A | 10.63 B | 2.49 | 2.94 | ||||
| PAR availability | n.s. | **(0.002) | **(0.190) | **(0.005) | ||||||||
| Mg rates | *(0.590) | **(0.007) | **(0.301) | **(0.009) | ||||||||
| PAR × Mg | n.s. | **(0.010) | n.s. | **(0.013) | ||||||||
| Chlorophyll Content (µg mg−1 FW) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | GDD | Treatments | ChlA | ChlB | ChlTot | ChlA / ChlB | ||||||||
| 2017 | FL | LR | o.m.† | FL | LR | o.m. | FL | LR | o.m. | FL | LR | o.m. | ||
| 143 | MG_0 | 0.58 c | 0.73 c | 0.65 | 0.29 c | 0.30 c | 0.29 | 0.87 c | 1.03 c | 0.95 | 2.01 c | 2.42 c | 2.22 | |
| MG_30 | 1.80 b | 3.10 a | 2.45 | 0.50 b | 0.61 a | 0.56 | 2.30 b | 3.71 a | 3.01 | 3.58 b | 5.08 a | 4.33 | ||
| MG_60 | 1.95 b | 2.96 a | 2.46 | 0.59 a | 0.59 a | 0.59 | 2.54 b | 3.55 a | 3.05 | 3.29 b | 5.06 a | 4.17 | ||
| o.m. | 1.44 | 2.26 | 0.46 | 0.50 | 1.90 | 2.76 | 2.96 | 4.18 | ||||||
| PAR availability | **(0.071) | **(0.008) | **(0.072) | **(0.147) | ||||||||||
| Mg rates | **(0.087) | **(0.014) | **(0.083) | **(0.220) | ||||||||||
| PAR × Mg | **(0.123) | **(0.020) | **(0.118) | *(0.311) | ||||||||||
| 358 | MG_0 | 0.75 c | 0.80 c | 0.77 | 0.32 c | 0.45 c | 0.38 | 1.07 c | 1.24 c | 1.15 | 2.40 b | 1.79 c | 2.09 | |
| MG_30 | 1.14 b | 2.54 a | 1.84 | 0.49 b | 0.80 a | 0.65 | 1.63 b | 3.34 a | 2.48 | 2.31 b | 3.18 a | 2.75 | ||
| MG_60 | 1.16 b | 2.66 a | 1.91 | 0.62 b | 0.87 a | 0.74 | 1.78 b | 3.53 a | 2.65 | 1.88 c | 3.08 a | 2.48 | ||
| o.m. | 1.01 | 2.00 | 0.48 | 0.70 | 1.49 | 2.70 | 2.19 | 2.68 | ||||||
| PAR availability | **(0.059) | **(0.012) | **(0.052) | *(0.119) | ||||||||||
| Mg rates | **(0.071) | **(0.029) | **(0.087) | **(0.144) | ||||||||||
| PAR × Mg | **(0.101) | *(0.041) | **(0.124) | **(0.203) | ||||||||||
| 714 | MG_0 | 0.80 d | 0.66 d | 0.73 | 0.31 e | 0.41 d | 0.36 | 1.11 d | 1.08 d | 1.09 | 2.56 ab | 1.63 d | 2.09 | |
| MG_30 | 1.25 c | 2.51 a | 1.88 | 0.55 c | 0.92 a | 0.74 | 1.80 c | 3.43 a | 2.62 | 2.27 bc | 2.74 a | 2.51 | ||
| MG_60 | 1.55 b | 2.58 a | 2.06 | 0.79 b | 0.98 a | 0.88 | 2.34 b | 3.56 a | 2.95 | 1.96 cd | 2.64 ab | 2.30 | ||
| o.m. | 1.20 | 1.92 | 0.55 | 0.77 | 1.75 | 2.69 | 2.26 | 2.34 | ||||||
| PAR availability | **(0.058) | **(0.015) | **(0.064) | n.s. | ||||||||||
| Mg rates | **(0.053) | **(0.027) | **(0.063) | *(0.122) | ||||||||||
| PAR × Mg | **(0.076) | **(0.039) | **(0.090) | ** (0.173) | ||||||||||
| 2018 | 158 | MG_0 | 0.44 c | 0.53 c | 0.48 | 0.23 | 0.26 | 0.24 B | 0.67 | 0.78 | 0.73 b | 1.92 b | 2.07 b | 1.99 |
