Cytosolic 5′-Nucleotidase II Is a Sensor of Energy Charge and Oxidative Stress: A Possible Function as Metabolic Regulator

 , ,
, ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Purification of Recombinant NT5C2
2.3. NT5C2 Activity Assays
2.4. Measurement of Kinetic Parameters of Recombinant NT5C2
2.5. Measurement of Recombinant NT5C2 Activity at Different Adenylate Energy Charge Values
2.6. Inactivation of NT5C2 in ADF Cells by H2O2
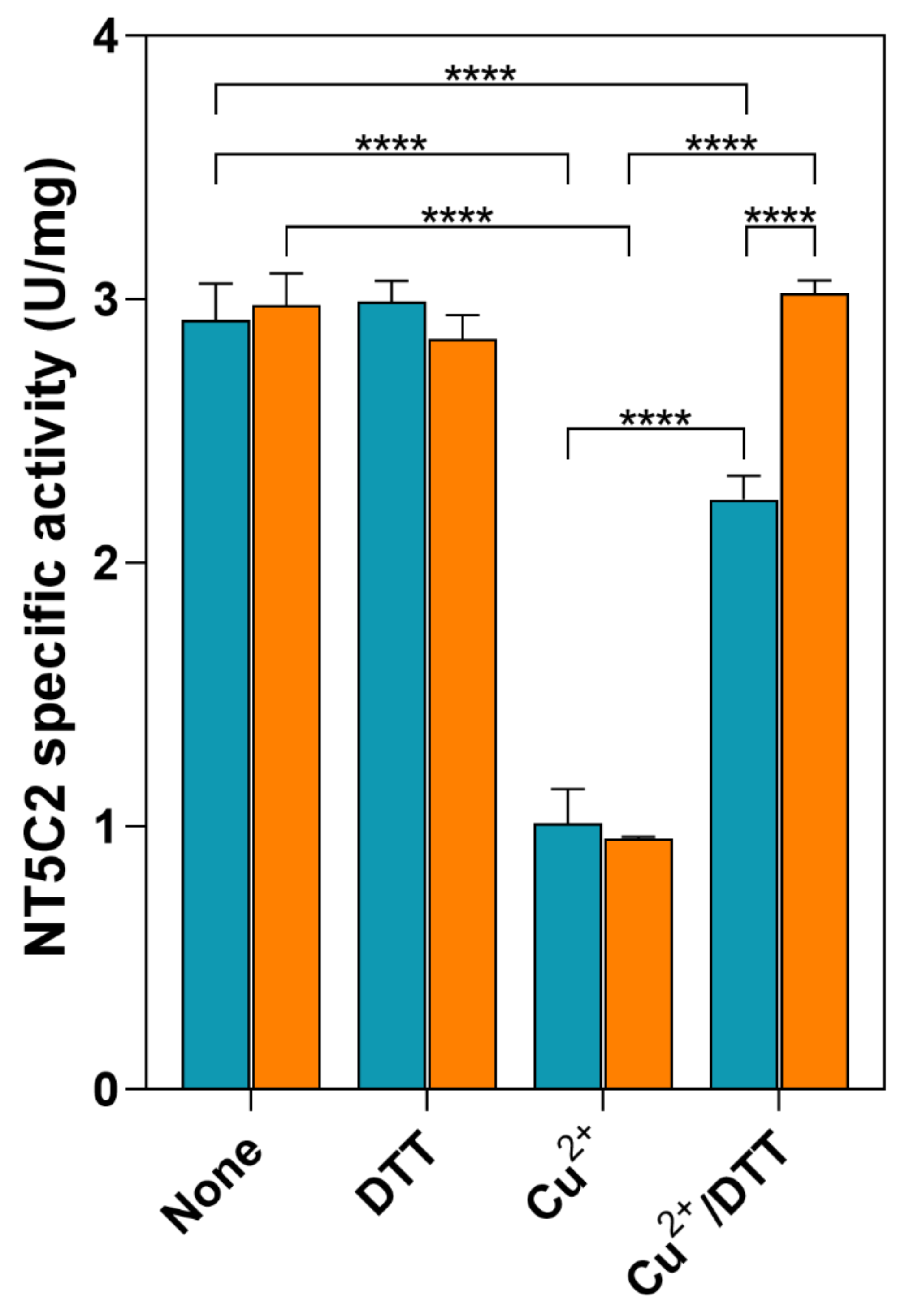
2.7. Inactivation of Recombinant NT5C2 by CuCl2
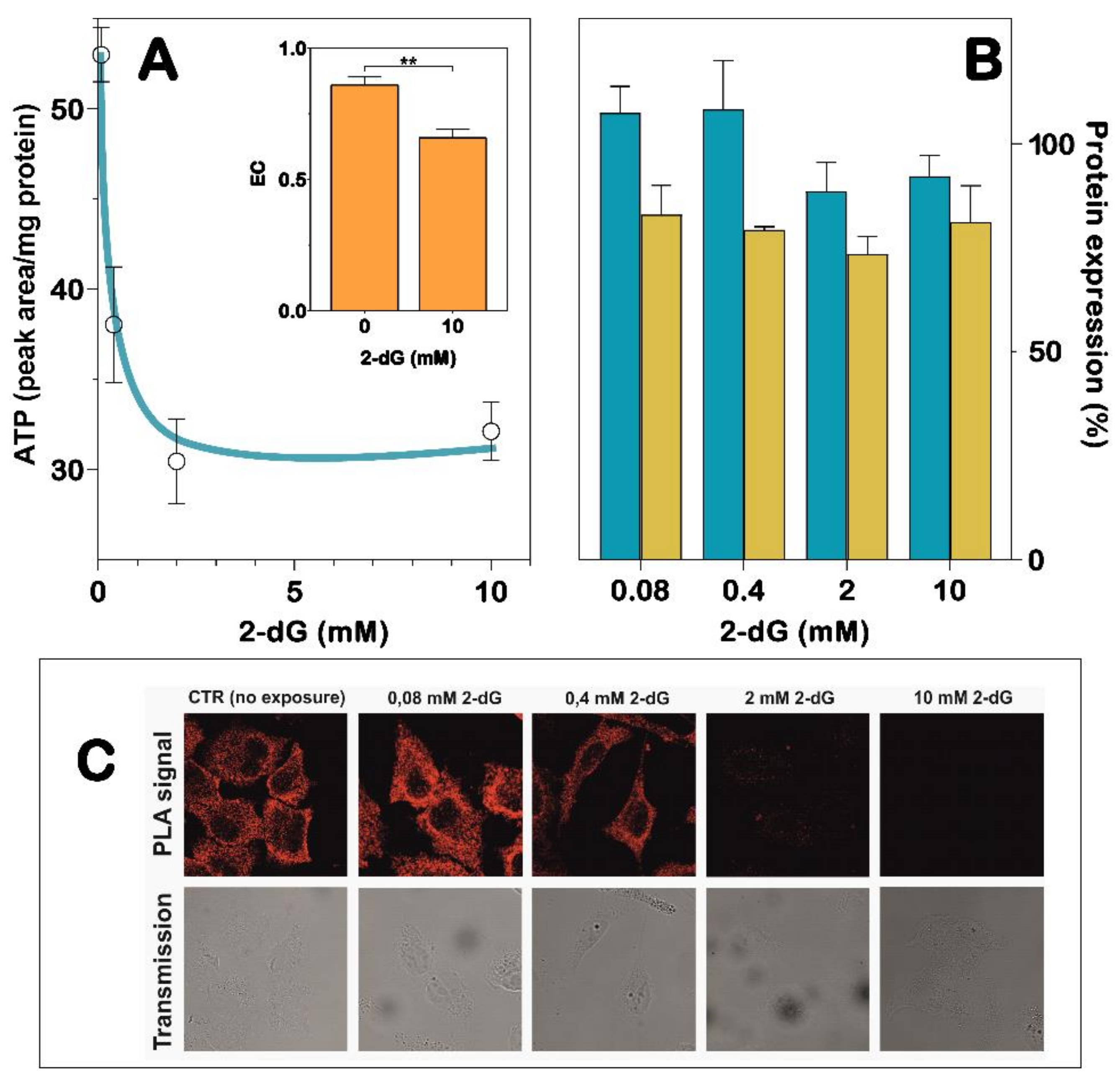
2.8. NT5C2/IPAF Proximity Ligation Assay in 2-dG-Exposed A549 Cells
2.9. Preparation of A549 Cell Lysates for ELISA Test and HPLC Analysis
2.10. ELISA Test
2.11. Other Methods
3. Results
3.1. The NT5C2 Activity Is Dependent on Adenylate Energy Charge
3.2. Effect of Oxidative Stress on NT5C2 in ADF Cells
3.3. Interaction of NT5C2 with IPAF in A549 Cells Is Affected by Oxidative Stress and Energy Charge
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tozzi, M.G.; Pesi, R.; Allegrini, S. On the Physiological Role of Cytosolic 5’-nucleotidase II (cN-II): Pathological and Therapeutical Implications. Curr. Med. Chem. 2013, 20, 4285–4291. [Google Scholar] [CrossRef]
- Baiocchi, C.; Pesi, R.; Camici, M.; Itoh, R.; Tozzi, M.G. Mechanism of the reaction catalysed by cytosolic 5′-nucleotidase/phosphotransferase: Formation of a phosphorylated intermediate. Biochem. J. 1996, 317, 797–801. [Google Scholar] [CrossRef] [Green Version]
- Itoh, R. Enzymatic Properties and Physiological Roles of Cytosolic 5’-Nucleotidase II. Curr. Med. Chem. 2013, 20, 4260–4284. [Google Scholar] [CrossRef] [PubMed]
- Walldén, K.; Stenmark, P.; Nyman, T.; Flodin, S.; Gräslund, S.; Loppnau, P.; Bianchi, V.; Nordlund, P. Crystal structure of human cytosolic 5’-nucleotidase II: Insights into allosteric regulation and substrate recognition. J. Biol. Chem. 2007, 282, 17828–17836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walldén, K.; Nordlund, P. Structural Basis for the Allosteric Regulation and Substrate Recognition of Human Cytosolic 5′-Nucleotidase II. J. Mol. Biol. 2011, 408, 684–696. [Google Scholar] [CrossRef] [PubMed]
