Effects of Nitrogen Fertilization and Seed Piece Applied Fungicides on Establishment, Tiller Dynamics, and Sucrose Yields in Successively Planted Sugarcane

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Sites Characteristics
2.2. Nitrogen Treatments
2.3. Fungicide Treatments
2.4. Tissue Analyses
2.5. Tiller Dynamics
2.6. Yield Measurements
2.7. Statistical Analyses
3. Results
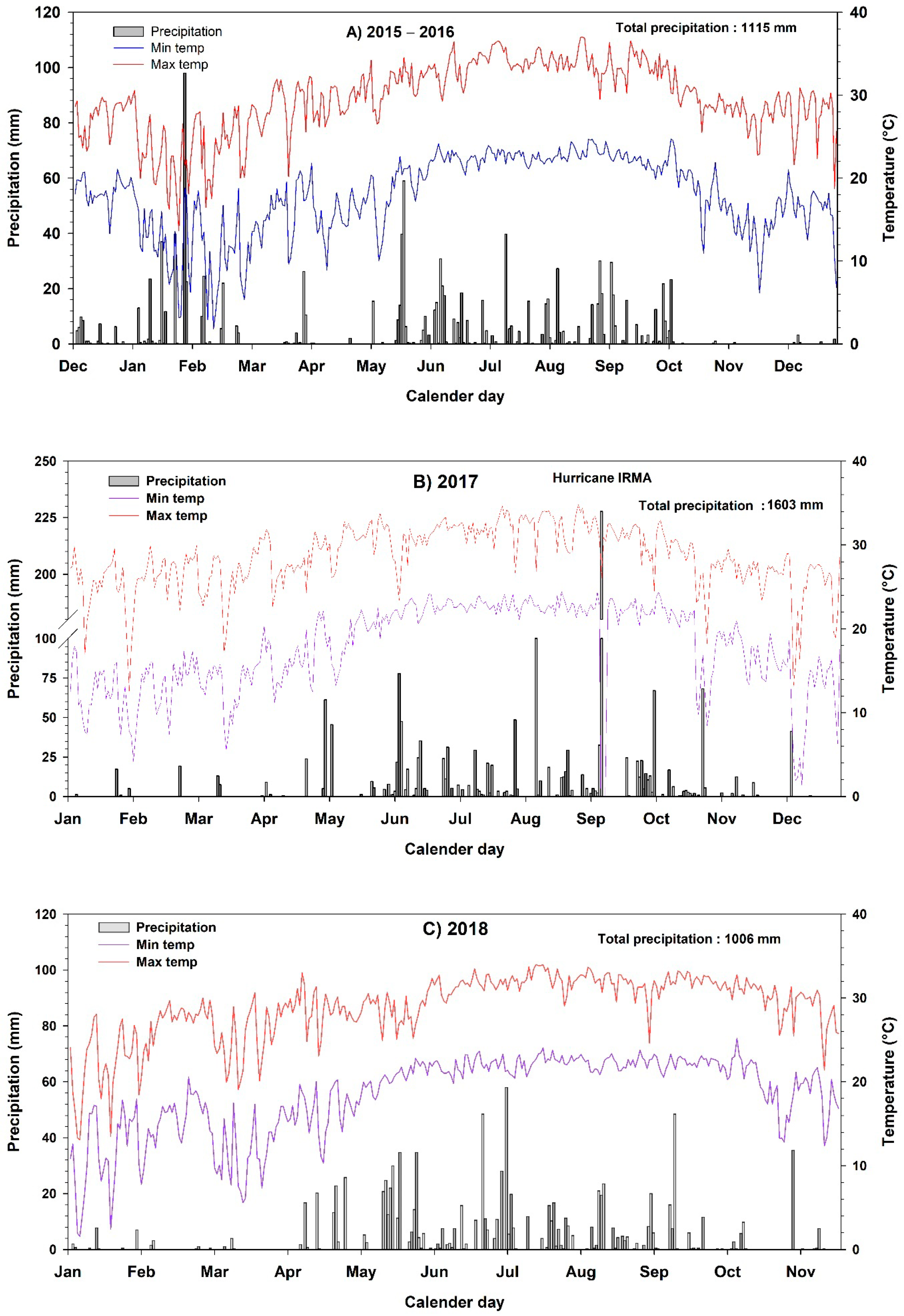
3.1. Weather Conditions
3.2. Sugarcane Establishment and Tiller Dynamics
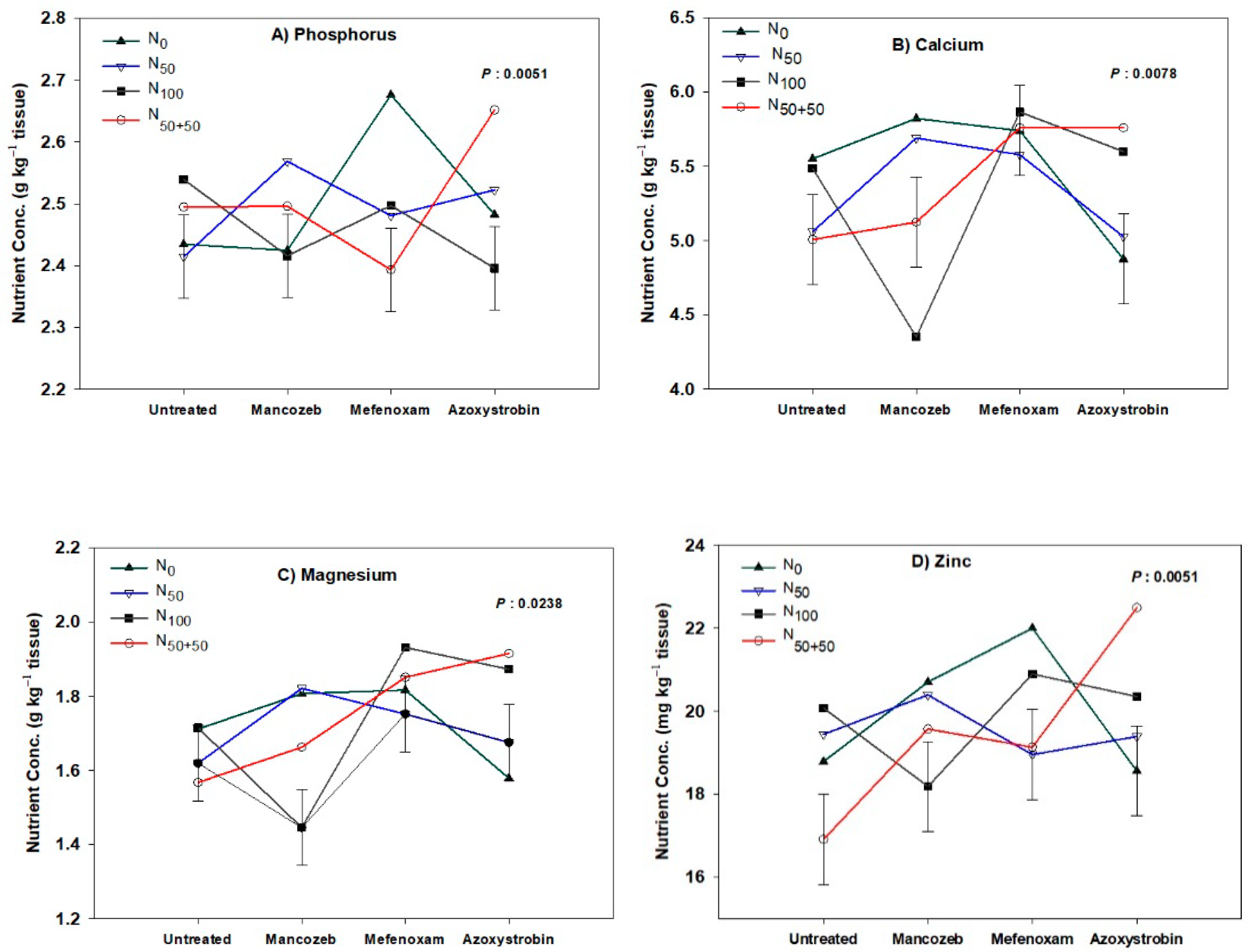
3.3. Tissue Nutrient Concentrations
3.3.1. Plant Crop
3.3.2. Ratoon Crops