| MG_30 | 1.46 b | 2.78 a | 2.12 | 0.76 | 0.67 | 0.71 A | 2.22 | 3.45 | 2.83 a | 1.95 b | 4.17 a | 3.06 | ||
| MG_60 | 1.30 b | 2.65 a | 1.97 | 0.55 | 0.59 | 0.57 A | 1.85 | 3.24 | 2.54 a | 2.35 b | 4.63 a | 3.49 | ||
| o.m. | 1.07 | 1.99 | 0.51 | 0.50 | 1.58 A | 2.49 B | 2.07 | 3.63 | ||||||
| PAR availability | **(0.112) | n.s. | ** (0.140) | **(0.579) | ||||||||||
| Mg rates | **(0.228) | **(0.065) | **(0.285) | **(0.521) | ||||||||||
| PAR × Mg | *(0.323) | n.s. | n.s. | **(0.737) | ||||||||||
| 376 | MG_0 | 0.55 | 0.65 | 0.60 B | 0.30 | 0.47 | 0.38 B | 0.85 b | 1.11 b | 0.98 | 1.80 | 1.41 | 1.60 B | |
| MG_30 | 1.02 | 2.36 | 1.69 A | 0.44 | 1.17 | 0.80 A | 1.46 b | 3.53 a | 2.49 | 2.32 | 2.14 | 2.23 AB | ||
| MG_60 | 1.13 | 2.73 | 1.93 A | 0.55 | 0.81 | 0.68 AB | 1.68 b | 3.53 a | 2.60 | 2.08 | 3.48 | 2.78 A | ||
| o.m. | 0.90 B | 1.91 A | 0.43 B | 0.81 A | 1.33 | 2.72 | 2.07 | 2.34 | ||||||
| PAR availability | ** (0.141) | ** (0.078) | **(0.209) | *(0.078) | ||||||||||
| Mg rates | **(0.278) | *(0.112) | **(0.301) | *(0.389) | ||||||||||
| PAR × Mg | n.s. | n.s. | *(0.426) | n.s. | ||||||||||
| 724 | MG_0 | 0.52 d | 0.59 d | 0.56 | 0.23 d | 0.32 d | 0.28 | 0.75 d | 0.91 d | 0.83 | 2.35 ab | 1.82 c | 2.08 | |
| MG_30 | 1.21 c | 2.34 a | 1.77 | 0.50 c | 1.15 a | 0.82 | 1.71 c | 3.48 a | 2.60 | 2.45 a | 2.04 bc | 2.24 | ||
| MG_60 | 1.62 b | 2.40 a | 2.01 | 0.85 b | 0.92 b | 0.89 | 2.47 b | 3.32 a | 2.90 | 1.90 c | 2.64 a | 2.27 | ||
| o.m. | 1.12 | 1.78 | 0.53 | 0.80 | 1.64 | 2.57 | 2.23 | 2.17 | ||||||
| PAR availability | **(0.060) | **(0.047) | **(0.105) | n.s. | ||||||||||
| Mg rates | **(0.054) | **(0.035) | **(0.079) | n.s. | ||||||||||
| PAR × Mg | **(0.076) | **(0.050) | **(0.111) | **(0.173) | ||||||||||
| Year | GDD | Treatments | NDVI670 | PRI | CI | SPAD | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2017 | FL | LR | o.m.† | FL | LR | o.m. | FL | LR | o.m. | FL | LR | o.m. | ||
| 143 | MG_0 | 0.366 | 0.333 | 0.350 B | −0.042 | −0.042 | −0.042 B | 0.165 | 0.179 | 0.172 B | 40.6 c | 39.9 c | 40.3 | |
| MG_30 | 0.443 | 0.459 | 0.451 AB | −0.029 | −0.024 | −0.026 A | 0.252 | 0.255 | 0.254 AB | 48.9 b | 54.7 a | 51.8 | ||
| MG_60 | 0.573 | 0.513 | 0.543 A | −0.024 | −0.030 | −0.027 A | 0.333 | 0.286 | 0.310 A | 47.1 b | 53.2 a | 50.2 | ||
| o.m. | 0.461 | 0.435 | −0.031 | −0.032 | 0.250 | 0.240 | 45.5 | 49.3 | ||||||
| PAR availability | n.s. | n.s. | n.s. | *(0.990) | ||||||||||
| Mg rates | **(0.045) | *(0.005) | **(0.029) | ** (1.271) | ||||||||||
| PAR × Mg | n.s. | n.s. | n.s. | *(1.798) | ||||||||||