- Dieck, C.L.; Tzoneva, G.; Forouhar, F.; Carpenter, Z.; Ambesi-Impiombato, A.; Sánchez-Martín, M.; Kirschner-Schwabe, R.; Lew, S.; Seetharaman, J.; Tong, L.; et al. Structure and Mechanisms of NT5C2 Mutations Driving Thiopurine Resistance in Relapsed Lymphoblastic Leukemia. Cancer Cell 2018, 34, 136.e6–147.e6. [Google Scholar] [CrossRef] [Green Version]
- Allegrini, S.; Careddu, M.G.; Cuccu, G.; Pesi, R.; Camici, M.; Ferrara, L.; Scaloni, A.; D’Ambrosio, C.; Tozzi, M.G. Mechanistic studies on bovine cytosolic 5’-nucleotidase II, an enzyme belonging to the HAD superfamily. JBIC J. Biol. Inorg. Chem. 2004, 271, 4881–4891. [Google Scholar] [CrossRef]
- Pesi, R.; Petrotto, E.; Colombaioni, L.; Allegrini, S.; Garcia-Gil, M.; Camici, M.; Jordheim, L.P.; Tozzi, M.G. Cytosolic 5′-Nucleotidase II Silencing in a Human Lung Carcinoma Cell Line Opposes Cancer Phenotype with a Concomitant Increase in p53 Phosphorylation. Int. J. Mol. Sci. 2018, 19, 2115. [Google Scholar] [CrossRef] [Green Version]
- Cividini, F.; Filoni, D.N.; Pesi, R.; Allegrini, S.; Camici, M.; Tozzi, M.G. IMP–GMP specific cytosolic 5′-nucleotidase regulates nucleotide pool and prodrug metabolism. Biochim. Biophys. Acta BBA Gen. Subj. 2015, 1850, 1354–1361. [Google Scholar] [CrossRef]
- Rampazzo, C.; Gazziola, C.; Ferraro, P.; Gallinaro, L.; Johansson, M.; Reichard, P.A.; Bianchi, V. Human high-Km 5’-nucleotidase effects of overexpression of the cloned cDNA in cultured human cells. JBIC J. Biol. Inorg. Chem. 1999, 261, 689–697. [Google Scholar] [CrossRef]
- Allegrini, S.; Filoni, D.N.; Galli, A.; Collavoli, A.; Pesi, R.; Camici, M.; Tozzi, M.G. Expression of Bovine Cytosolic 5′-Nucleotidase (cN-II) in Yeast: Nucleotide Pools Disturbance and Its Consequences on Growth and Homologous Recombination. PLoS ONE 2013, 8, e63914. [Google Scholar] [CrossRef] [PubMed]
- Bricard, G.; Cros-Perrial, E.; Machon, C.; Dumontet, C.; Jordheim, L.P. Stably transfected adherent cancer cell models with decreased expression of 5′-nucleotidase cN-II. Nucleosides Nucleotides Nucleic Acids 2016, 35, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Cadassou, O.; Raza, M.Z.; Machon, C.; Gudefin, L.; Armanet, C.; Chettab, K.; Guitton, J.; Tozzi, M.G.; Dumontet, C.; Cros-Perrial, E.; et al. Enhanced migration of breast and lung cancer cells deficient for cN-II and CD73 via COX-2/PGE2/AKT axis regulation. Cell Oncol. 2020, 1–15. [Google Scholar] [CrossRef]
