Effects of Water Stress on Gas Exchange, Water Relations and Leaf Structure in Two Ornamental Shrubs in the Mediterranean Area





Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Conditions and Plant Materials
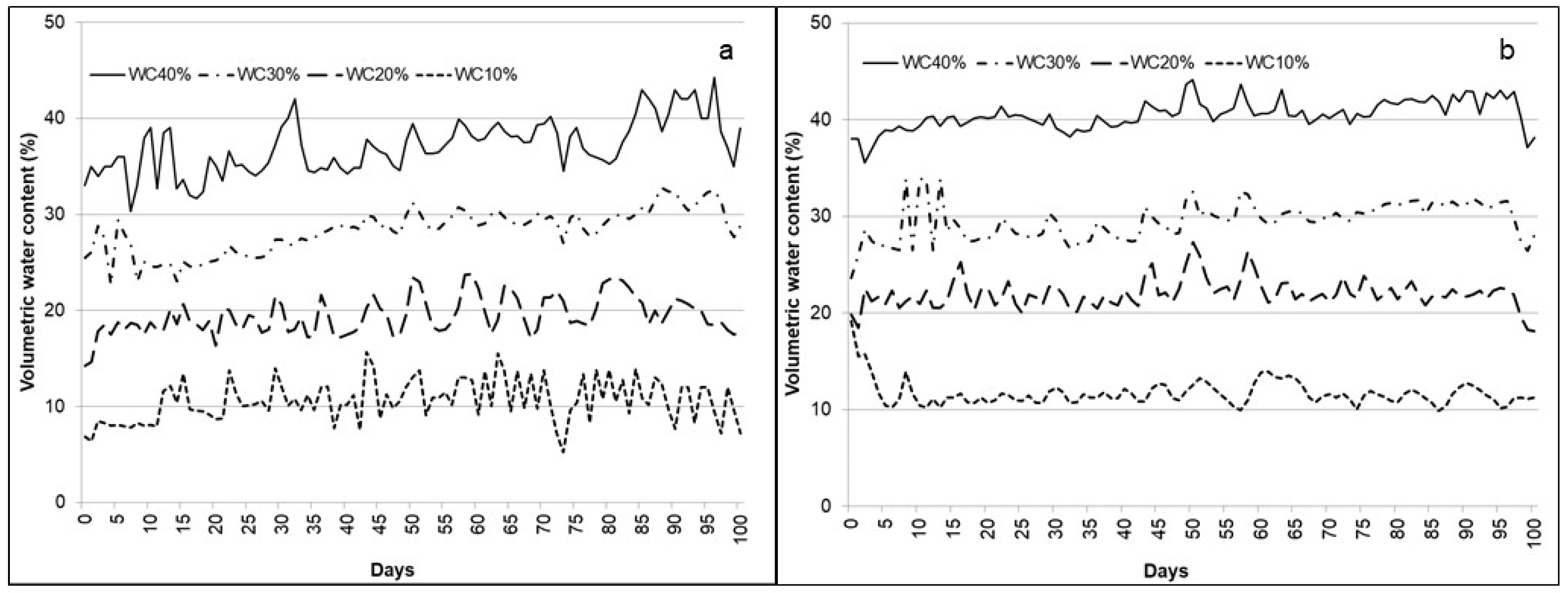
2.2. Substrate Moisture Measurements and Water Supply
2.3. Meteorological Data
2.4. Biomass, Leaf Area and SPAD Index
2.5. Leaf Anatomy
2.6. Plant Physiological Measurements
2.6.1. Leaf Gas Exchanges and Chlorophyll a Fluorescence
2.6.2. Leaf Water Content
2.7. Statistical Analysis
3. Results
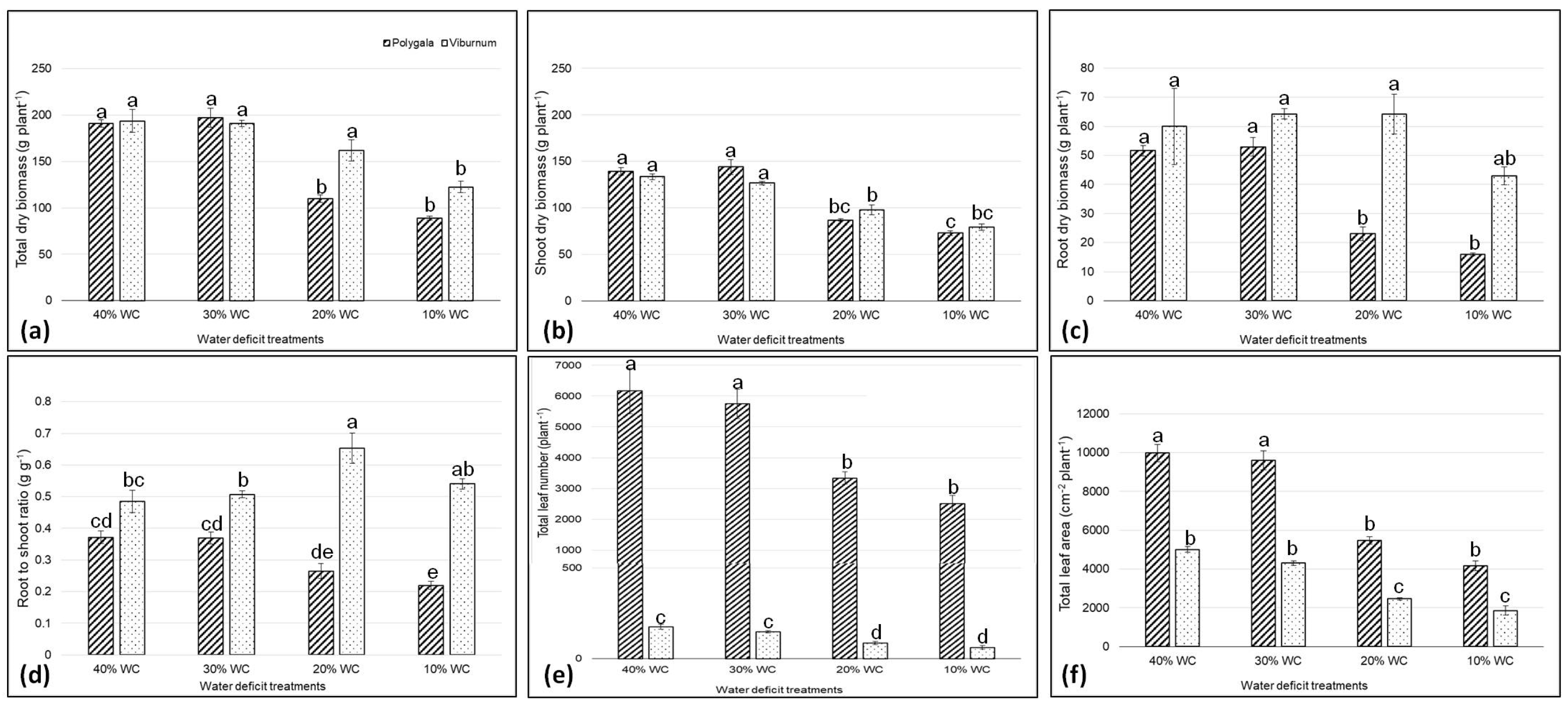
3.1. Biomass and Leaf Area
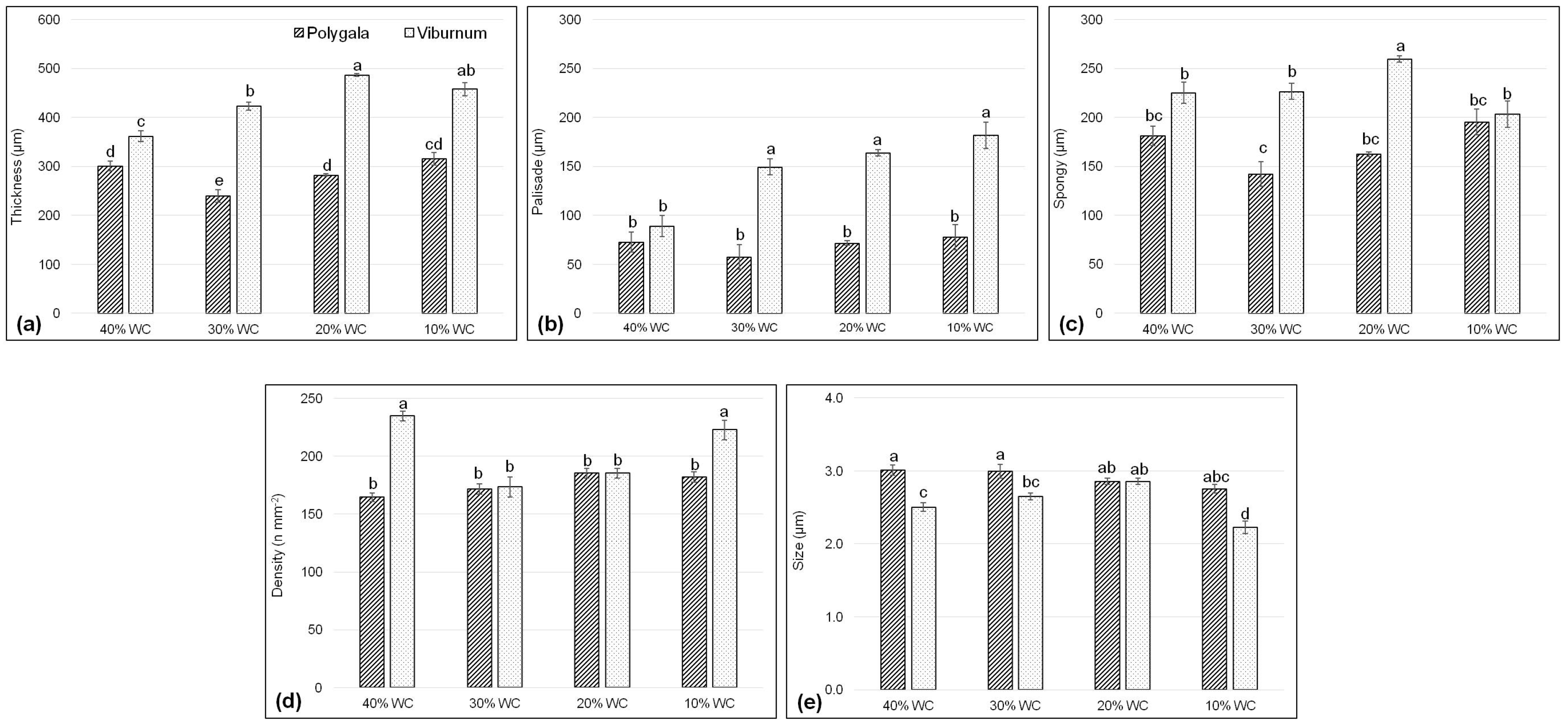
3.2. Leaf Anatomy
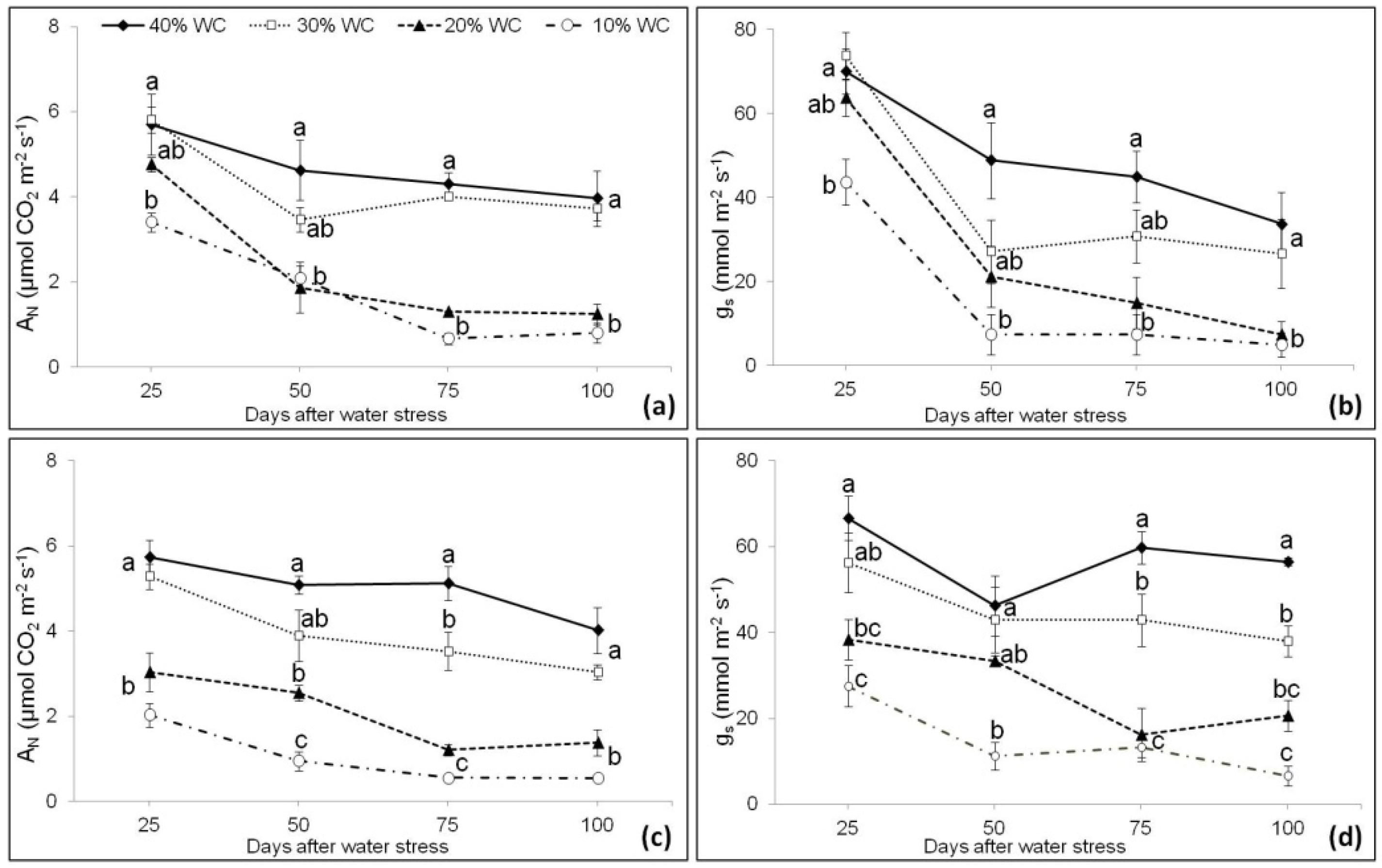
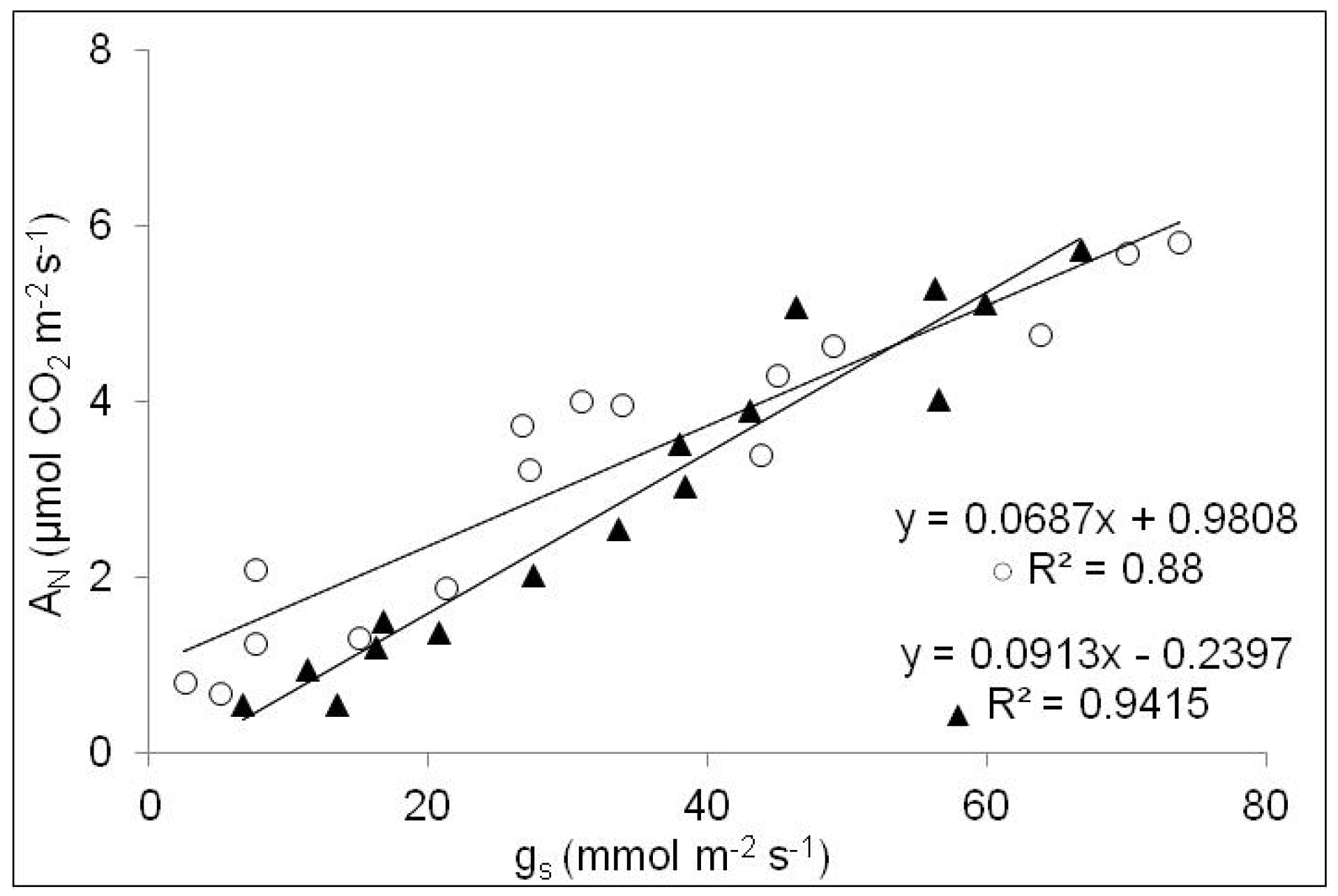
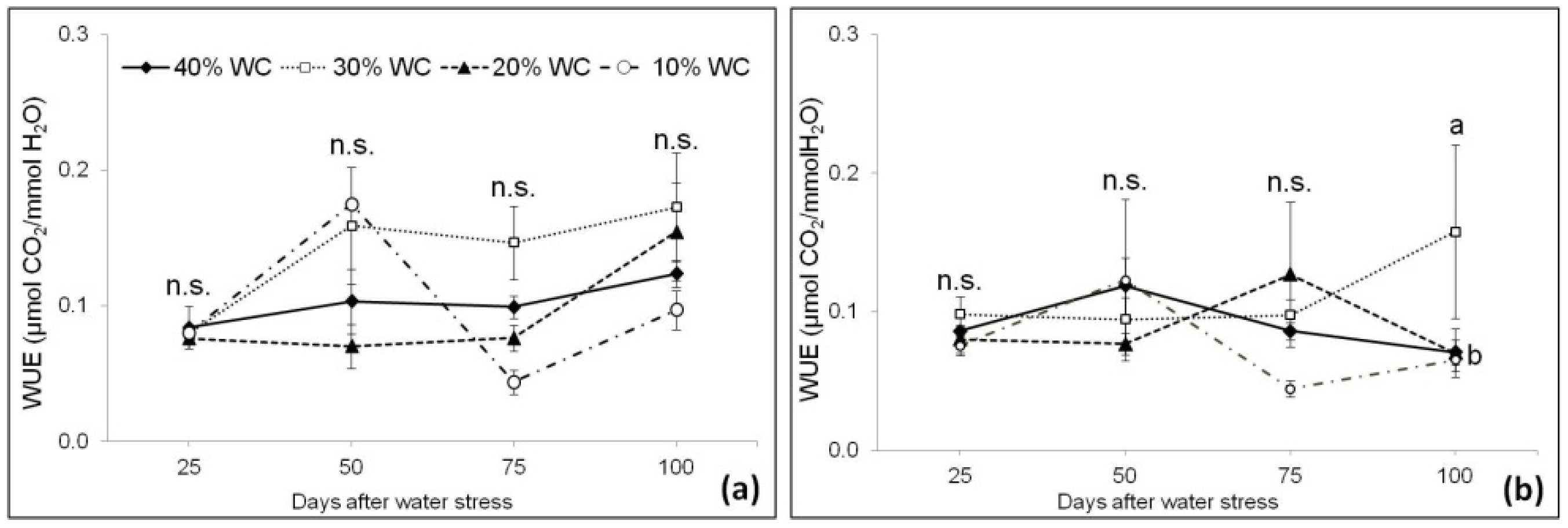
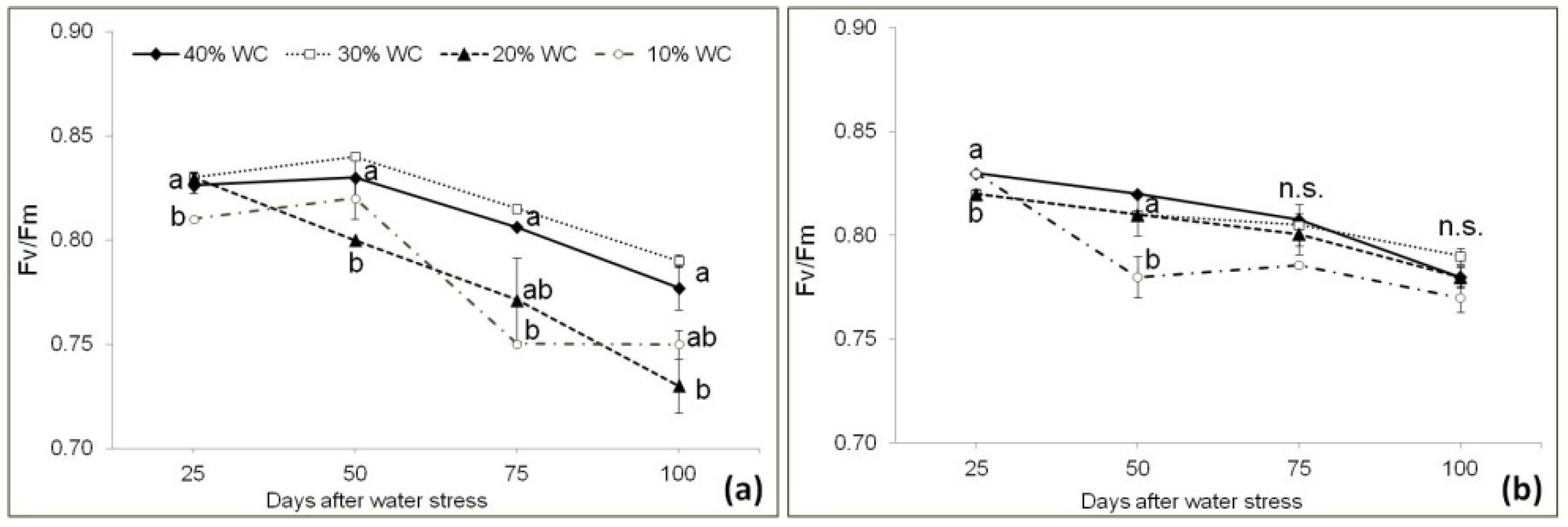
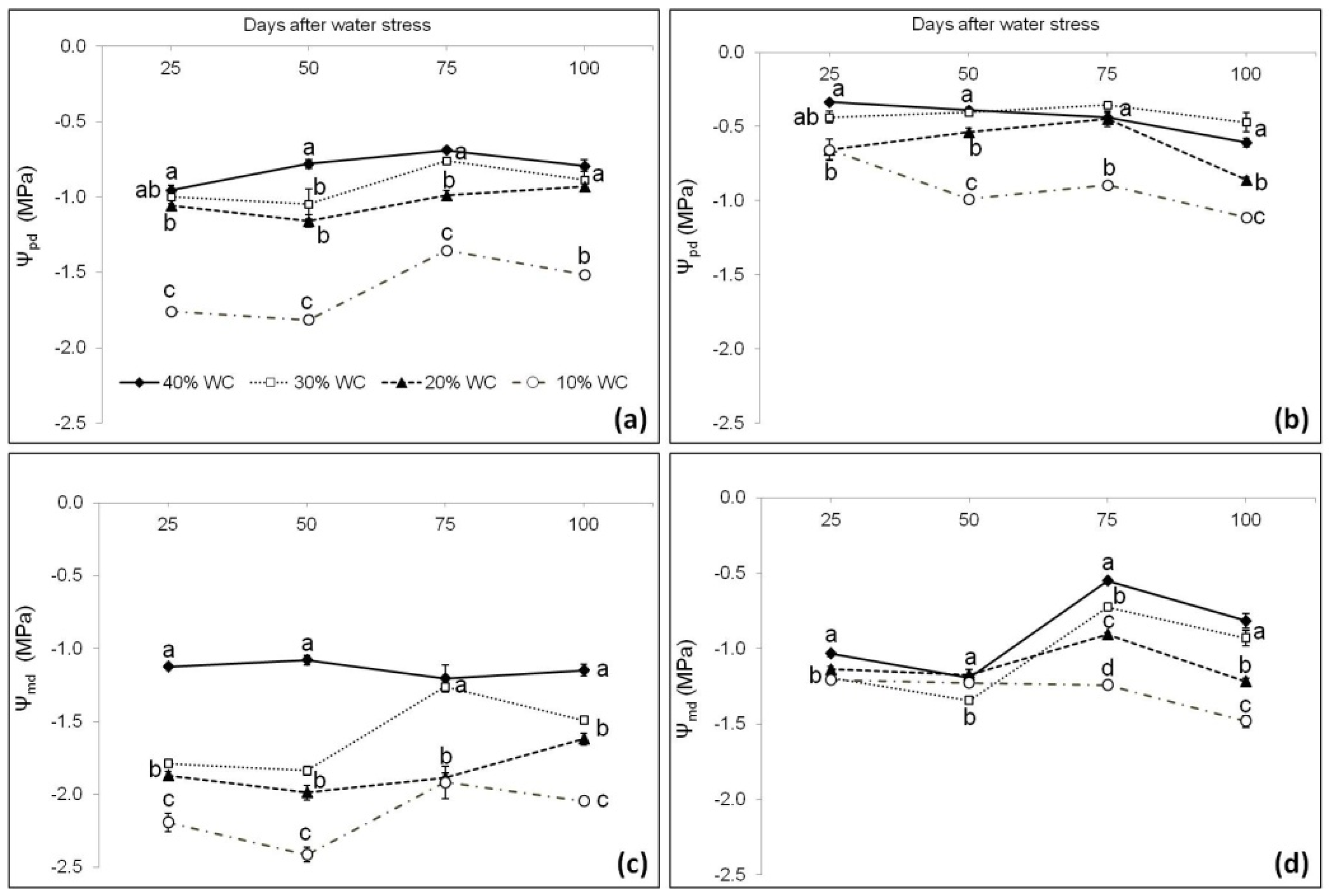
3.3. Plant Physiological Parameters
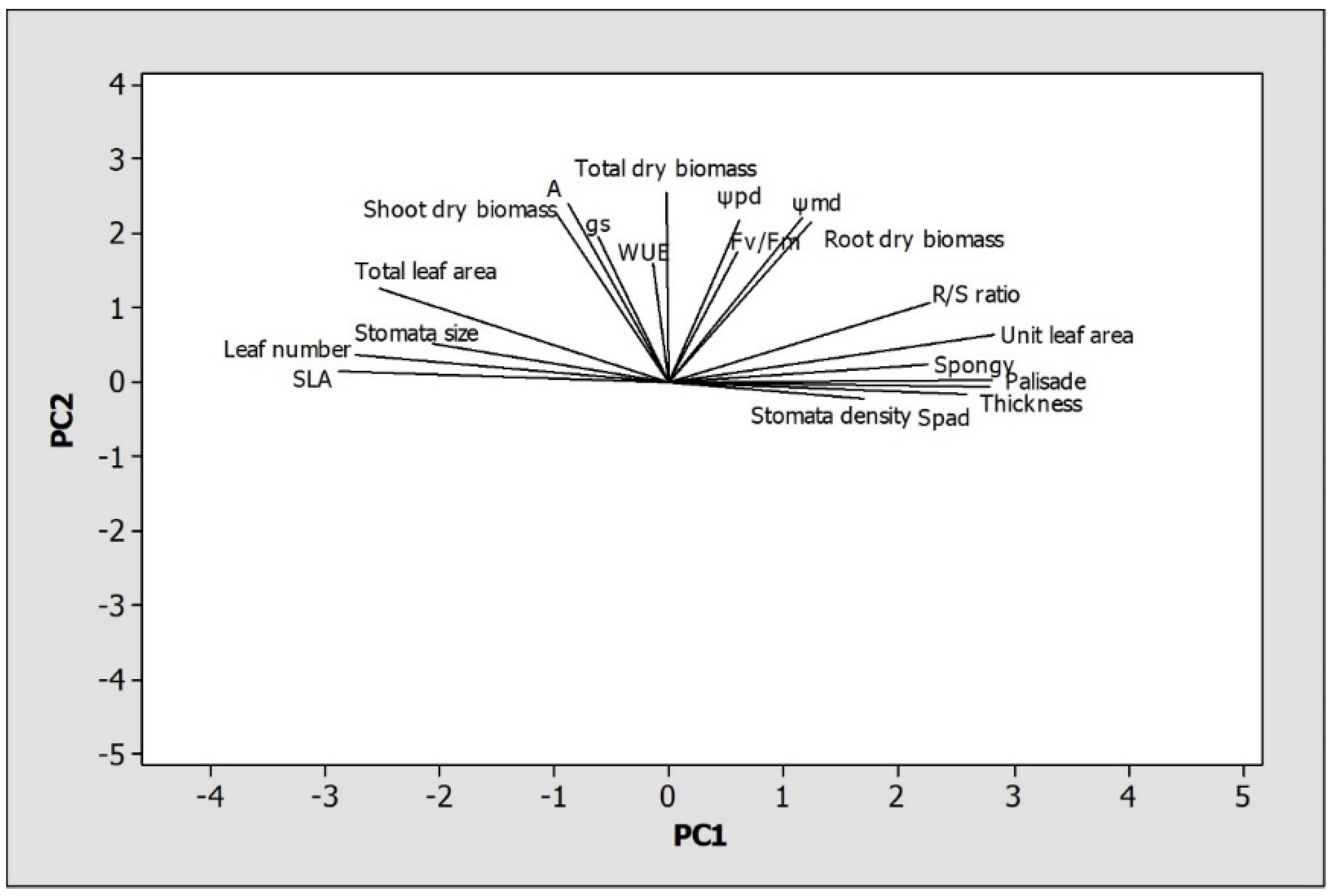
3.4. Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- WWAP (World Water Assessment Programme). The United Nations World Water Development Report 2014: Water and Energy; UNESCO: Paris, France, 2014. [Google Scholar]
- Paz, S.; Negev, M.; Clermont, A.; Green, M.S. Health aspects of climate change in cities with Mediterranean climate, and local adaptation plans. Int. J. Environ. Res. Public Health 2016, 13, 438. [Google Scholar] [CrossRef] [PubMed]
- Dallman, P.R. Plant Life in the World’s Mediterranean Climates: California, Chile, South Africa, Australia, and the Mediterranean Basin; University of California Press: Oakland, CA, USA, 1998. [Google Scholar]
- Rundel, P.W.; Arroyo, M.T.; Cowling, R.M.; Keeley, J.E.; Lamont, B.B.; Vargas, P. Mediterranean biomes: Evolution of their vegetation, floras, and climate. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 383–407. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Peñuelas, J. Drought-induced oxidative stress in strawberry tree (Arbutus unedo L.) growing in Mediterranean field conditions. Plant Sci. 2004, 166, 1105–1110. [Google Scholar] [CrossRef]
- Filella, I.; Llusia, J.; Piñol, J.; Peñuelas, J. Leaf gas exchange and fluorescence of Phillyrea latifolia, Pistacia lentiscus and Quercus ilex saplings in severe drought and high temperature conditions. Environ. Exp. Bot. 1998, 39, 213–220. [Google Scholar] [CrossRef]
- Ferrante, A.; Trivellini, A.; Scuderi, D.; Romano, D.; Vernieri, P. Post-production physiology and handling of ornamental potted plants. Postharvest Biol. Technol. 2015, 100, 99–108. [Google Scholar] [CrossRef]
- Álvarez, S.; Sánchez-Blanco, M.J. Changes in growth rate, root morphology and water use efficiency of potted Callistemon citrinus plants in response to different levels of water deficit. Sci. Hortic. 2013, 156, 54–62. [Google Scholar] [CrossRef]
- Medrano, H.; Flexas, J.; Galmés, J. Variability in water use efficiency at the leaf level among Mediterranean plants with different growth forms. Plant Soil 2009, 317, 17–29. [Google Scholar] [CrossRef]