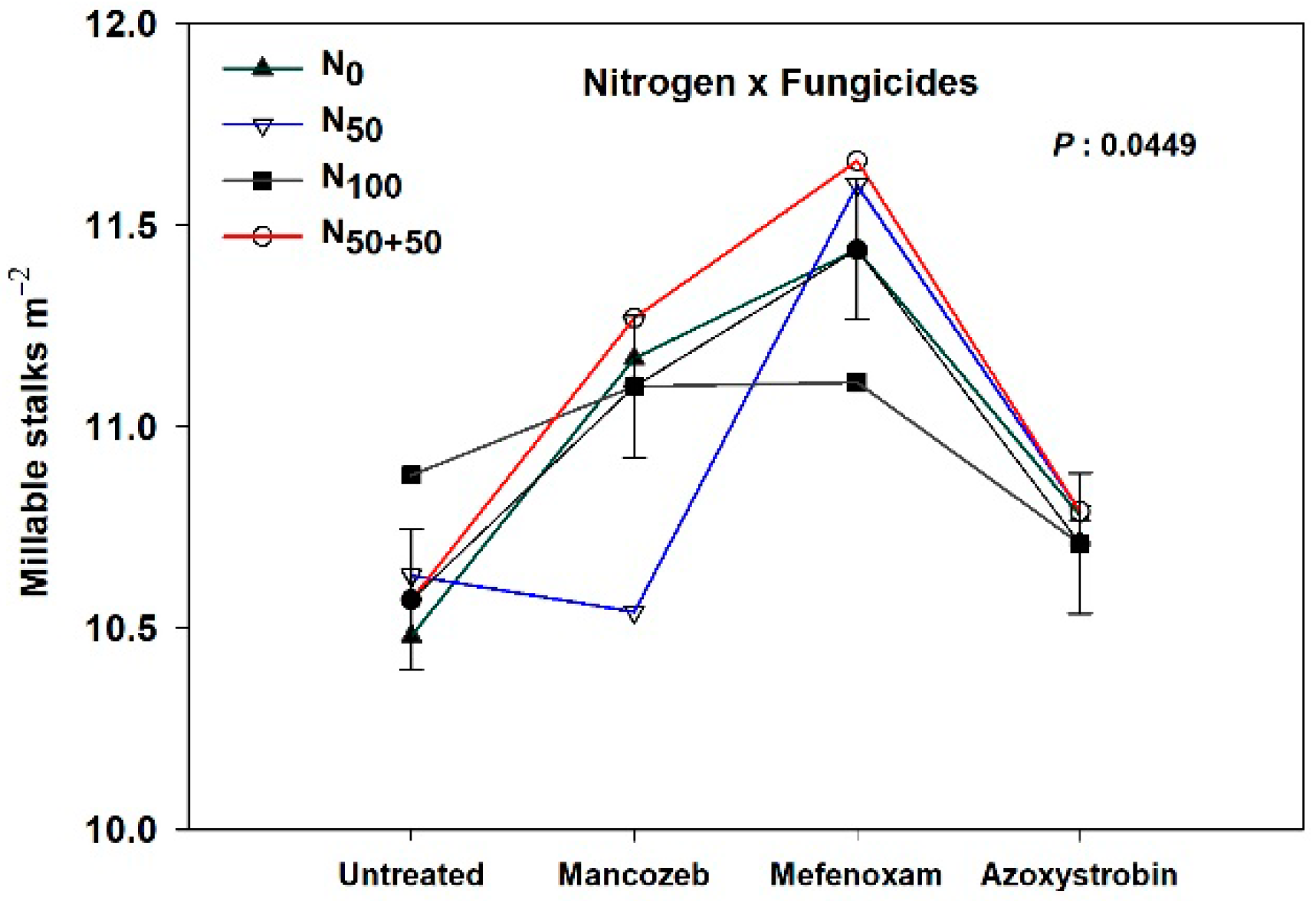
3.4. Yield Attributes and Sucrose Yields
3.4.1. Plant Crops
3.4.2. Ratoon Crops
3.4.3. Average Yields
4. Discussion
4.1. Nitrogen Effects on Sugarcane Tiller Dynamics and Crop Yields
4.2. Fungicidal Effects on Sugarcane Tiller Dynamics and Crop Yields
4.3. Effects on Tissue Nutrient Concentrations
4.4. Economic Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Hodges, A.W.; Rahmani, M.; Stevens, T.J. Economic Contributions of Agriculture, Natural Resources, and Related Industries in Florida for 2010. Available online: http://www.fred.ifas.ufl.edu/economic-impact- (accessed on 14 October 2018).