- Kviklyte, S.; Vertommen, D.; Yerna, X.; Andersén, H.; Xu, X.; Gailly, P.; Bohlooly, Y.M.; Oscarsson, J.; Rider, M.H. Effects of genetic deletion of soluble 5′-nucleotidases NT5C1A and NT5C2 on AMPK activation and nucleotide levels in contracting mouse skeletal muscles. Am. J. Physiol. Metab. 2017, 313, E48–E62. [Google Scholar] [CrossRef] [PubMed]
- Johanns, M.; Kviklyte, S.; Chuang, S.-J.; Corbeels, K.; Jacobs, R.; Herinckx, G.; Vertommen, D.; Schakman, O.; Duparc, T.; Cani, P.D.; et al. Genetic deletion of soluble 5′-nucleotidase II reduces body weight gain and insulin resistance induced by a high-fat diet. Mol. Genet. Metab. 2019, 126, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Cividini, F.; Cros-Perrial, E.; Pesi, R.; Machon, C.; Allegrini, S.; Camici, M.; Dumontet, C.; Jordheim, L.; Tozzi, M. Cell proliferation and drug sensitivity of human glioblastoma cells are altered by the stable modulation of cytosolic 5′-nucleotidase II. Int. J. Biochem. Cell Biol. 2015, 65, 222–229. [Google Scholar] [CrossRef]
- Tzoneva, G.; Dieck, C.L.; Oshima, K.; Ambesi-Impiombato, A.; Sánchez-Martín, M.; Madubata, C.J.; Khiabanian, H.; Yu, J.; Waanders, E.; Iacobucci, I.; et al. Clonal evolution mechanisms in NT5C2 mutant-relapsed acute lymphoblastic leukaemia. Nat. Cell Biol. 2018, 553, 511–514. [Google Scholar] [CrossRef]
- Moriyama, T.; Liu, S.-G.; Li, J.; Meyer, J.; Zhao, X.; Yang, W.; Shao, Y.; Heath, R.; Hnizda, A.; Carroll, W.L.; et al. Mechanisms of NT5C2-Mediated Thiopurine Resistance in Acute Lymphoblastic Leukemia. Mol. Cancer Ther. 2019, 18, 1887–1895. [Google Scholar] [CrossRef] [Green Version]
- Camici, M.; Allegrini, S.; Tozzi, M.G. Interplay between adenylate metabolizing enzymes and AMP-activated protein kinase. FEBS J. 2018, 285, 3337–3352. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, S.S.; Karlsson, H.K.R.; Szekeres, F.; Chibalin, A.V.; Krook, A.; Zierath, J.R. Suppression of 5′-Nucleotidase Enzymes Promotes AMP-activated Protein Kinase (AMPK) Phosphorylation and Metabolism in Human and Mouse Skeletal Muscle. J. Biol. Chem. 2011, 286, 34567–34574. [Google Scholar] [CrossRef] [Green Version]