- Álvarez, S.; Rodríguez, P.; Broetto, F.; Sánchez-Blanco, M.J. Long term responses and adaptive strategies of Pistacia lentiscus under moderate and severe deficit irrigation and salinity: Osmotic and elastic adjustment, growth, ion uptake and photosynthetic activity. Agric. Water Manag. 2018, 202, 253–262. [Google Scholar] [CrossRef]
- Toscano, S.; Ferrante, A.; Romano, D. Response of Mediterranean ornamental plants to drought stress. Horticulturae 2019, 5, 6. [Google Scholar] [CrossRef]
- Cirillo, C.; Rouphael, Y.; Caputo, R.; Raimondi, G.; De Pascale, S. The influence of deficit irrigation on growth, ornamental quality, and water use efficiency of three potted Bougainvillea genotypes grown in two shapes. Hort. Sci 2014, 49, 1284–1291. [Google Scholar] [CrossRef]
- Toscano, S.; Ferrante, A.; Tribulato, A.; Romano, D. Leaf physiological and anatomical responses of Lantana and Ligustrum species under different water availability. Plant Physiol. Biochem. 2018, 127, 380–392. [Google Scholar] [CrossRef] [PubMed]
- Fraser, L.H.; Greenall, A.; Carlyle, C.; Turkington, R.; Ross Friedman, C. Adaptive phenotypic plasticity of Pseudoroegneria spicata: Response of stomatal density, leaf area and biomass to changes in water supply and increased temperature. Ann. Bot. 2009, 103, 769–775. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Stützel, H. Biomass partitioning, specific leaf area, and water use efficiency of vegetable amaranth (Amaranthus spp.) in response to drought stress. Sci. Hortic. 2004, 102, 15–27. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. In Sustainable Agriculture; Springer: Dordrecht, The Netherlands, 2009; pp. 153–188. [Google Scholar]
- Vallejo, V.R.; Bautista, S.; Cortina, J. Restoration for soil protection after disturbances. In Life and Environment in the Mediterranean; Trabaud, L., Ed.; Wit Press: Southampton, UK, 2000; pp. 301–343. [Google Scholar]
- Vilagrosa, A.; Hernández, E.I.; Luis, V.C.; Cochard, H.; Pausas, J.G. Physiological differences explain the co-existence of different regeneration strategies in Mediterranean ecosystems. New Phytol. 2014, 201, 1277–1288. [Google Scholar] [CrossRef] [PubMed]
- Anjum, S.; Xie, X.Y.; Wang, L.C.; Saleem, M.F.; Man, C.; Wang, L. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar] [CrossRef]
- Cirillo, C.; De Micco, V.; Rouphael, Y.; Balzano, A.; Caputo, R.; De Pascale, S. Morpho-anatomical and physiological traits of two Bougainvillea genotypes trained to two shapes under deficit irrigation. Trees 2017, 31, 173–187. [Google Scholar] [CrossRef]
- Campbell, D.R.; Wu, C.A.; Travers, S.E. Photosynthetic and growth responses of reciprocal hybrids to variation in water and nitrogen availability. Am. J. Bot. 2010, 97, 925–933. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Demmig, B.; Björkman, O. Comparison of the effect of excessive light on chlorophyll fluorescence (77K) and photon yield of O2 evolution in leaves of higher plants. Planta 1987, 171, 171–184. [Google Scholar] [CrossRef]
