Exogenous ABA Induces Osmotic Adjustment, Improves Leaf Water Relations and Water Use Efficiency, But Not Yield in Soybean under Water Stress



Abstract
:1. Introduction
2. Material and Methods
2.1. ABA Treatment for All Experiments
2.2. Experiment 1
2.3. Experiment 2
2.4. Experiment 3
2.5. Statistical Analysis
3. Results
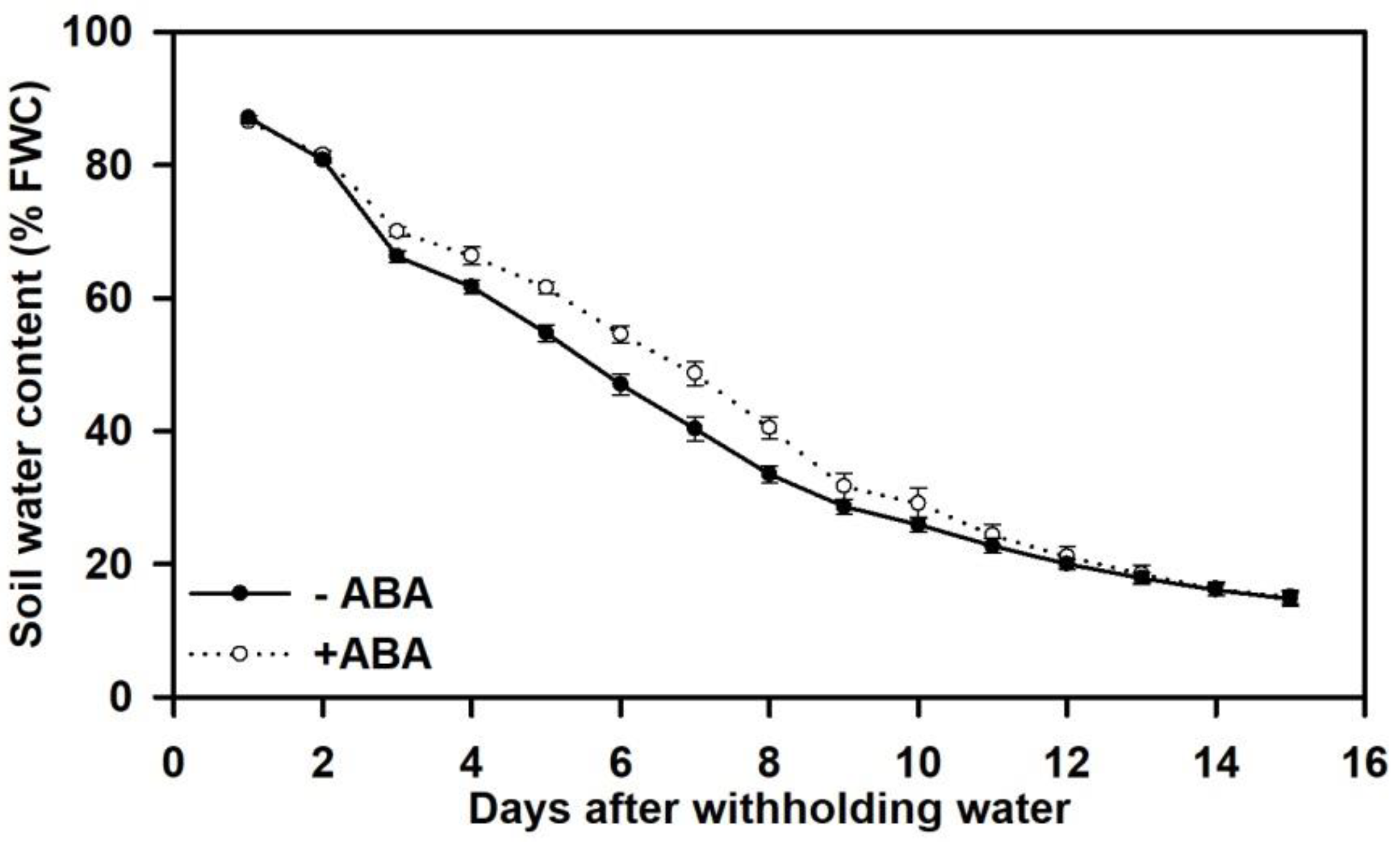
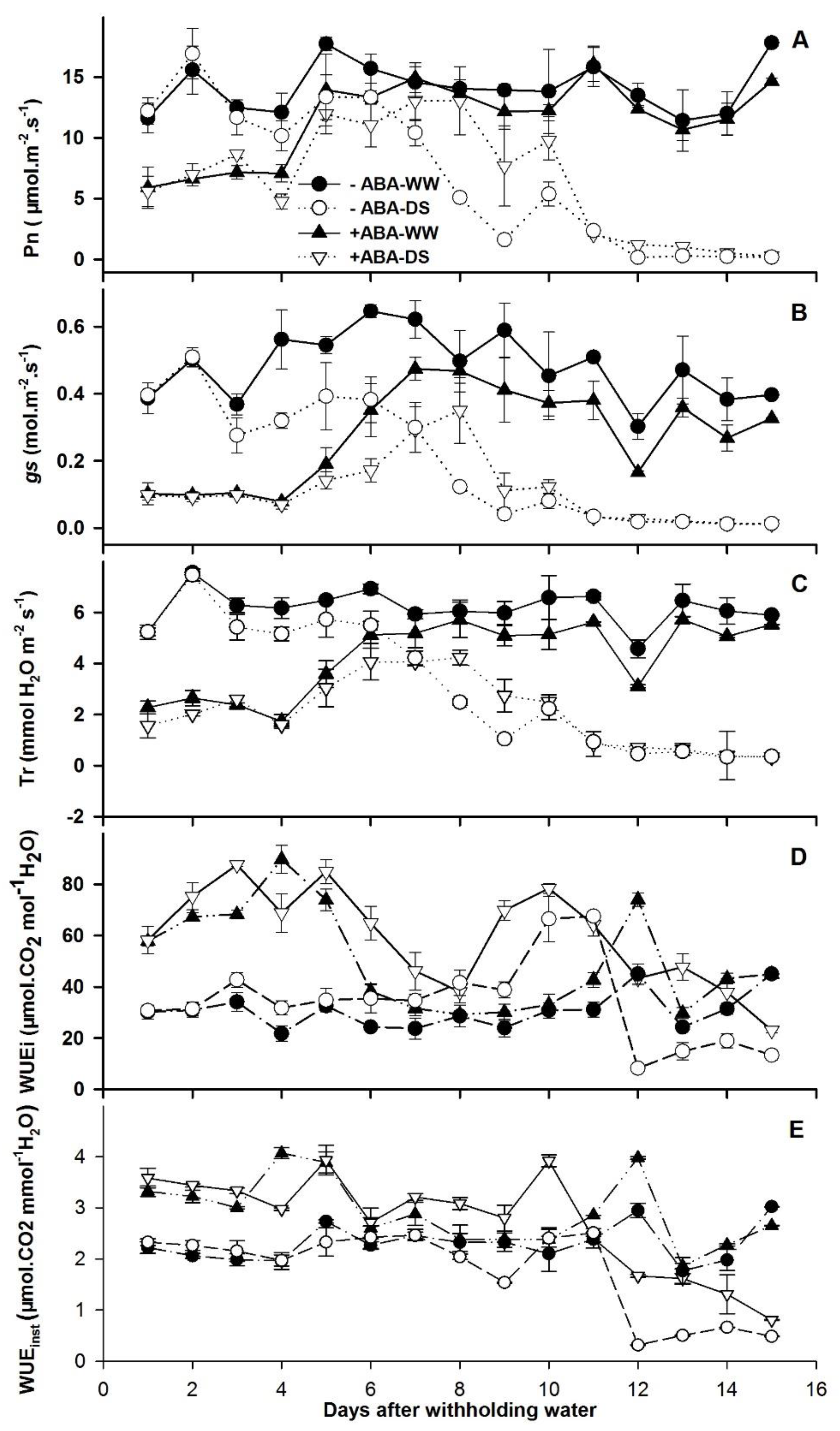
3.1. Soil Drying, Leaf ABA Concentration, Photosynthesis and WUE at the Leaf Level
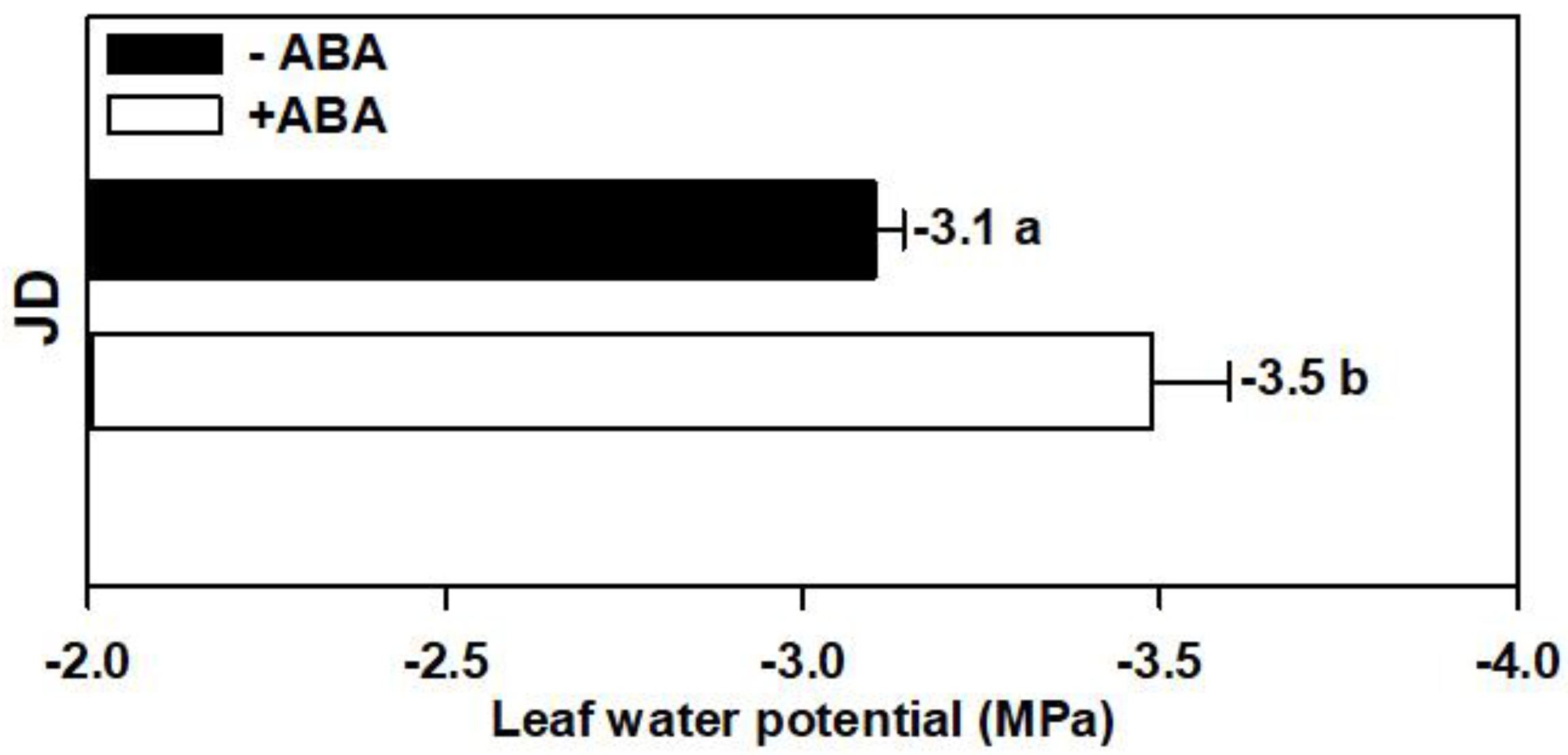
3.2. Leaf Water Relations, Desiccant Tolerance and Osmotic Adjustment
3.3. Flower and Pod Numbers, Yield, Yield Components, and WUEg
4. Discussion
4.1. Exogenous ABA Improved Leaf Desiccant Tolerance and RWC under Water Stress by Increasing OA
4.2. Exogenous ABA Had No Effect on Grain Yield But Increased WUEg under Water Stress
4.3. Exogenous Applications of ABA Increased WUEi and WUEinst by Reducing gs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Koornneef, M.; Hanhart, C.J.; Hilhorst, H.W.M.; Karssen, C.M. Invivo inhibition of seed development and reserve protein accumulation in recombinants of abscisic-acid biosynthesis and responsiveness mutants in Arabidopsis-Thaliana. Plant Physiol. 1989, 90, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, S.; Davies, W.J. Drought, ozone, ABA and ethylene: New insights from cell to plant to community. Plant Cell Environ. 2010, 33, 510–525. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Davies, W.J. Changes in the concentration of ABA in xylem sap as a function of changing soil-water status can account for changes in leaf conductance and growth. Plant Cell Environ. 1990, 13, 277–285. [Google Scholar] [CrossRef]