- Rice, R.L.; Baucum, L.; Davidson, W. Sugarcane Variety Census: Florida 2013. Sugar J. 2014, 76, 10–19. [Google Scholar]
- Glaz, B.; Ulloa, M.F. Fallow and successive planting effects on sugarcane yields in Florida. J.Am.Soc.Sugar Cane. Technol. 1995, 15, 41–53. [Google Scholar]
- Sandhu, H.S. Yield decline under sugarcane monoculture in Histosols and its potential causes. In Proceedings of the 75th Annual Convention and Sugar Expo, The Sugarcane Technoloists of Association of India, Kochi, Kerala, India, 3–5 August 2017; pp. 224–230. [Google Scholar]
- Garside, A.L.; Bramley, R.G.V.; Bristow, K.L.; Holt, J.A.; Magarey, R.C.; Nable, R.O.; Pankhurst, C.E.; Skjemstad, J.O. Garside 1997-Comparisons between paired old and new land sites for sugarcane growth and yield and soil chemical, physical and biological properties. In Proceedings of the Australian Society of Sugar Cane Technologists, Cairns, Queensland, Australia, 29 April–2 May 1997; pp. 60–66. [Google Scholar]
- Bennett, A.J.; Bending, G.D.; Chandler, D.; Hilton, S.; Mills, P. Meeting the demand for crop production: the challenge of yield decline in crops grown in short rotations. Biol. Rev. 2012, 87, 52–71. [Google Scholar] [CrossRef] [PubMed]
- Magarey, R.C. Microbiological aspects of sugarcane yield decline. Aust. J. Agric. Res. 1996, 47, 307–322. [Google Scholar] [CrossRef]
- Garside, A.; Bell, M.; Magary, R. Effect of breaks from sugarcane monoculture on growth and yield of subsequent sugarcane crops. Proc. South African Sugarcane Technol. 2002, 74, 12–18. Available online: https://pdfs.semanticscholar.org/b3fe/dbfbb697394013e59e196847a24db6146b2e.pdf (accessed on 14 October 2018).
- Magarey, R.C.; Grace, D.J. The relationship between root parameters, soil treatment and sugarcane shoot growth in glasshouse experiments. Proc. Aust. Soc. Sugarcane Technol. 1997, 19, 87–92. Available online: http://hdl.handle.net/11079/14333 (accessed on 10 March 2018).
- Hoy, J.W.; Schneider, R.W. Role of Pythium in sugarcane stubble decline: Pathogenicity and virulence of Pythium species. Phytopathology 1988, 78, 1688–1692. [Google Scholar] [CrossRef]
- Raid, R.N.; Rott, P. Sugarcane Pineapple Disease or Sugarcane Pineapple Set Rot. Available online: http://edis.ifas.ufl.edu/sc005 (accessed on 10 March 2018).
- Raid, R.N. Fungicidal control of pineapple disease of sugarcane. J. Am. Soc. Sugar Cane Technol. 1990, 10, 45–50. [Google Scholar]
- Snyder, G.H. Soils of EAA. In Everglades Agricultural Area (EAA): Water, Soil, Crop, and Environmental Management; Bottcher, A.B., Izuno, F.T., Eds.; University of Florida Press: Gainesville, FL, USA, 1994; pp. 27–41. [Google Scholar]
- Mccray, J.M.; Sandhu, H.S.; Rice, R.W.; Odero, D.C. Nutrient Requirements for Sugarcane Production on Florida Muck Soils. Available online: http://edis.ifas.ufl.edu/sc076 (accessed on 18 August 2018).