- Huxley, A.J.; Griffiths, M. New Royal Horticultural Society Dictionary of Gardening; Grove’s Dictionaries Inc.: New York, NY, USA, 1999. [Google Scholar]
- Meerow, A.W.; Ayala-Silva, T. Polygala myrtifolia ‘Chapman Field’, an everblooming subtropical flowering shrub. HortScience 2005, 40, 864–865. [Google Scholar] [CrossRef]
- Fini, A.; Ferrini, F.; Frangi, P.; Amoroso, G.; Giordano, C. Growth, leaf gas exchange and leaf anatomy of three ornamental shrubs grown under different light intensities. Eur. J. Hortic. Sci. 2010, 75, 111–117. [Google Scholar]
- Bussotti, F.; Ferrini, F.; Pollastrini, M.; Fini, A. The challenge of Mediterranean sclerophyllous vegetation under climate change: From acclimation to adaptation. Environ. Exper. Bot. 2014, 103, 80–98. [Google Scholar] [CrossRef]
- Cirillo, C.; Rouphael, Y.; Caputo, R.; Raimondi, G.; Sifola, M.I.; De Pascale, S. Effects of high salinity and the exogenous application of an osmolyte on growth, photosynthesis, and mineral composition in two ornamental shrubs. J. Hortic. Sci. Biotechnol. 2016, 91, 14–22. [Google Scholar] [CrossRef]
- Garcıa-Navarro, M.C.; Evans, R.Y.; Montserrat, R.S. Estimation of relative water use among ornamental landscape species. Sci. Hortic. 2004, 99, 163–174. [Google Scholar] [CrossRef]
- Gómez-Bellot, M.J.; Ortuño, M.F.; Nortes, P.A.; Bernavé, A.; Fernández, F.; Sánchez-Blanco, M.J. Effectiveness of bacterial inoculation in alleviation of salinity on water status, mineral content, gas exchange and photosynthetic parameters of Viburnum tinus L. plants. Sci. Hortic. 2018, 237, 303–310. [Google Scholar] [CrossRef]
- Cirillo, C.; De Micco, V.; Arena, C.; Carillo, P.; Pannico, A.; De Pascale, S.; Rouphael, Y. Biochemical, Physiological and anatomical mechanisms of adaptation of Callistemon citrinus and Viburnum lucidum to NaCl and CaCl2 salinization. Front Plant Sci. 2019, 10, 742. [Google Scholar] [CrossRef] [PubMed]
- Farieri, E.; Toscano, S.; Ferrante, A.; Romano, D. Identification of ornamental shrubs tolerant to saline aerosol for coastal urban and peri-urban greening. Urban For. Urban. Green. 2016, 18, 9–18. [Google Scholar] [CrossRef]
- Starr, J.L.; Paltineanu, I.C. Methods for Measurement of Soil Water Content: Capacitance Devices; Methods of Soil Analysis; United States Department of Agriculture, Agricultural Research Service: Washington, DC, USA, 2002. [Google Scholar]
- Schreiber, U.; Schliwa, U.; Bilger, W. Continuous recording of photochemical and non-photochemical chlorophyll fluorescence quenching with a new type of modulation fluorometer. Photosynth. Res. 1986, 10, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Scholander, P.F.; Bradstreet, E.D.; Hemmingsen, E.A.; Hammel, H.T. Sap pressure in vascular plants: Negative hydrostatic pressure can be measured in plants. Science 1965, 148, 339–346. [Google Scholar] [CrossRef]