- Duarte, R.R.; Bachtel, N.D.; Côtel, M.-C.; Lee, S.H.; Selvackadunco, S.; Watson, I.A.; Hovsepian, G.A.; Troakes, C.; Breen, G.D.; Nixon, D.F.; et al. The Psychiatric Risk Gene NT5C2 Regulates Adenosine Monophosphate-Activated Protein Kinase Signaling and Protein Translation in Human Neural Progenitor Cells. Biol. Psychiatry 2019, 86, 120–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordheim, L.P. Expanding the clinical relevance of the 5′-nucleotidase cN-II/NT5C2. Purinergic Signal. 2018, 14, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Camici, M.; Garcia-Gil, M.; Tozzi, M.G. The Inside Story of Adenosine. Int. J. Mol. Sci. 2018, 19, 784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cividini, F.; Tozzi, M.G.; Galli, A.; Pesi, R.; Camici, M.; Dumontet, C.; Jordheim, L.P.; Allegrini, S. Cytosolic 5’-Nucleotidase II Interacts with the Leucin Rich Repeat of NLR Family Member Ipaf. PLoS ONE 2015, 10, e0121525. [Google Scholar] [CrossRef] [Green Version]
- Allegrini, S.; Pesi, R.; Tozzi, M.G.; Fiol, J.C.; Johnson, B.R.; Eriksson, S. Bovine cytosolic IMP/GMP-specific 5′-nucleotidase: Cloning and expression of active enzyme in Escherichia coli. Biochem. J. 1997, 328, 483–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tozzi, M.G.; Camici, M.; Pesi, R.; Allegrini, S.; Sgarrella, F.; Ipata, P. Nucleoside phosphotransferase activity of human colon carcinoma cytosolic 5′-nucleotidase. Arch. Biochem. Biophys. 1991, 291, 212–217. [Google Scholar] [CrossRef]
- Pesi, R.; Turriani, M.; Allegrini, S.; Scolozzi, C.; Camici, M.; Ipata, P.; Tozzi, M.G. The Bifunctional Cytosolic 5′-Nucleotidase: Regulation of the Phosphotransferase and Nucleotidase Activities. Arch. Biochem. Biophys. 1994, 312, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Itoh, R.; Oka, J.; Ozasa, H.; Headrick, J.P.; Willis, R.J.; Fokina, O.; Herrmann, C.; Forchhammer, K.; Verhoeven, A.J.M.; Marszalek, J.; et al. Regulation of rat heart cytosol 5′-nucleotidase by adenylate energy charge. Biochem. J. 1986, 235, 847–851. [Google Scholar] [CrossRef] [Green Version]
- Balestri, F.; Giannecchini, M.; Sgarrella, F.; Carta, M.C.; Tozzi, M.G.; Camici, M. Purine and pyrimidine nucleosides preserve human astrocytoma cell adenylate energy charge under ischemic conditions. Neurochem. Int. 2007, 50, 517–523. [Google Scholar] [CrossRef]