- Shih, S.F.; Mishoe, J.W.; Jones, J.W.; Myhre, D.L. Modeling the subsidence of Everglades organic soil. Trans. ASAE 1978, 21, 1151–1156. [Google Scholar] [CrossRef]
- Terry, R.E. Nitrogen Mineralization in Florida Histosols1. Soil Sci. Soc. Am. J. 1980, 44, 747. [Google Scholar] [CrossRef]
- Shih, S.F.; Glaz, B.; Barnes, R.E., Jr. Subsidence of organic soils in the Everglades Agricultural Area during the past 19 years. Annu. Proc. Soil Crop Sci. Soc. Fl. 1998, 57, 20–29. [Google Scholar]
- Snyder, G.H. Everglades Agricultural Area soil subsidence and land use projections. Soil Crop Sci. Soc. Fl. Proc. 2005, 64, 44–51. [Google Scholar]
- Fontaine, S.; Henault, C.; Aamor, A.; Bdioui, N.; Bloor, J.M.G.M.G.; Maire, V.; Mary, B.; Revaillot, S.; Maron, P.A.A. Fungi mediate long term sequestration of carbon and nitrogen in soil through their priming effect. Soil Biol. Biochem. 2011, 43, 86–96. [Google Scholar] [CrossRef]
- Robertson, F.A.; Thorburn, P.J. Management of sugarcane harvest residues: Consequences for soil carbon and nitrogen. Aust. J. Soil Res. 2007, 45, 13–23. [Google Scholar] [CrossRef]
- Sall, S.N.; Masse, D.; Bernhard-Reversat, F.; Guisse, A.; Chotte, J.-L. Microbial activities during the early stage of laboratory decomposition of tropical leaf litters: The effect of interactions between litter quality and exogenous inorganic nitrogen. Biol. Fertil. Soils 2003, 39, 103–111. [Google Scholar] [CrossRef]
- Meyer, J.; Rein, P.; Turner, P.; Mathias, K.; McGregor, C. Good Management Practices Manual for the Cane Sugar Industry (Final). Int. Financ. Corp. 2011, 696. [Google Scholar]
- Glaz, B.; Reed, S.T.; Albano, J.P. Sugarcane Response to Nitrogen Fertilization on a Histosol with Shallow Water Table and Periodic Flooding. J. Agron. Crop Sci. 2008, 194, 369–379. [Google Scholar] [CrossRef]
- Jennewein, S.P. Determining management effcts on soil gas flux, microbial biomass, nutrient cycling, and crop yield to identify sustainable practices in the Everglades Agricultural Area. Ph.D. Thesis, University of Florida, Gainesville, FL, USA, 2017. Available online: http://ufdc.ufl.edu/UFE0051277/00001 (accessed on 15 October 2018).
- Rice, R.W.; Gilbert, R.A.; Daroub, S.H. Application of the soil taxonomy key to the organic soils of the Everglades Agricultural Area. Univ. Florida Doc. SS-AGR-246: 10. Available online: http://ufdcimages.uflib.ufl.edu/IR/00/00/15/82/00001/AG15100.pdf (accessed on 20 May 2018).
- Murphy, J.; Riley, J.P.J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Snyder, G.H. Development of a Silicon Soil Test for Histosol-Grown Rice; Belle Glade EREC Research Report EV; Agriculture Research Education Center, University of Florida: Gainesville, FL, USA, 1991; pp. 29–39. [Google Scholar]
- Edme, S.J.; Tai, P.Y.P.; Glaz, B.; Gilbert, R.A.; Miller, J.D.; Davidson, J.O.; Dunckelman, J.W.; Comstock, J.C. Registration of ‘CP 96-1252’ Sugarcane. Crop Sci. 2005, 45, 423. [Google Scholar] [CrossRef]
- Vuyyuru, M.; Sandhu, H.S.; McCray, J.M.; Raid, R.N. Effects of Soil-Applied Fungicides on Sugarcane Root and Shoot Growth, Rhizosphere Microbial Communities, and Nutrient Uptake. Agronomy 2018, 8, 223. [Google Scholar] [CrossRef]
- USEPA. Methods for the Determination of Inorganic Substances in Environmental Samples, USEPA Office of Research and Development, 8/93. Cincinnati, OH, EPA 600/R-93/100; United States; 1993. Available online: http://monitoringprotocols.pbworks.com/f/EPA600-R-63-100.pdf (accessed on 15 October 2018).
- USEPA. Methods for the Determination of Metals in Environmental Samples, USEPA Office of Research and Development, Supp. 1, 5/94. Cincinnati, OH, EPA 600/R-94/111; William Andrew: Norwich, UK, 2014; ISBN 9780815513988.
- Anderson, D.; Bowen, J. Sugarcane Nutrition; Potash and Phosphate Institute: Atlanta, GA, USA, 1990. [Google Scholar]
- McCray, J.; Ji, S.; Powell, G.; Montes, G.; Perdomo, R.; Luo, Y. Seasonal concentrations of leaf nutrients in Florida sugarcane. Sugar Cane Int. 2009, 27, 17–24. [Google Scholar]
- Elwali, A.M.O.; Gascho, G.J. Soil Testing, Foliar Analysis, and DRIS as Guides for Sugarcane Fertilization1. Agron. J. 1984, 76, 466. [Google Scholar] [CrossRef]
- Legendre, B.L. The core/press method for predicting the sugar yield from cane for use in cane payment. Sugar J 1992, 54, 2–7. [Google Scholar]
- Littell, R.C.; Henry, P.R.; Ammerman, C.B. Statistical analysis of repeated measures data using SAS procedures. J. Anim. Sci. 1998, 76, 1216–1231. [Google Scholar] [CrossRef] [PubMed]
- Bailey, R.G. Description of the Ecoregions of the United States; U.S. Department of Agriculture, Miscellaneous Publication: Washington, DC, USA, 1980; p. 1391. [Google Scholar]
- FAWN—Florida Automated Weather Network. Available online: https://fawn.ifas.ufl.edu/data/reports/ (accessed on 10 August 2018).