- Lockett, L.; Montague, T.; McKenney, C.; Auld, D. Assessing public opinion on water conservation and water conserving landscapes in the semiarid southwestern United States. HortTechnology 2002, 12, 392–396. [Google Scholar] [CrossRef]
- Cicevan, R.; Al Hassan, M.; Sestras, A.F.; Prohens, J.; Vicente, O.; Sestras, R.E.; Boscaiu, M. Screening for drought tolerance in cultivars of the ornamental genus Tagetes (Asteraceae). PeerJ 2016, 4, e2133. [Google Scholar] [CrossRef]
- Niu, G.; Rodriguez, D.S.; Aguiniga, L.; Mackay, W. Salinity tolerance of Lupinus havardii and Lupinus texenis. HortScience 2007, 42, 526–528. [Google Scholar] [CrossRef]
- Álvarez, S.; Navarro, A.; Nicolás, E.; Sánchez-Blanco, M.J. Transpiration, photosynthetic responses, tissue water relations and dry mass partitioning in Callistemon plants during drought conditions. Sci. Hortic. 2011, 129, 306–312. [Google Scholar] [CrossRef]
- Elansary, H.O.; Salem, M.Z.M. Morphological and physiological responses and drought resistance enhancement of ornamental shrubs by trinexapac-ethyl application. Sci. Hortic. 2015, 189, 1–11. [Google Scholar] [CrossRef]
- Toscano, S.; Scuderi, D.; Giuffrida, F.; Romano, D. Responses of Mediterranean ornamental shrubs to drought stress and recovery. Sci. Hortic. 2014, 178, 145–153. [Google Scholar] [CrossRef]
- Toscano, S.; Farieri, E.; Ferrante, A.; Romano, D. Physiological and biochemical responses in two ornamental shrubs to drought stress. Front. Plant Sci. 2016, 7, 645. [Google Scholar] [CrossRef]
- Sánchez-Blanco, M.J.; Rodriguez, P.; Morales, M.A.; Torrecillas, A. Comparative growth and water relations of Cistus albidus and Cistus monspeliensis plants during water deficit conditions and recovery. Plant Sci. 2002, 162, 107–113. [Google Scholar] [CrossRef]
- Franco, J.A.; Martinéz-Sanchéz, J.J.; Fernández, J.A.; Bañón, S. Selection and nursery production of ornamental plants for landscaping and xerogardening in semi-arid and environments. J. Hortic. Sci. Biotechnol. 2006, 81, 3–17. [Google Scholar] [CrossRef]
- Zollinger, N.; Kjelgren, R.; Cerny-Koenig, T.; Kopp, K.; Koenig, R. Drought responses of six ornamental herbaceous perennials. Sci. Hortic. 2006, 109, 267–274. [Google Scholar] [CrossRef]
- Mugnai, S.; Ferrante, A.; Petrognani, L.; Serra, G.; Vernieri, P. Stress-induced variation in leaf gas exchange and chlorophyll a fluorescence in Callistemon plants. Res. J. Agric. Biol. Sci. 2009, 4, 913–921. [Google Scholar]
- Navarro, A.; Álvarez, S.; Castillo, M.; Bañón, S.; Sánchez-Blanco, M.J. Changes in tissue-water relations, photosynthetic activity, and growth of Myrtus communis plants in response to different conditions of water availability. J. Hortic. Sci. Biotechnol. 2009, 84, 541–547. [Google Scholar] [CrossRef]