- Micheli, V.; Sestini, S.; Rocchigiani, M.; Jacomelli, G.; Manzoni, F.; Peruzzi, L.; Gathof, B.; Zammarchi, E.; Pompucci, G. Hypoxanthine-guanine phosphoribosyltransferase deficiency and erythrocyte synthesis of pyridine coenzymes. Life Sci. 1999, 64, 2479–2487. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bevington, A.; Kemp, G.J.; Russell, R.G.G. Factors Controlling the Intracellular Concentration of Orthophosphate (Pi) in Mammalian Cells. In Phosphate and Mineral Homeostasis. Advances in Experimental Medicine and Biology; Massry, S.G., Olmer, M., Ritz, E., Eds.; Springer: Boston, MA, USA, 1986; Volume 208, pp. 469–478. [Google Scholar] [CrossRef]
- Greenwood, H.E.; McCormick, P.N.; Gendron, T.; Glaser, M.; Pereira, R.; Maddocks, O.D.K.; Sander, K.; Zhang, T.; Koglin, N.; Lythgoe, M.F.; et al. Measurement of Tumor Antioxidant Capacity and Prediction of Chemotherapy Resistance in Preclinical Models of Ovarian Cancer by Positron Emission Tomography. Clin. Cancer Res. 2019, 25, 2471–2482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peroni, E.; Scali, V.; Balestri, F.; Cappiello, M.; Mura, U.; Del-Corso, A.; Moschini, R. Pathways of 4-Hydroxy-2-Nonenal Detoxification in a Human Astrocytoma Cell Line. Antioxidants 2020, 9, 385. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Li, J.; Wang, F.; Hu, J.; Wang, S.; Sun, Y. 2-Deoxy-D-glucose targeting of glucose metabolism in cancer cells as a potential therapy. Cancer Lett. 2014, 355, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Tzoneva, G.; Perez-Garcia, A.; Carpenter, Z.; Khiabanian, H.; Tosello, V.; Allegretta, M.; Paietta, E.; Racevskis, J.; Rowe, J.M.; Tallman, M.S.; et al. Activating mutations in the NT5C2 nucleotidase gene drive chemotherapy resistance in relapsed ALL. Nat. Med. 2013, 19, 368–371. [Google Scholar] [CrossRef] [Green Version]
- Careddu, M.G.; Allegrini, S.; Pesi, R.; Camici, M.; Garcia-Gil, M.; Tozzi, M.G. Knockdown of cytosolic 5′-nucleotidase II (cN-II) reveals that its activity is essential for survival in astrocytoma cells. Biochim. Biophys. Acta BBA Bioenerg. 2008, 1783, 1529–1535. [Google Scholar] [CrossRef] [Green Version]