- Mccray, J.M.; Mylavarapu, R. Sugarcane Nutrient Management Using Leaf Analysis. IFAS Ext. 2013. Available online: https://edis.ifas.ufl.edu/ag345 (accessed on 10 March 2017).
- Porter, P.S.; Sanchez, C. Nitrogen in the organic soils of the Everglades Agricultural Area. In Everglades Agricultural Area (EAA): Water, Soil, Crop, and Environmental Management; Bottcher, A.B., Izuno, F.T., Eds.; University of Florida: Gainesville, FL, USA, 1992; p. 318. [Google Scholar]
- McCray, J.M.; Rice, R.W.; Luo, Y.; Ji, S. Phosphorus Fertilizer Calibration for Sugarcane on Everglades Histosols. Commun. Soil Sci. Plant Anal. 2012, 43, 2691–2707. [Google Scholar] [CrossRef]
- Wright, A.L.; Inglett, P.W. Soil Organic Carbon and Nitrogen and Distribution of Carbon-13 and Nitrogen-15 in Aggregates of Everglades Histosols. Soil Sci. Soc. Am. J. 2009, 73, 427. [Google Scholar] [CrossRef]
- Hanlon, E.A.; Anderson, D.L.; Diaz, O.A. Nitrogen mineralizations in histosols of the everglades agricultural area. Commun. Soil Sci. Plant Anal. 2008, 28, 73–87. [Google Scholar] [CrossRef]
- Hochmuth, G.; Hanlon, E.; Snyder, G.; Nagata, R.; Schueneman, T. Fertilization of Sweet Corn, Celery, Romaine, Escarole, Endive, and Radish on Organic Soils in Florida. Available online: http://edis.ifas.ufl.edu (accessed on 11 Feburay 2018).
- Wright, A.L.; Odero, D.C. Nitrogen Cycling and Management for Romaine and Crisphead Lettuce Grown on Organic Soils. Available online: http://www.ipni.net/article/IPNI-3153 (accessed on 13 November 2018).
- Vasantha, S.; Gupta, C.; Shekinah, D.E. Physiological studies on tiller production and its senescence in sugarcane—Response comparison between plant and ratoon crops. Indian J. Agric. Sci. 2014, 84, 24–27. [Google Scholar]
- Lofton, J.; Tubaña, B. Effect of Nitrogen Rates and Application Time on Sugarcane Yield and Quality. J. Plant Nutr. 2015, 38, 161–176. [Google Scholar] [CrossRef]
- Bell, M.J.; Garside, A.L. Growth and yield responses to amending the sugarcane monoculture: interactions between break history and nitrogen fertiliser. Crop Pasture Sci. 2014, 65, 287–299. [Google Scholar] [CrossRef] [Green Version]
- Muchovej, R.M.; Newman, P.R. Nitrogen fertilization of sugarcane on a sandy soil: I. Yield and leaf nutrient composition. J. Am. Soc. Sugar Cane Technol. 2004, 24, 210–224. Available online: http://www.assct.org/journal/JASSCT PDF Files/volume 24/A03-10 Muchovej final.pdf (accessed on 10 August 2018).
- McCray, J.M.; Morgan, K.T.; Baucum, L.; Ji, S. Sugarcane Yield Response to Nitrogen on Sand Soils. Agron. J. 2014, 106, 1461. [Google Scholar] [CrossRef]
- Garside, A.L.; Bell, M.J.; Magarey, R.C. Monoculture Yield Decline—Fact Not Fiction. Proc. Int. Soc. Sugar Cane Technol. 2001, 24, 16–21. [Google Scholar]
- Pankhurst, C.E.; Blair, B.L.; Magarey, R.C.; Stirling, G.R.; Garside, A.L. Effects of biocides and rotation breaks on soil organisms associated with the poor early growth of sugarcane in continuous monoculture. Plant Soil 2005, 268, 255–269. [Google Scholar] [CrossRef]
- Bending, G.D.; Rodríguez-Cruz, M.S.; Lincoln, S.D. Fungicide impacts on microbial communities in soils with contrasting management histories. Chemosphere 2007, 69, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Niu, J.; Zhang, W.; Chen, X.; Li, C.; Yuan, L.; Xie, J. Potassium nutrition of crops under varied regimes of nitrogen supply. Plant Soil 2010, 335, 21–34. [Google Scholar] [CrossRef]
- Fageria, V.D. Nutrient interactions in crop plants. J. Plant Nutr. 2001, 24, 1269–1290. [Google Scholar] [CrossRef]
- Malhi, S.S.; Nyborg, M.; Harapiak, J.T.; Heaney, D.J. Long-term effects of ammonium nitrate application on soil pH, and micronutrients in soil and bromegrass hay. In Plant-Soil Interactions at Low pH: Principles and Management; Springer: Dordrecht, The Netherlands, 1995; pp. 105–110. [Google Scholar]
- Kaler, A.S.; McCray, J.M.; Wright, A.L.; Erickson, J.E. Sugarcane yield and plant nutrient response to sulfur-amended Everglades histosols. J. Plant Nutr. 2017, 40, 187–196. [Google Scholar] [CrossRef]
- Robinson, N.; Brackin, R.; Vinall, K.; Soper, F.; Holst, J.; Gamage, H.; Paungfoo-Lonhienne, C.; Rennenberg, H.; Lakshmanan, P.; Schmidt, S. Nitrate Paradigm Does Not Hold Up for Sugarcane. PLoS ONE 2011, 6, e19045. [Google Scholar] [CrossRef]
- USDA ERS Fertilizer Use and Price. Available online: https://www.ers.usda.gov/data-products/fertilizer-use-and-price.aspx (accessed on 10 October 2018).