- Maggio, A.; Raimondi, G.; Martino, A.; De Pascale, S. Salt stress response in tomato beyond the salinity tolerance threshold. Environ. Exp. Bot. 2007, 59, 276–282. [Google Scholar] [CrossRef]
- Wu, F.; Bao, W.; Li, F.; Wu, N. Effects of drought stress and N supply on the growth, biomass partitioning and water-use efficiency of Sophora davidii seedlings. Environ. Exp. Bot. 2008, 63, 248–255. [Google Scholar] [CrossRef]
- Smirnoff, N. Plant resistance to environmental stress. Curr. Opin. Biotechnol. 1998, 9, 214–219. [Google Scholar] [CrossRef]
- Davies, M.J.; Harrison-Murray, R.; Atkinson, C.J.; Grant, O.M. Application of deficit irrigation to container-grown hardy ornamental nursery stock via overhead irrigation, compared to drip irrigation. Agric. Water Manag. 2016, 163, 244–254. [Google Scholar] [CrossRef] [Green Version]
- Pollastrini, M.; Desotgiu, R.; Camin, F.; Ziller, L.; Marzuoli, R.; Gerosa, G.; Bussotti, F. Intra-annual pattern of photosynthesis, growth and stable isotope partitioning in a poplar clone subjected to ozone and water stress. Water Air Soil Pollut. 2013, 224, 1761. [Google Scholar] [CrossRef]
- Sánchez-Blanco, M.J.; Ortuño, M.F.; Bañon, S.; Álvarez, S. Deficit irrigation as a strategy to control growth in ornamental plants and enhance their ability to adapt to drought conditions. J. Hortic. Sci. Biotechnol. 2019, 94, 137–150. [Google Scholar] [CrossRef]
- Aguirrezabal, L.; Bouchier-Combaud, S.; Radziejwoski, A.; Dauzat, M.; Cookson, S.J.; Granier, C. Plasticity to soil water deficit in Arabidopsis thaliana: Dissection of leaf development into underlying growth dynamic and cellular variables reveals invisible phenotypes. Plant Cell Environ. 2006, 29, 2216–2227. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K.; Tran, L.S.P. Response of plants to water stress. Front. Plant Sci. 2014, 5, 86. [Google Scholar] [CrossRef] [Green Version]
- Stanton, K.M.; Mickelbart, M.V. Maintenance of water uptake and reduced water loss contribute to water stress tolerance of Spiraea alba Du Roi and Spiraea tomentosa L. Hortic. Res. 2014, 1, 14033. [Google Scholar] [CrossRef]
- Waghorn, M.J.; Whitehead, D.; Watt, M.S.; Mason, E.G.; Harrington, J.J. Growth, biomass, leaf area and water-use efficiency of juvenile Pinus radiata in response to water deficits. N. Z. J. For. Sci. 2015, 45, 3. [Google Scholar] [CrossRef]
- Santana-Vieira, D.D.S.; Freschi, L.; da Hora Almeida, L.A.; De Moraes, D.H.S.; Neves, D.M.; Dos Santos, L.M.; Bertolde, F.Z.; dos Santos Soares Filho, W.; Coelho Filho, M.A.; da Silva Gesteira, A. Survival strategies of citrus rootstocks subjected to drought. Sci. Rep. 2016, 6, 38775. [Google Scholar] [CrossRef]
- Clauw, P.; Coppens, F.; Beuf, K.; Dhondt, S.; Daele, T.V.; Maleux, K.; Storme, V.; Clement, L.; Gonzalez, N.; Inzé, D. Leaf responses to mild drought stress in natural variants of Arabidopsis. Plant Physiol. 2015, 167, 800–816. [Google Scholar] [CrossRef]