- Davies, W.J.; Zhang, J.H. Root signals and the regulation of growth and development of plants in drying soil. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 55–76. [Google Scholar] [CrossRef]
- Du, Y.L.; Wang, Z.Y.; Fan, J.W.; Turner, N.C.; Wang, T.; Li, F.M. β-Aminobutyric acid increases abscisic acid accumulation and desiccation tolerance and decreases water use but fails to improve grain yield in two spring wheat cultivars under soil drying. J. Exp. Bot. 2012, 63, 4849–4860. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.L.; Wang, Z.Y.; Fan, J.W.; Turner, N.C.; He, J.; Wang, T.; Li, F.M. Exogenous abscisic acid reduces water loss and improves antioxidant defence, desiccation tolerance and transpiration efficiency in two spring wheat cultivars subjected to a soil water deficit. Funct. Plant Biol. 2013, 40, 494–506. [Google Scholar] [CrossRef]
- Travaglia, C.; Reinoso, H.; Cohen, A.; Luna, C.; Tommasino, E.; Castillo, C.; Bottini, R. Exogenous ABA increases yield in field-grown wheat with moderate water restriction. J. Plant Growth Regul. 2010, 29, 366–374. [Google Scholar] [CrossRef]
- Hussain, S.; Saleem, M.F.; Ashraf, M.Y.; Cheema, M.A.; Haq, M.A. Abscisic acid, a stress hormone helps in improving water relations and yield of sunflower (Helianthus annuus L.) hybrids under drought. Pak. J. Bot. 2010, 42, 2177–2189. [Google Scholar]
- He, J.; Du, Y.L.; Wang, T.; Turner, N.C.; Xi, Y.; Li, F.M. Old and new cultivars of soya bean (Glycine max L.) subjected to soil drying differ in abscisic acid accumulation, water relations characteristics and yield. J. Agron. Crop Sci. 2016, 202, 372–383. [Google Scholar] [CrossRef]