- Roka, F.M.; Baucum, L.E.; Alvarez, J. Costs and Returns for Sugarcane Production on Muck Soils in Southern Florida 2008–2009. Available online: http://edis.ifas.ufl.edu (accessed on 10 October 2018).
- USDA ERS Sugar and Sweeteners Yearbook Tables. Available online: https://www.ers.usda.gov/data-products/sugar-and-sweeteners-yearbook-tables.aspx (accesed on 20 November 2018).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site 1 | Site 2 | |
|---|---|---|
| Location | 26°39′53″ N 80°38′14″ W | 26°65′96″ N 80°63′10″ W |
| Soil thickness (cm) | 26 | 28 |
| pH | 7.4 | 7.6 |
| Total carbon (g kg−1) | 428.15 | 394.29 |
| Total nitrogen (g kg−1) | 23.07 | 24.23 |
| C:N ratio | 18.58 | 16.27 |
| Mehlich-3 phosphorus (mg kg−1) | 40 | 22 |
| Potassium (mg kg−1) | 108 | 119 |
| Calcium (mg kg−1) | 9438 | 10,106 |
| Magnesium (mg kg−1) | 1335 | 1344 |
| Silicon (mg kg−1) | 50 | 39 |
| Crop Calendar | ||
| Sugarcane stubble crop harvest | 15 November 2015 | 22 October 2016 |
| Sugarcane planting | 10 December 2015 | 18 November 2016 |
| Plow-out to replant duration | 25 days | 27 days |
| Plant crop sampling | 10 December 2016 (12 Months) | 15 November 2016 (12 Months) |
| Plant crop machine harvest | 05 March 2017 | 17 December 2017 |
| Ratoon crop sampling | 19 December 2017 (9 months, 15 days) | 19 November 2018 (11 months, 2 days) |
| Fixed Effects | N | P | K | Ca | Mg | Fe | Cu | Zn | Mn | NBI |
|---|---|---|---|---|---|---|---|---|---|---|
| Plant crop 2016 | ||||||||||
| Nitrogen (N) | 0.4879 | 0.5836 | 0.7305 | 0.8379 | 0.9707 | 0.8486 | 0.9693 | 0.8971 | 0.8132 | 0.6094 |
| Fungicide (F) | 0.7359 | 0.7596 | 0.8570 | 0.0835 | 0.0447 | 0.8567 | 0.8738 | 0.2139 | 0.5661 | 0.0610 |
| N × F | 0.9057 | 0.0051 | 0.3963 | 0.0078 | 0.0238 | 0.8255 | 0.2769 | 0.0237 | 0.0876 | 0.7388 |
| Month (M) | 0.0044 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.1670 | 0.2856 | <0.0001 | 0.1939 |
| N × M | 0.3480 | 0.8649 | 0.6978 | 0.3737 | 0.4212 | 0.5646 | 0.9556 | 0.9985 | 0.1405 | 0.4705 |
| N × F × M | 0.0317 | 0.6764 | 0.4719 | 0.2116 | 0.6076 | 0.8580 | 0.9958 | 0.3790 | 0.5748 | 0.8639 |
| Month | N | P | K | Ca | Mg | Fe | Cu | Zn | Mn | NBI |
|---|---|---|---|---|---|---|---|---|---|---|
| g kg−1 | mg kg−1 | |||||||||
| April 2017 | 24.12 a | 2.56 a | 10.91 b | 5.09 a | 1.68 b | 77.79 a | 3.73 a | 20.82 a | 15.09 b | 99.05 a |
| May 2017 | 23.03 b | 2.53 a | 11.05 b | 6.17 a | 1.68 b | 80.93 a | 3.73 a | 19.38 a | 25.62 a | 91.72 a |
| June 2017 | 23.31 ab | 2.36 b | 12.12 a | 4.90 a | 1.83 a | 69.95 b | 3.52 a | 19.75 a | 26.46 a | 89.14 a |
| Fixed Effects | N | P | K | Ca | Mg | Fe | Cu | Zn | Mn | NBI |
|---|---|---|---|---|---|---|---|---|---|---|
| Ratoon crop 2016 | ||||||||||
| Nitrogen (N) | 0.0394 | 0.5356 | 0.4445 | 0.0376 | 0.7050 | 0.2111 | 0.6541 | 0.9006 | 0.5941 | 0.0347 |
| Month (M) | 0.0064 | <0.0001 | <0.0001 | <0.0001 | 0.0016 | <0.0001 | <0.0001 | 0.3149 | 0.1753 | 0.6225 |
| N × M | 0.4427 | 0.5323 | 0.2029 | 0.6689 | 0.3739 | 0.3296 | 0.4168 | 0.1403 | 0.6337 | 0.5053 |
| Ratoon crop 2017 | ||||||||||
| Nitrogen (N) | 0.8993 | 0.4838 | 0.7590 | 0.0352 | 0.0223 | 0.7859 | 0.2115 | 0.0237 | 0.6762 | 0.0418 |
| Month (M) | 0.3007 | <0.0001 | 0.0024 | <0.0001 | 0.1872 | 0.5827 | 0.0098 | 0.0360 | 0.0002 | 0.1001 |
| N × M | 0.0543 | 0.0984 | 0.4709 | 0.8468 | 0.7066 | 0.5313 | 0.0689 | 0.0595 | 0.3876 | 0.1688 |
| Nitrogen | N g kg−1 | Ca g kg−1 | NBI | Ca g kg−1 | Mg g kg−1 | Zn mg kg−1 | NBI |