| 358 | MG_0 | 0.393 | 0.434 | 0.414 | −0.058 | −0.048 | −0.053 | 0.155 | 0.188 | 0.171 | 35.2 | 39.6 | 37.4 B | |
| MG_30 | 0.448 | 0.466 | 0.457 | −0.055 | −0.041 | −0.048 | 0.210 | 0.229 | 0.219 | 45.5 | 54.8 | 50.2 A | ||
| MG_60 | 0.464 | 0.524 | 0.494 | −0.048 | −0.039 | −0.044 | 0.238 | 0.279 | 0.259 | 48.7 | 55.2 | 52.0 A | ||
| o.m. | 0.435 | 0.475 | −0.054 A | −0.043 B | 0.201 | 0.232 | 43.1 B | 49.9 A | ||||||
| PAR availability | n.s. | *(0.003) | n.s. | ** (1.156) | ||||||||||
| Mg rates | n.s. | n.s. | n.s. | ** (1.639) | ||||||||||
| PAR × Mg | n.s. | n.s. | n.s. | n.s. | ||||||||||
| 714 | MG_0 | 0.564 | 0.576 | 0.570 AB | −0.044 | −0.031 | −0.037 B | 0.270 | 0.295 | 0.282 B | 46.3 | 41.5 | 43.9 B | |
| MG_30 | 0.481 | 0.531 | 0.506 B | −0.044 | −0.032 | −0.038 B | 0.241 | 0.267 | 0.254 B | 52.0 | 52.0 | 52.0 A | ||
| MG_60 | 0.616 | 0.652 | 0.634 A | −0.036 | −0.027 | −0.031 A | 0.326 | 0.331 | 0.328 A | 48.9 | 55.7 | 52.3 A | ||
| o.m. | 0.554 | 0.586 | −0.041 B | −0.030 A | 0.279 | 0.298 | 49.1 | 49.8 | ||||||
| PAR availability | n.s. | **(0.001) | n.s. | n.s. | ||||||||||
| Mg rates | **(0.027) | **(0.001) | **(0.029) | ** (2.090) | ||||||||||
| PAR × Mg | n.s. | n.s. | n.s. | n.s. | ||||||||||
| 2018 | 158 | MG_0 | 0.423 | 0.475 | 0.449 B | −0.012 | 0.000 | −0.006 | 0.140 | 0.132 | 0.136 B | 24.4 d | 29.4 c | 25.9 |
| MG_30 | 0.592 | 0.617 | 0.604 A | −0.006 | −0.002 | −0.004 | 0.193 | 0.180 | 0.187 A | 31.5 c | 36.4 b | 34.0 | ||
| MG_60 | 0.564 | 0.672 | 0.618 A | −0.014 | −0.005 | −0.010 | 0.188 | 0.226 | 0.207 A | 35.9 b | 47.8 a | 41.9 | ||
| o.m. | 0.526 B | 0.588 A | −0.010 | −0.002 | 0.174 | 0.179 | 29.9 | 37.9 | ||||||
| PAR availability | *(0.020) | n.s. | n.s. | **(0.945) | ||||||||||
| Mg rates | **(0.022) | n.s. | **(0.014) | **(1.160) | ||||||||||
| PAR × Mg | n.s. | n.s. | n.s. | **(1.640) | ||||||||||
| 376 | MG_0 | 0.569 | 0.567 | 0.568 | −0.026 | −0.024 | −0.025 | 0.315 | 0.290 | 0.303 | 31.0 d | 31.6 d | 31.3 | |
| MG_30 | 0.658 | 0.530 | 0.594 | −0.025 | −0.028 | −0.026 | 0.335 | 0.250 | 0.292 | 33.7 cd | 39.0 b | 36.4 | ||
| MG_60 | 0.565 | 0.704 | 0.634 | −0.026 | −0.010 | −0.018 | 0.289 | 0.353 | 0.321 | 36.0 bc | 50.7 a | 43.4 | ||
| o.m. | 0.597 | 0.600 | −0.025 | −0.020 | 0.313 | 0.298 | 33.6 | 40.5 | 37.0 | |||||
| PAR availability | n.s. | n.s. | n.s. | **(1.055) | ||||||||||
| Mg rates | n.s. | n.s. | n.s. | **(0.964) | ||||||||||
| PAR × Mg | n.s. | n.s. | n.s. | **(1.364) | ||||||||||
| 724 | MG_0 | 0.461 | 0.465 | 0.463 B | −0.037 | −0.041 | −0.039 B | 0.220 | 0.218 | 0.219 B | 30.6 e | 39.3 c | 35.0 | |