- Gago, J.; Douthea, C.; Florez-Sarasab, I.; Escalonaa, J.M.; Galmesa, J.; Fernieb, A.R.; Flexasa, J.; Medrano, H. Opportunities for improving leaf water use efficiency under climatechange conditions. Plant Sci. 2014, 226, 108–119. [Google Scholar] [CrossRef]
- Jones, M.M.; Turner, N.C. Osmotic adjustment in expanding and fully expanded leaves of sunflower in response to water deficits. Aust. J. Plant Physiol. 1980, 7, 181–192. [Google Scholar] [CrossRef]
- Morgan, J.M. Osmoregulation and water-stress in higher-plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1984, 35, 299–319. [Google Scholar] [CrossRef]
- Turner, N.C.; Wright, G.C.; Siddique, K.H.H. Adaptation of grain legumes (pulses) to water-limited environments. Adv. Agron. 2001, 71, 193–231. [Google Scholar]
- Turner, N.C. Turgor maintenance by osmotic adjustment, an adaptive mechanism for coping with plant water deficits. Plant Cell Environ. 2016, 40, 1–3. [Google Scholar] [CrossRef] [PubMed]
- James, A.T.; Lawn, R.J.; Cooper, M. Genotypic variation for drought stress response traits in soybean. I. Variation in soybean and wild Glycine spp. for epidermal conductance, osmotic potential, and relative water content. Aust. J. Agric. Res. 2008, 59, 656–669. [Google Scholar] [CrossRef]
- James, A.T.; Lawn, R.J.; Cooper, M. Genotypic variation for drought stress response traits in soybean. II. Inter-relations between epidermal conductance, osmotic potential, relative water content, and plant survival. Aust. J. Agric. Res. 2008, 59, 670–678. [Google Scholar] [CrossRef]
- Blum, A. Osmotic adjustment is a prime drought stress adaptive engine in support of plant production. Plant Cell Environ. 2017, 40, 4–10. [Google Scholar] [CrossRef]