- Jordheim, L.P.; Puy, J.-Y.; Cros-Perrial, E.; Peyrottes, S.; Lefebvre, I.; Périgaud, C.; Dumontet, C. Determination of the enzymatic activity of cytosolic 5′-nucleotidase cN-II in cancer cells: Development of a simple analytical method and related cell line models. Anal. Bioanal. Chem. 2015, 407, 5747–5758. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Peskin, A.V.; Parsons-Mair, H.N. Thiol Oxidase Activity of Copper, Zinc Superoxide Dismutase. J. Biol. Chem. 2002, 277, 1906–1911. [Google Scholar] [CrossRef] [Green Version]
- Balestri, F.; Moschini, R.; Cappiello, M.; Mura, U.; Del-Corso, A. Thiol oxidase ability of copper ion is specifically retained upon chelation by aldose reductase. JBIC J. Biol. Inorg. Chem. 2017, 22, 559–565. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Kinetic Parameters | WT | C547A |
|---|---|---|
| KM Ino (mM) | 1.00 ± 0.059 | 0.83 ± 0.061 |
| KM IMP (mM) | 0.10 ± 0.018 | 0.12 ± 0.022 |
| K50 Mg2+ (mM) | 2.00 ± 0.114 | 0.60 ± 0.101 |
| K50 ATP (mM) | 2.00 ± 0.190 | 1.00 ± 0.087 |
| Ki Pi (mM) | 2.00 ± 0.156 | 2.00 ± 0.218 |
| kcat (sec−1) | 52.90 ± 3.35 | 228.80 ± 16.56 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pesi, R.; Allegrini, S.; Balestri, F.; Garcia-Gil, M.; Cividini, F.; Colombaioni, L.; Jordheim, L.P.; Camici, M.; Tozzi, M.G. Cytosolic 5′-Nucleotidase II Is a Sensor of Energy Charge and Oxidative Stress: A Possible Function as Metabolic Regulator. Cells 2021, 10, 182. https://doi.org/10.3390/cells10010182
Pesi R, Allegrini S, Balestri F, Garcia-Gil M, Cividini F, Colombaioni L, Jordheim LP, Camici M, Tozzi MG. Cytosolic 5′-Nucleotidase II Is a Sensor of Energy Charge and Oxidative Stress: A Possible Function as Metabolic Regulator. Cells. 2021; 10(1):182. https://doi.org/10.3390/cells10010182
Chicago/Turabian StylePesi, Rossana, Simone Allegrini, Francesco Balestri, Mercedes Garcia-Gil, Federico Cividini, Laura Colombaioni, Lars Petter Jordheim, Marcella Camici, and Maria Grazia Tozzi. 2021. "Cytosolic 5′-Nucleotidase II Is a Sensor of Energy Charge and Oxidative Stress: A Possible Function as Metabolic Regulator" Cells 10, no. 1: 182. https://doi.org/10.3390/cells10010182
APA StylePesi, R., Allegrini, S., Balestri, F., Garcia-Gil, M., Cividini, F., Colombaioni, L., Jordheim, L. P., Camici, M., & Tozzi, M. G. (2021). Cytosolic 5′-Nucleotidase II Is a Sensor of Energy Charge and Oxidative Stress: A Possible Function as Metabolic Regulator. Cells, 10(1), 182. https://doi.org/10.3390/cells10010182