| MG_30 | 0.625 | 0.672 | 0.649 A | −0.015 | −0.018 | −0.016 A | 0.337 | 0.330 | 0.334 A | 35.2 d | 48.9 b | 42.1 | ||
| MG_60 | 0.605 | 0.709 | 0.657 A | −0.018 | −0.011 | −0.014 A | 0.323 | 0.393 | 0.358 A | 37.4 d | 51.8 a | 44.6 | ||
| o.m. | 0.564 | 0.615 | −0.023 | −0.023 | 0.293 | 0.314 | 34.4 | 46.7 | ||||||
| PAR availability | n.s. | n.s. | n.s. | **(0.471) | ||||||||||
| Mg rates | **(0.025) | **(0.003) | **(0.023) | **(0.604) | ||||||||||
| PAR × Mg | n.s. | n.s. | n.s. | **(0.854) | ||||||||||
| Sugars Content (g 100 g−1 DW) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | Treatments | Sucrose | Glucose | Fructose | Total Sugars | |||||||||
| 2017 | FL | LR | o.m.† | FL | LR | o.m. | FL | LR | o.m. | FL | LR | o.m. | ||
| Leaves | MG_0 | 4.03 | 3.57 | 3.80 A | 1.37 c | 1.43 bc | 1.40 | 0.97 | 1.03 | 1.00 A | 6.43 | 5.97 | 6.20 A | |
| MG_30 | 2.43 | 3.33 | 2.88 AB | 1.07 d | 1.70 ab | 1.38 | 0.70 | 0.73 | 0.72 B | 4.20 | 5.77 | 4.98 B | ||
| MG_60 | 2.27 | 1.87 | 2.07 B | 1.07 d | 1.97 a | 1.52 | 0.63 | 0.80 | 0.72 B | 3.97 | 4.63 | 4.30 B | ||
| o.m. | 2.91 | 2.92 | 1.19 | 1.68 | 0.77 | 0.86 | 4.87 | 5.46 | ||||||
| PAR availability | n.s. | **(0.183) | n.s. | n.s. | ||||||||||
| Mg rates | **(0.358) | n.s. | *(0.079) | **(0.350) | ||||||||||
| PAR × Mg | n.s. | **(0.025) | n.s. | n.s. | ||||||||||
| Storage root | MG_0 | 48.70 | 52.27 | 50.48 B | 0.83 | 0.73 | 0.78 | 0.23 | 0.13 | 0.18 B | 49.77 | 53.13 | 51.45 B | |
| MG_30 | 52.20 | 54.33 | 53.27 AB | 0.93 | 0.97 | 0.95 | 0.43 | 0.53 | 0.48 A | 53.57 | 55.83 | 54.70 AB | ||
| MG_60 | 56.57 | 53.23 | 54.90 A | 0.87 | 1.00 | 0.93 | 0.37 | 0.47 | 0.42 A | 57.80 | 54.70 | 56.25 A | ||
| o.m. | 52.49 | 53.28 | 0.88 | 0.90 | 0.34 | 0.38 | 53.71 | 54.56 | ||||||
| PAR availability | n.s. | n.s. | n.s. | n.s. | ||||||||||
| Mg rates | *(1.365) | n.s. | **(0.060) | *(1.365) | ||||||||||
| PAR × Mg | n.s. | n.s. | n.s. | n.s. | ||||||||||
| 2018 | ||||||||||||||
| Leaves | MG_0 | 4.43 | 3.17 | 3.80 A | 1.07 | 1.40 | 1.23 | 0.90 | 0.93 | 0.92 A | 6.40 | 5.50 | 5.95 A | |
| MG_30 | 2.70 | 2.17 | 2.43 B | 1.00 | 1.33 | 1.17 | 0.53 | 0.73 | 0.63 B | 4.23 | 4.23 | 4.23 B | ||
| MG_60 | 2.40 | 1.80 | 2.10 B | 0.97 | 1.33 | 1.15 | 0.57 | 0.60 | 0.58 B | 3.93 | 3.73 | 3.83 B | ||
| o.m. | 3.18 | 2.38 | 1.01 B | 1.36 A | 0.67 | 0.76 | 4.86 | 4.49 | ||||||
| PAR availability | n.s. | *(0.095) | n.s. | n.s. | ||||||||||
| Mg rates | **(0.178) | n.s. | **(0.077) | **(0.229) | ||||||||||