- Osakabe, Y.; Arinaga, N.; Umezawa, T.; Katsura, S.; Nagamachi, K.; Tanaka, H.; Ohiraki, H.; Yamada, K.; Seo, S.U.; Abo, M.; et al. Osmotic stress responses and plant growth controlled by potassium transporters in Arabidopsis. Plant Cell 2013, 25, 609–624. [Google Scholar] [CrossRef]
- Turner, N.C. Measurement of plant water status by the pressure chamber technique. IRRI Sci. 1988, 9, 289–308. [Google Scholar] [CrossRef]
- Fan, X.W.; Li, F.M.; Song, L.; Xiong, Y.C.; An, L.Z.; Jia, Y.; Fang, X.W. Defense strategy of old and modern spring wheat varieties during soil drying. Physiol. Plant. 2009, 136, 310–323. [Google Scholar] [CrossRef]
- Turner, N.C.; Abbo, S.; Berger, J.D.; Chaturvedi, S.K.; French, R.J.; Ludwig, C.; Mannur, D.M.; Singh, S.J.; Yadava, H.S. Osmotic adjustment in chickpea (Cicer arietinum L.) results in no yield benefit under terminal drought. J. Exp. Bot. 2007, 58, 187–194. [Google Scholar] [CrossRef] [PubMed]
- He, Z. Enzyme linked Immunosorbent Assay for Endogenous Plant Hormones. In Guidance to Experiment on Chemical Control in Crop Plants; He, Z., Ed.; Beijing Agricultural University Publishers: Beijing, China, 1993; pp. 60–68. [Google Scholar]
- Yang, J.C.; Zhang, J.H.; Wang, Z.Q.; Zhu, Q.S.; Wang, W. Hormonal changes in the grains of rice subjected to water stress during grain filling. Plant Physiol. 2001, 127, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Auge, R.M.; Kubikova, E.; Moore, J.L. Foliar dehydration tolerance of mycorrhizal cowpea, soybean and bush bean. New Phytol. 2001, 151, 535–541. [Google Scholar] [CrossRef]
- Turner, N.C.; Jones, M.M. Turgor maintenance by osmotic adjustment: A review and evaluation. In Adaptation of Plants to Water and High Temperature Stress; Turner, N.C., Kramer, P.J., Eds.; Wiley: New York, NY, USA, 1980; pp. 87–103. [Google Scholar]