- da Cunha Cruz, Y.; Scarpa, A.L.M.; Pereira, M.P.; de Castro, E.M.; Pereira, F.J. Growth of Typha domingensis as related to leaf physiological and anatomical modifications under drought conditions. Acta Physiol. Plant. 2019, 41, 64. [Google Scholar] [CrossRef]
- Cassaniti, C.; Romano, D.; Hop, M.E.C.M.; Flowers, T.J. Growing floricultural crops with brackish water. Environ. Exp. Bot. 2013, 92, 165–175. [Google Scholar] [CrossRef]
- Merwad, A.R.M.; Desoky, E.S.M.; Rady, M.M. Response of water deficit-stressed Vigna unguiculata performances to silicon, proline or methionine foliar application. Sci. Hortic. 2018, 228, 132–144. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Sankar, B.; Murali, P.V.; Gomathinayagam, M.; Lakshmanan, G.M.A.; Panneerselvam, R. Water deficit stress effects on reactive oxygen metabolism in Catharanthus roseus: Impacts on ajmalicine accumulation. Colloids Surf. B Biointerfaces 2008, 62, 105–111. [Google Scholar] [CrossRef]
- Kiani, S.P.; Maury, P.; Sarrafi, A.; Grieu, P. QTL analysis of chlorophyll fluorescence parameters in sunflower (Helianthus annuus L.) under well-watered and water-stressed conditions. Plant Sci. 2008, 175, 565–573. [Google Scholar] [CrossRef]
- Tahkokorpi, M.; Taulavuori, K.; Laine, K.; Taulavuori, E. After-effects of drought-related winter stress in previous and current year stems of Vaccinium myrtillus L. Environ. Exp. Bot. 2007, 61, 85–93. [Google Scholar] [CrossRef]
- Jiang, Y.; Huang, B. Drought and heat stress injury to two cool-season turfgrass in relation to antioxidant metabolism and lipid peroxidation. Crop Sci. 2001, 41, 436–442. [Google Scholar] [CrossRef]
- García-Valenzuela, X.; García-Moya, E.; Rascón-Cruz, Q.; Herrera-Estrella, L.; Aguado-Santacruz, G.A. Chlorophyll accumulation is enhanced by osmotic stress in graminaceous chlorophyllic cells. J. Plant Physiol. 2005, 162, 650–661. [Google Scholar] [CrossRef]
- Li, S.; Pezeshki, S.R.; Goodwin, S. Effects of soil moisture regimes on photosynthesis and growth in cattail (Typha latifolia). Acta Oecol. 2004, 25, 17–22. [Google Scholar] [CrossRef]
- Yamashita, N.; Koike, N.; Ishida, A. Leaf ontogenetic dependence of light acclimation in invasive and native subtropical trees of different successional status. Plant Cell Environ. 2002, 25, 1341–1356. [Google Scholar] [CrossRef]
- Souza, P.U.; Lima, L.K.S.; Soares, T.L.; de Jesus, O.N.; Coelho Filho, M.A.; Girardi, E.A. Biometric, physiological and anatomical responses of Passiflora spp. to controlled water deficit. Sci. Hortic. 2018, 229, 77–90. [Google Scholar] [CrossRef]
- Guerfel, M.; Baccouri, O.; Boujnah, D.; Chaïbi, W.; Zarrouk, M. Impacts of water stress on gas exchange, water relations, chlorophyll content and leaf structure in the two main Tunisian olive (Olea europaea L.) cultivars. Sci. Hortic. 2009, 119, 257–263. [Google Scholar] [CrossRef]
- Lee, J.S. The relationship between stomatal opening and photosynthetic activity of the mesophyll in Commelina communis L. J. Environ. Sci. Int. 2006, 15, 1109–1117. [Google Scholar] [CrossRef]