|---|---|---|---|---|---|---|---|
| Ratoon crop 2016 | Ratoon crop 2017 | ||||||
| N0 | 23.94 b | 4.31 a | 207.12 a | 4.03 ab | 1.14 ab | 10.40 ab | 152.09 a |
| N50 | 25.50 a | 3.78 b | 173.15 ab | 3.72 b | 1.04 b | 9.94 b | 112.06 ab |
| N100 | 24.77 ab | 4.05 ab | 124.84 b | 4.32 a | 1.27 a | 11.09 ab | 95.32 ab |
| N50+50 | 25.04 ab | 4.27 ab | 134.56 b | 4.26 ab | 1.23 ab | 12.05 a | 82.43 b |
| Month | N | P | K | Ca | Mg | Fe | Cu | Zn | Mn | NBI |
|---|---|---|---|---|---|---|---|---|---|---|
| g kg−1 | mg kg−1 | |||||||||
| 2016 Ratoon crop | ||||||||||
| May 2017 | 23.80 b | 1.83 b | 8.55 b | 4.38 a | 1.21 a | 44.37 b | 2.08 b | 11.93 a | 11.93 a | 168.80 a |
| June 2017 | 26.09 a | 1.63 c | 7.50 c | 4.57 a | 1.24 a | 44.90 b | 2.48 a | 12.51 a | 12.51 a | 163.36 a |
| July 2017 | 24.50 ab | 2.01 a | 11.82 a | 3.35 b | 1.10 b | 53.37 a | 2.07 b | 13.25 a | 13.25 a | 147.59 a |
| 2017 Ratoon crop | ||||||||||
| April 2018 | 20.53 a | 2.55 a | 9.94 a | 4.49 a | 1.22 a | 60.43 a | 1.79 a | 11.96 a | 16.45 b | 165.36 a |
| May 2018 | 20.13 a | 2.08 b | 8.33 b | 3.93 b | 1.20 a | 63.83 a | 1.48 ab | 9.99 b | 21.51 a | 155.13 a |
| June 2018 | 20.32 a | 2.30 b | 7.59 b | 3.82 b | 1.08 a | 59.04 a | 1.45 b | 10.67 ab | 19.43 a | 158.91 a |
| Effect | d.f | Stalks m−2 | Stalk Weight | TCH | KST | TSH |
|---|---|---|---|---|---|---|
| Nitrogen (N) | 3 | 0.0051 | 0.2259 | 0.0004 | 0.6237 | 0.0188 |
| Fungicides (F) | 3 | 0.0920 | 0.7040 | 0.0175 | 0.8881 | 0.1592 |
| N × F | 9 | 0.6689 | 0.2902 | 0.3668 | 0.9508 | 0.3730 |
| Year (Y) | 1 | 0.0837 | 0.0004 | <0.0001 | <0.0001 | 0.1338 |
| N × Y | 3 | 0.1783 | 0.8602 | 0.1738 | 0.4977 | 0.8351 |
| F × Y | 3 | 0.7233 | 0.0603 | 0.2570 | 0.0635 | 0.4632 |
| N × F × Y | 9 | 0.8990 | 0.9718 | 0.9172 | 0.6495 | 0.9363 |
| Crop (C) | 1 | 0.0031 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| N × C | 3 | 0.5961 | 0.8299 | 0.2608 | 0.7421 | 0.4319 |
| F × C | 3 | 0.0853 | 0.1167 | 0.1291 | 0.6843 | 0.0863 |
| N × F × C | 9 | 0.5674 | 0.2653 | 0.4970 | 0.5402 | 0.8165 |
| Y × C | 1 | <0.0001 | <0.0001 | <0.0001 | 0.0007 | <0.0001 |
| N × Y × C | 3 | 0.0047 | 0.2033 | 0.0003 | 0.7163 | 0.0258 |
| N × F × Y × C | 3 | 0.3343 | 0.4237 | 0.5952 | 0.3643 | 0.9329 |
| Main Effects | Plant Crop 2015 | Plant Crop 2016 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Stalks m−2 | Stalk Weight (kg) | TCH (t ha−1) | KST (kg t−1) | TSH (t ha−1) | Stalks m−2 | Stalk Weight (kg) | TCH (t ha−1) | KST (kg t−1) | TSH (t ha−1) | Dry Biomass (t ha−1) | |
| Nitrogen (N) | |||||||||||
| N0 | 10.97 a | 1.15 a | 126.17 a | 131.11 a | 16.53 a | 11.54 b | 1.14 b | 132.31 c | 109.01 a | 14.41 c | 37.23 c |
| N50 | 10.92 a | 1.16 a | 127.71 a | 131.86 a | 16.83 a | 11.88 ab | 1.19 ab | 141.94 b | 106.41 a | 15.11 bc | 40.13 b |
| N100 | 10.95 a | 1.14 a | 124.94 a | 127.51 a | 15.94 a | 12.04 ab | 1.20 a | 145.09 b | 106.45 a | 15.44 b | 40.32 b |
| N50+50 | 11.07 a | 1.14 a | 126.53 a | 130.94 a | 16.57 a | 12.38 a | 1.22 a | 150.94 a | 108.34 a | 16.28 a | 42.74 a |
| Fungicide (F) | |||||||||||
| Untreated | 10.63 c | 1.18 a | 125.65 ab | 128.29 a | 16.11 a | 11.70 a | 1.17 a | 137.06 b | 107.48 a | 14.73 b | 38.55 c |
| Mancozeb | 11.02 b | 1.09 b | 120.50 b | 132.24 a | 15.93 a | 11.92 a | 1.20 a | 143.04 b | 106.04 a | 15.15 ab | 40.03 b |
| Mefenoxam | 11.45 a | 1.15 ab | 131.88 a | 131.20 a | 17.34 a | 12.30 a | 1.19 a | 146.69 a | 107.51 a | 15.77 a | 41.52 a |