- Sinclair, T.R. Water and nitrogen limitations in soybean grain production: I. Model development. Field Crops Res. 1986, 15, 125–141. [Google Scholar] [CrossRef]
- Egli, D.B. Flowering, pod Set and reproductive success in Soya Bean. J. Agron. Crop Sci. 2005, 191, 283–291. [Google Scholar] [CrossRef]
- Palta, J.A.; Ludwig, C. Pod set and seed yield as affected by cytokininapplication and terminal drought in narrow-leafed lupin. Aust. J. Agric. Res. 1997, 48, 81–90. [Google Scholar] [CrossRef]
- Dracup, M.; Reader, M.A.; Palta, J.A. Variation in yield of narrow-leafed lupin caused by terminal drought. Aust. J. Agric. Res. 1998, 49, 799–810. [Google Scholar] [CrossRef]
- Leport, L.; Turner, N.C.; Davies, S.L.; Siddique, K.H.M. Variation in podproduction and abortion among chickpea cultivars under terminal drought. Eur. J. Agron. 2006, 24, 236–246. [Google Scholar] [CrossRef]
- Fang, X.W.; Turner, N.C.; Yan, G.J.; Li, F.M.; Siddique, K.H.M. Flower numbers, pod production, pollen viability, and pistil function are reduced and flower and pod abortion increased in chickpea (Cicer arietinum L.) under terminal drought. J. Exp. Bot. 2010, 61, 335–345. [Google Scholar] [CrossRef]
- Farooq, M.; Gogoi, N.; Barthakur, S.; Baroowa, B.; Bharadwaj, N.; Alghamdi, S.S.; Siddique, K.H.M. Drought stress in grain legumes during reproduction and grain filling. J. Agron. Crop Sci. 2017, 203, 81–102. [Google Scholar] [CrossRef]
- Liu, F.; Jensen, C.R.; Andersen, M.N. Pod set related to photosynthetic rate and endogenous ABA in soybeans subjected to different water regimes and exogenous ABA and BA at early reproductive stages. Ann. Bot. 2004, 94, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Fischer, R.A.; Turner, N.C. Plant productivity in the arid and semiarid zones. Annu. Rev. Plant Physiol. 