- Mathobo, R.; Marais, D.; Steyn, J.M. The effect of drought stress on yield, leaf gaseous exchange and chlorophyll fluorescence of dry beans (Phaseolus vulgaris L.). Agric. Water Manag. 2017, 180, 118–125. [Google Scholar] [CrossRef]
- Monclus, R.; Dreyer, E.; Villar, M.; Delmotte, F.M.; Delay, D.; Petit, J.M.; Barbaroux, C.; Le Thiec, D.; Bréchet, C.; Brignolas, F. Impact of drought on productivity and water use efficiency in 29 genotypes of Populus deltoids × Populus nigra. New Phytol. 2006, 169, 765–777. [Google Scholar] [CrossRef]
- Deuner, S.; Zanandrea, I.; Silveira, N.M.; Mesquita, A.C.; Goulart, P.F.P.; Alves, J.D. Stomatal behavior and components of the antioxidative system in coffee plants under water stress. Sci. Agric. 2011, 68, 77–85. [Google Scholar] [CrossRef]
- Morales, F.; Abadía, A.; AbadÞa, J. Photoinhibition and Photoprotection under Nutrient Deficiencies, Drought and Salinity. In Photoprotection, Photoinhibition, Gene Regulation, and Environment; Advances in Photosynthesis and, Respiration; Demmig-Adams, B., Adams, W.W., Mattoo, A.K., Eds.; Springer: Dordrecht, The Netherlands, 2008; Volume 21, pp. 65–85. [Google Scholar]
- Hessini, K.; Ghandour, M.; Albouchi, A.; Soltani, A.; Werner, K.H.; Abdelly, C. Biomass production, photosynthesis, and leaf water relations of Spartina alterniflora under moderate water stress. J. Plant Res. 2008, 121, 311–318. [Google Scholar] [CrossRef]
- Ferrar, P.J.; Osmond, C.B. Nitrogen supply as a factor influencing photoinhibition and photosynthetic acclimation after transfer of shade-grown Solanum dulcamara to bright light. Planta 1986, 168, 563–570. [Google Scholar] [CrossRef]
- Osmond, C.B.; Anderson, J.M.; Ball, M.C.; Egerton, J.J.G. Compromising efficiency: The molecular ecology of light-resource utilization in plants. In Physiological Plant Ecology; Scholes, J.D., Barker, M.G., Eds.; Blackwell Science: Oxford, UK, 1999. [Google Scholar]
- Osmond, C.B. What Is Photoinhibition? Some Insights from Comparisons of Shade and Sun Plants. In Photoinhibition of Photosynthesis-From Molecular Mechanisms to the Field; Baker, N.R., Bowyer, J.R., Eds.; BIOS Scientific Publishers: Oxford, UK, 1994; pp. 1–24. [Google Scholar]
- Murata, N.; Takahashi, S.; Nishiyama, Y.; Allakhverdiev, S.I. Photoinhibition of photosystem II under environmental stress. Biochim. Biophys. Acta 2007, 1767, 414–421. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Treatments | Total Dry Biomass (g plant−1) | Shoot Dry Biomass (g plant−1) | Root Dry Biomass (g plant−1) | R/S Ratio (g g−1) | Leaf (n. plant−1) | Total Leaf Area (cm2 plant−1) | Unit Leaf Area (cm2 plant−1) | SLA (cm2 g−1) | SPAD Index |
|---|---|---|---|---|---|---|---|---|---|---|
| Polygala | 146.7 ± 8.6 b | 110.8 ± 9.6 | 35.9 ± 5.1 b | 0.31 ± 0.0 b | 4443.6 ± 507.9 a | 7304.6 ± 779.9 a | 1.7 ± 0.1 b | 99.8 ± 3.1 a | 53.7 ± 0.9 b | |