| PAR × Mg | n.s. | n.s. | n.s. | n.s. | ||||||||||
| Storage root | MG_0 | 41.10 | 41.53 | 41.32 B | 0.67 ab | 0.60 ab | 0.63 | 0.33 | 0.20 | 0.27 A | 42.10 | 42.33 | 42.22 B | |
| MG_30 | 55.40 | 49.20 | 52.30 A | 0.77 a | 0.57 ab | 0.63 | 0.30 | 0.23 | 0.27 A | 56.47 | 50.00 | 53.23 A | ||
| MG_60 | 50.93 | 52.13 | 51.53 A | 0.47 b | 0.73 a | 0.63 | 0.20 | 0.13 | 0.17 B | 51.60 | 53.00 | 52.30 A | ||
| o.m. | 49.14 | 47.62 | 0.63 | 0.63 | 0.28 A | 0.19 B | 50.06 | 48.44 | ||||||
| PAR availability | n.s. | n.s. | **(0.015) | n.s. | ||||||||||
| Mg rates | *(1.918) | n.s. | **(0.025) | **(1.879) | ||||||||||
| PAR × Mg | n.s. | *(0.108) | n.s. | n.s. | ||||||||||
| 2017 | 2018 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatments | BC + BX | BC/BX | BC + BX | BC/BX | ||||||||
| FL | LR | o.m.† | FL | LR | o.m. | FL | LR | o.m. | FL | LR | o.m. | |
| MG_0 | 29.22 a | 28.12 a | 28.67 | 1.66 a | 1.09 c | 1.38 | 25.50 | 26.08 | 25.79 A | 1.65 | 1.24 | 1.44 A |
| MG_30 | 25.10 b | 23.10 c | 24.10 | 1.38 b | 0.84 d | 1.11 | 19.98 | 22.15 | 21.06 B | 1.44 | 1.02 | 1.23 AB |
| MG_60 | 18.84 d | 20.13 d | 19.49 | 0.79 d | 0.74 d | 0.76 | 17.20 | 18.88 | 18.04 C | 1.15 | 0.91 | 1.03 B |
| o.m. | 24.38 | 23.78 | 1.28 | 0.89 | 20.90 | 22.37 | 1.41 | 1.06 | ||||
| PAR availability | n.s. | **(0.043) | n.s. | n.s. | ||||||||
| Mg rates | **(0.487) | **(0.032) | **(0.905) | **(0.066) | ||||||||
| PAR × Mg | *(0.689) | **(0.046) | n.s. | n.s. | ||||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Egidio, S.; Galieni, A.; Stagnari, F.; Pagnani, G.; Pisante, M. Yield, Quality and Physiological Traits of Red Beet Under Different Magnesium Nutrition and Light Intensity Levels. Agronomy 2019, 9, 379. https://doi.org/10.3390/agronomy9070379
D’Egidio S, Galieni A, Stagnari F, Pagnani G, Pisante M. Yield, Quality and Physiological Traits of Red Beet Under Different Magnesium Nutrition and Light Intensity Levels. Agronomy. 2019; 9(7):379. https://doi.org/10.3390/agronomy9070379
Chicago/Turabian StyleD’Egidio, Sara, Angelica Galieni, Fabio Stagnari, Giancarlo Pagnani, and Michele Pisante. 2019. "Yield, Quality and Physiological Traits of Red Beet Under Different Magnesium Nutrition and Light Intensity Levels" Agronomy 9, no. 7: 379. https://doi.org/10.3390/agronomy9070379
APA StyleD’Egidio, S., Galieni, A., Stagnari, F., Pagnani, G., & Pisante, M. (2019). Yield, Quality and Physiological Traits of Red Beet Under Different Magnesium Nutrition and Light Intensity Levels. Agronomy, 9(7), 379. https://doi.org/10.3390/agronomy9070379