1978, 29, 277–317. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Treatment | Rate of Soil Drying (% h−1) | Soil Water Content (% FC) | Days After Withholding Water | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| H1 | H2 | H3 | H4 | H5 | H6 | H4 | H5 | H6 | ||
| −ABA | 0.22 a | 85.4 ± 0.3 a | 66.2 ± 0.9 a | 54.7 ± 1.2 a | 33.5 ± 1.2 a | 20.0 ± 0.8 a | 16.1 ± 0.9 a | 8 | 12 | 14 |
| +ABA | 0.21 b | 84.6 ± 0.7 a | 66.2 ± 0.5 a | 54.6 ± 1.3 a | 29.1 ± 2.3 b | 18.5 ± 1.3 a | 15.1 ± 1.1 a | 10 | 13 | 15 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, J.; Jin, Y.; Palta, J.A.; Liu, H.-Y.; Chen, Z.; Li, F.-M. Exogenous ABA Induces Osmotic Adjustment, Improves Leaf Water Relations and Water Use Efficiency, But Not Yield in Soybean under Water Stress. Agronomy 2019, 9, 395. https://doi.org/10.3390/agronomy9070395
He J, Jin Y, Palta JA, Liu H-Y, Chen Z, Li F-M. Exogenous ABA Induces Osmotic Adjustment, Improves Leaf Water Relations and Water Use Efficiency, But Not Yield in Soybean under Water Stress. Agronomy. 2019; 9(7):395. https://doi.org/10.3390/agronomy9070395
Chicago/Turabian StyleHe, Jin, Yi Jin, Jairo A. Palta, Hong-Yan Liu, Zhu Chen, and Feng-Min Li. 2019. "Exogenous ABA Induces Osmotic Adjustment, Improves Leaf Water Relations and Water Use Efficiency, But Not Yield in Soybean under Water Stress" Agronomy 9, no. 7: 395. https://doi.org/10.3390/agronomy9070395
APA StyleHe, J., Jin, Y., Palta, J. A., Liu, H. -Y., Chen, Z., & Li, F. -M. (2019). Exogenous ABA Induces Osmotic Adjustment, Improves Leaf Water Relations and Water Use Efficiency, But Not Yield in Soybean under Water Stress. Agronomy, 9(7), 395. https://doi.org/10.3390/agronomy9070395