| Viburnum | 167.2 ± 9.5 a | 109.4 ± 6.8 | 57.9 ± 4.2 a | 0.54 ± 0.0 a | 117.8 ± 14.5 b | 3408.7 ± 393.9 b | 29.5 ± 0.7 a | 56.3 ± 2.3 b | 73.8 ± 1.5 a | |
| 40% WC | 192.2 ± 5.8 a | 136.4 ± 2.7 a | 55.8 ± 6.2 a | 0.41 ± 0.0 | 3170.0 ± 578.7 a | 7488.9 ± 1131.1 a | 15.3 ± 6.1 | 88.1 ± 5.7 a | 60.4 ± 2.4 b | |
| 30% WC | 193.9 ± 4.9 a | 135.3 ± 5.3 a | 58.6 ± 3.0 a | 0.44 ± 0.0 | 2949.8 ± 670.9 a | 6952.7 ± 1214.2 a | 15.4 ± 6.1 | 82.0 ± 4.5 ab | 62.7 ± 2.6 b | |
| 20% WC | 134.0 ± 12.9 b | 92.4 ± 3.4 b | 43.6 ± 4.8 ab | 0.46 ± 0.1 | 1713.9 ± 533.4 b | 3971.9 ± 680.2 b | 15.3 ± 6.2 | 74.8 ± 4.2 b | 63.3 ± 2.6 b | |
| 10% WC | 105.6 ± 8.0 c | 76.2 ± 2.3 c | 29.5 ± 6.2 b | 0.32 ± 0.1 | 1289.0 ± 560.6 b | 3013.3 ± 527.4 c | 16.4 ± 6.6 | 67.0 ± 3.7 c | 68.6 ± 1.1 a | |
| Significance 1 | ||||||||||
| Species (S) | ** | ns | *** | *** | *** | *** | *** | *** | *** | |
| Treatments (T) | *** | *** | *** | ns | *** | *** | ns | *** | ** | |
| S × T | ** | * | * | ** | *** | *** | ns | ns | ns |
| Species | Treatments | Thickness (µm) | Stomata | |||
|---|---|---|---|---|---|---|
| Total | Palisade | Spongy | Density (n/mm2) | Size (µm) | ||
| Polygala | 284.7 ± 10.0 b | 69.9 ± 2.5 b | 170.3 ± 6.2 b | 175.8 ± 2.5 b | 2.9 ± 0.0 b | |
| Viburnum | 432.4 ± 14.8 a | 146.1 ± 11.3 a | 228.7 ± 9.9 a | 204.0 ± 5.6 a | 2.6 ±0.1 a | |
| 40% WC | 331.6 ± 15.6 b | 81.0 ± 8.2 c | 203.2 ± 15.7 | 199.5 ± 9.4 ab | 2.8 ± 0.1 a | |
| 30% WC | 331.5 ± 41.7 b | 103.4 ± 21.0 b | 184.2 ± 20.1 | 172.5 ± 4.8 c | 2.9 ± 0.1 a | |
| 20% WC | 384.5 ± 45.7 a | 117.8 ± 20.7 ab | 211.1 ± 22.1 | 185.3 ± 3.0 bc | 2.9 ± 0.0 a | |
| 10% WC | 386.7 ± 33.3 a | 129.8 ± 23.6 a | 199.4 ± 8.2 | 202.3 ± 7.0 a | 2.5 ± 0.1 b | |
| Significance † | ||||||
| Species (S) | *** | *** | ns | *** | *** | |
| Treatments (T) | *** | *** | *** | *** | *** | |
| S × T † | *** | *** | * | *** | *** | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tribulato, A.; Toscano, S.; Di Lorenzo, V.; Romano, D. Effects of Water Stress on Gas Exchange, Water Relations and Leaf Structure in Two Ornamental Shrubs in the Mediterranean Area. Agronomy 2019, 9, 381. https://doi.org/10.3390/agronomy9070381
Tribulato A, Toscano S, Di Lorenzo V, Romano D. Effects of Water Stress on Gas Exchange, Water Relations and Leaf Structure in Two Ornamental Shrubs in the Mediterranean Area. Agronomy. 2019; 9(7):381. https://doi.org/10.3390/agronomy9070381
Chicago/Turabian StyleTribulato, Alessandro, Stefania Toscano, Virginia Di Lorenzo, and Daniela Romano. 2019. "Effects of Water Stress on Gas Exchange, Water Relations and Leaf Structure in Two Ornamental Shrubs in the Mediterranean Area" Agronomy 9, no. 7: 381. https://doi.org/10.3390/agronomy9070381
APA StyleTribulato, A., Toscano, S., Di Lorenzo, V., & Romano, D. (2019). Effects of Water Stress on Gas Exchange, Water Relations and Leaf Structure in Two Ornamental Shrubs in the Mediterranean Area. Agronomy, 9(7), 381. https://doi.org/10.3390/agronomy9070381