| Azoxystrobin | 10.80 bc | 1.17 a | 127.32 ab | 129.67 a | 16.50 a | 11.92 a | 1.20 a | 142.89 b | 109.19 a | 15.61 a | 40.42 ab |
| p-Values | |||||||||||
| N | 0.6247 | 0.8279 | 0.8937 | 0.6192 | 0.6395 | 0.0151 | 0.0346 | <0.0001 | 0.4924 | <0.001 | <0.0001 |
| F | <0.0001 | 0.0191 | 0.0240 | 0.6995 | 0.1991 | 0.1268 | 0.6549 | <0.0001 | 0.5184 | 0.0075 | <0.0001 |
| N × F | 0.0449 | 0.1785 | 0.0728 | 0.9456 | 0.5309 | 0.4596 | 0.1365 | 0.2377 | 0.1856 | 0.0525 | 0.4473 |
| Main Effects | Ratoon Crop 2016 | Ratoon Crop 2017 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Stalks m−2 | Stalk Weight (kg) | TCH (t ha−1) | KST (kg t−1) | TSH (t ha−1) | Dry Biomass (t ha−1) | Stalks m−2 | Stalk Weight (kg) | TCH (t ha−1) | KST (kg t−1) | TSH (t ha−1) | Dry Biomass (t ha−1) | |
| Nitrogen (N) | ||||||||||||
| N0 | 11.05 b | 0.844 a | 93.36 b | 128.47 a | 11.94 b | 29.66 b | 10.90 b | 1.33 a | 145.51 b | 94.84 a | 13.79 a | 43.50 b |
| N50 | 10.99 b | 0.863 a | 94.88 b | 126.57 a | 12.01 b | 30.02 b | 10.82 b | 1.36 a | 147.28 ab | 92.50 a | 13.63 a | 44.09 ab |
| N100 | 10.99 b | 0.890 a | 96.79 b | 131.09 a | 12.65 b | 31.01 ab | 11.37 a | 1.34 a | 152.67 a | 92.42 a | 14.10 a | 45.80 a |
| N50+50 | 12.15 a | 0.878 a | 106.37 a | 127.57 a | 13.66 a | 34.14 a | 10.92 b | 1.33 a | 145.37 b | 94.90 a | 13.78 a | 43.95 ab |
| Fungicide (F) | ||||||||||||
| Untreated | 11.12 a | 0.863 a | 95.58 a | 127.97 a | 12.33 a | 30.25 a | 11.01 a | 1.30 b | 144.18 a | 95.33 a | 13.73 a | 43.49 a |
| Mancozeb | 11.23 a | 0.875 a | 98.40 a | 125.76 a | 12.38 a | 31.26 a | 10.93 a | 1.37 a | 149.75 a | 94.19 a | 14.10 a | 44.98 a |
| Mefenoxam | 11.10 a | 0.880 a | 97.11 a | 130.69 a | 12.73 a | 31.25 a | 11.05 a | 1.33 ab | 148.93 a | 89.07 a | 13.27 a | 44.21 a |
| Azoxystrobin | 11.70 a | 0.856 a | 100.32 a | 129.62 a | 11.91 a | 32.07 a | 10.91 a | 1.35 ab | 147.98 a | 96.06 a | 14.20 a | 44.66 a |
| p-values | ||||||||||||
| N | 0.0739 | 0.6790 | 0.0990 | 0.6725 | 0.0872 | 0.0448 | 0.0002 | 0.6247 | 0.0275 | 0.7448 | 0.8228 | 0.0497 |
| F | 0.6216 | 0.9275 | 0.8540 | 0.5774 | 0.7916 | 0.7642 | 0.2407 | 0.0522 | 0.1741 | 0.1111 | 0.2665 | 0.3361 |
| N × F | 0.8382 | 0.8897 | 0.7969 | 0.9598 | 0.8239 | 0.7044 | 0.5071 | 0.3012 | 0.6188 | 0.5392 | 0.2858 | 0.6047 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vuyyuru, M.; Sandhu, H.S.; McCray, M.J.; Raid, R.N.; Erickson, J.E. Effects of Nitrogen Fertilization and Seed Piece Applied Fungicides on Establishment, Tiller Dynamics, and Sucrose Yields in Successively Planted Sugarcane. Agronomy 2019, 9, 387. https://doi.org/10.3390/agronomy9070387
Vuyyuru M, Sandhu HS, McCray MJ, Raid RN, Erickson JE. Effects of Nitrogen Fertilization and Seed Piece Applied Fungicides on Establishment, Tiller Dynamics, and Sucrose Yields in Successively Planted Sugarcane. Agronomy. 2019; 9(7):387. https://doi.org/10.3390/agronomy9070387
Chicago/Turabian StyleVuyyuru, Murali, Hardev S. Sandhu, Mabry J. McCray, Richard N. Raid, and John E. Erickson. 2019. "Effects of Nitrogen Fertilization and Seed Piece Applied Fungicides on Establishment, Tiller Dynamics, and Sucrose Yields in Successively Planted Sugarcane" Agronomy 9, no. 7: 387. https://doi.org/10.3390/agronomy9070387
APA StyleVuyyuru, M., Sandhu, H. S., McCray, M. J., Raid, R. N., & Erickson, J. E. (2019). Effects of Nitrogen Fertilization and Seed Piece Applied Fungicides on Establishment, Tiller Dynamics, and Sucrose Yields in Successively Planted Sugarcane. Agronomy, 9(7), 387. https://doi.org/10.3390/agronomy9070387