
First Episode Psychosis and Schizophrenia Are Systemic Neuro-Immune Disorders Triggered by a Biotic Stimulus in Individuals with Reduced Immune Regulation and Neuroprotection

 , , and
, , and
Abstract
:1. Introduction
2. Methods
2.1. Selection of Seed Proteins
2.2. PPI Network Construction
2.3. Enrichment Analysis
2.4. Annotation Analysis and Annotation Visualization
3. Results
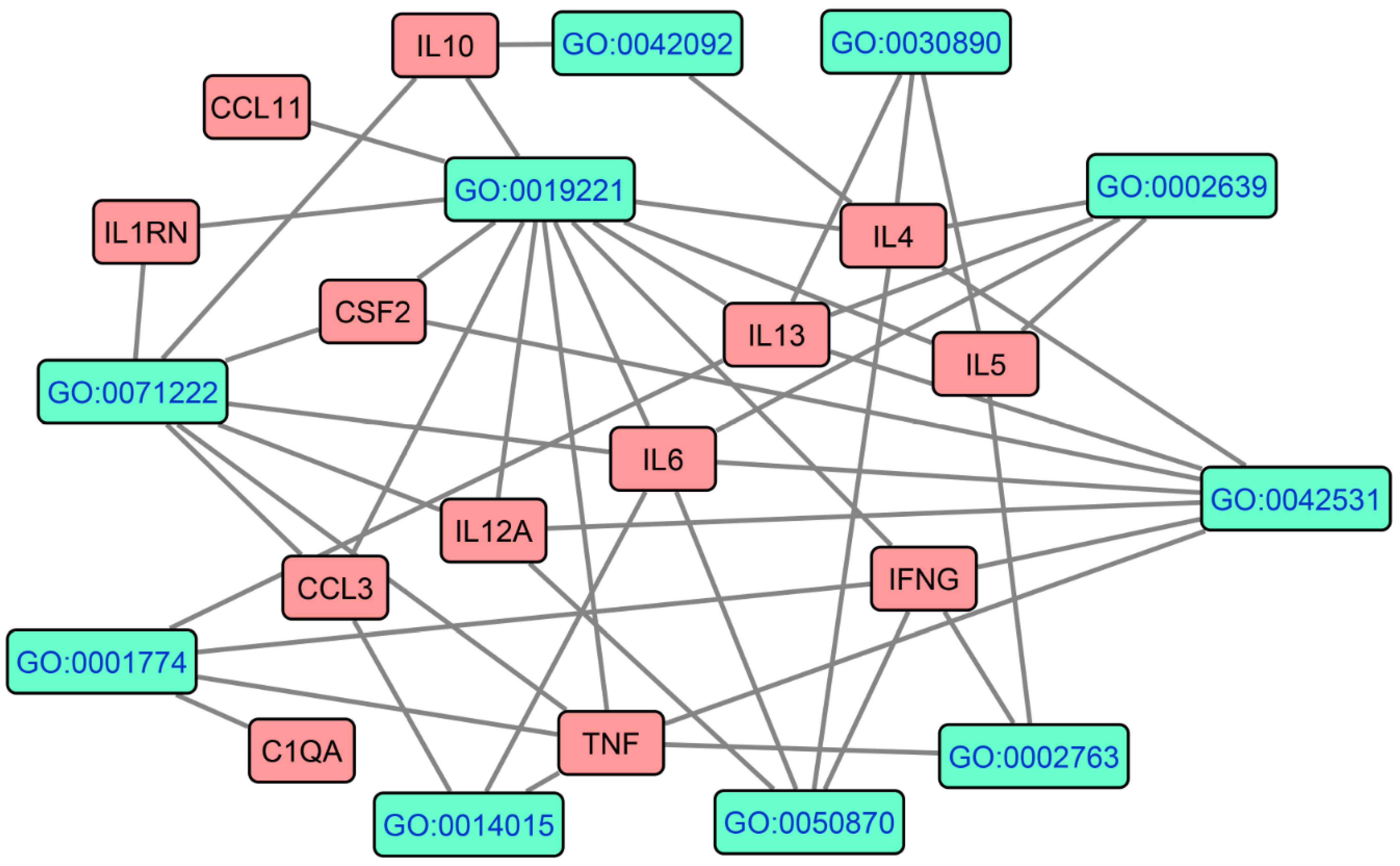
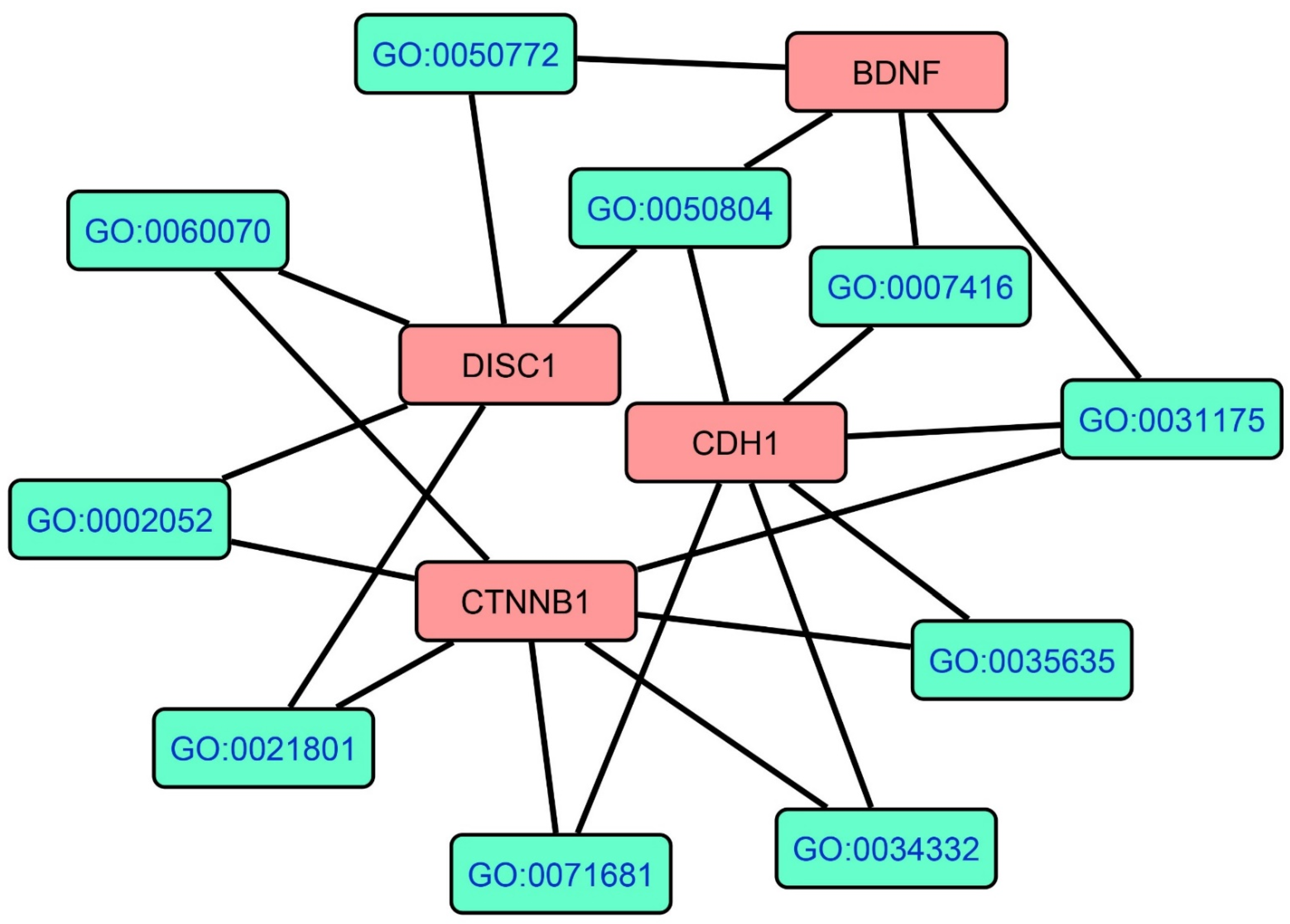
3.1. The PPI Network Topography of FEP and FES/FEP
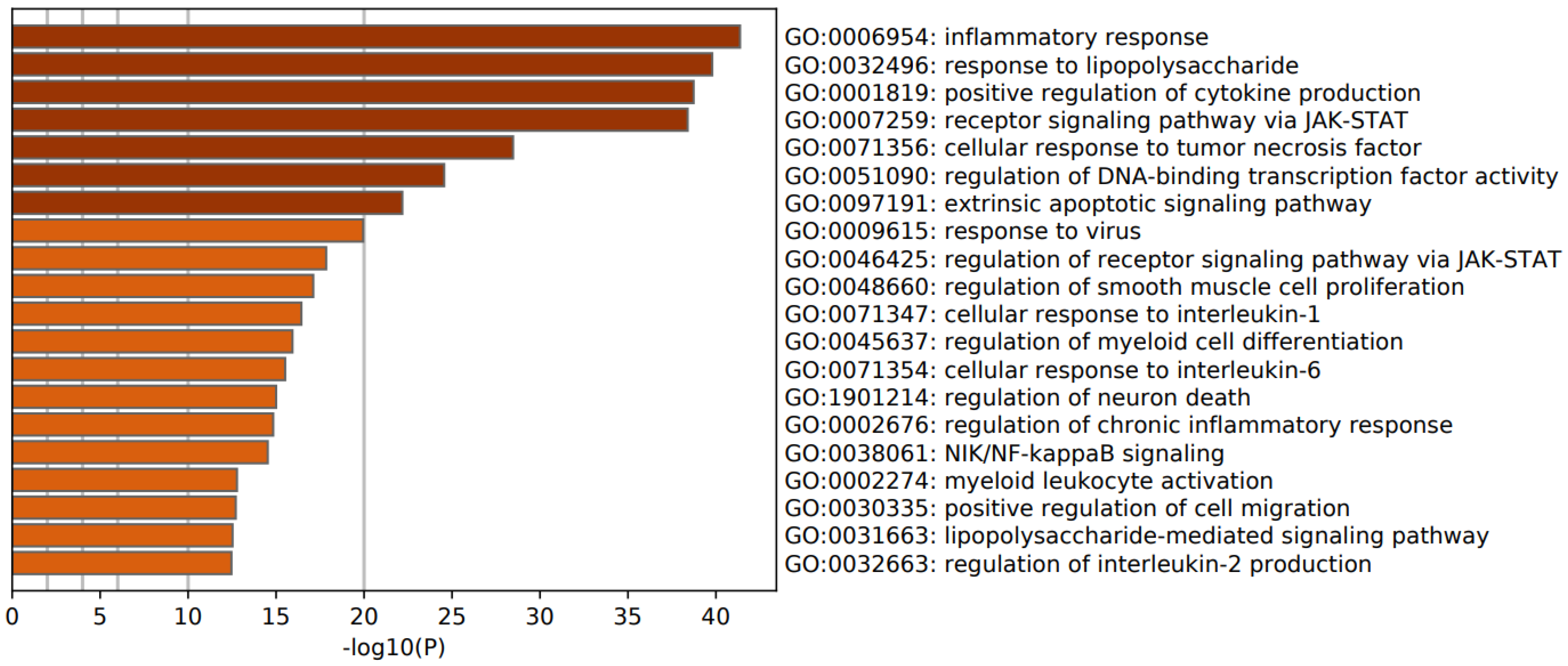
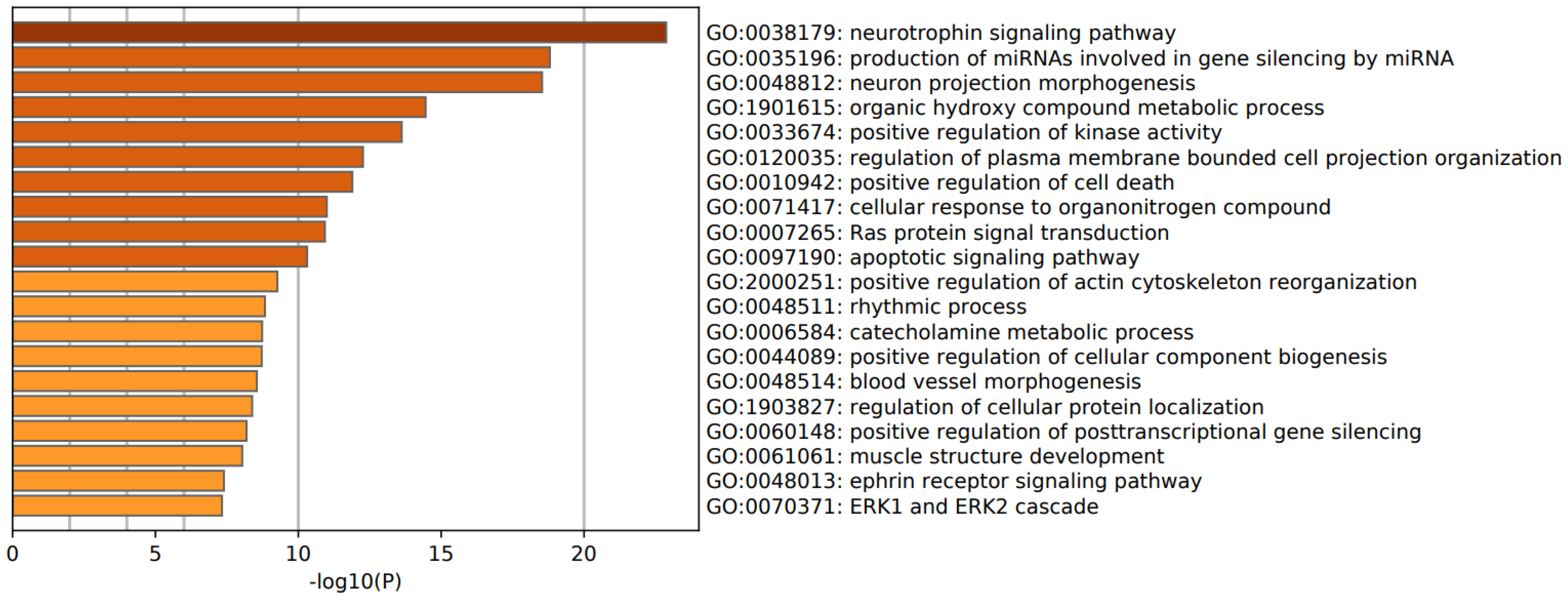
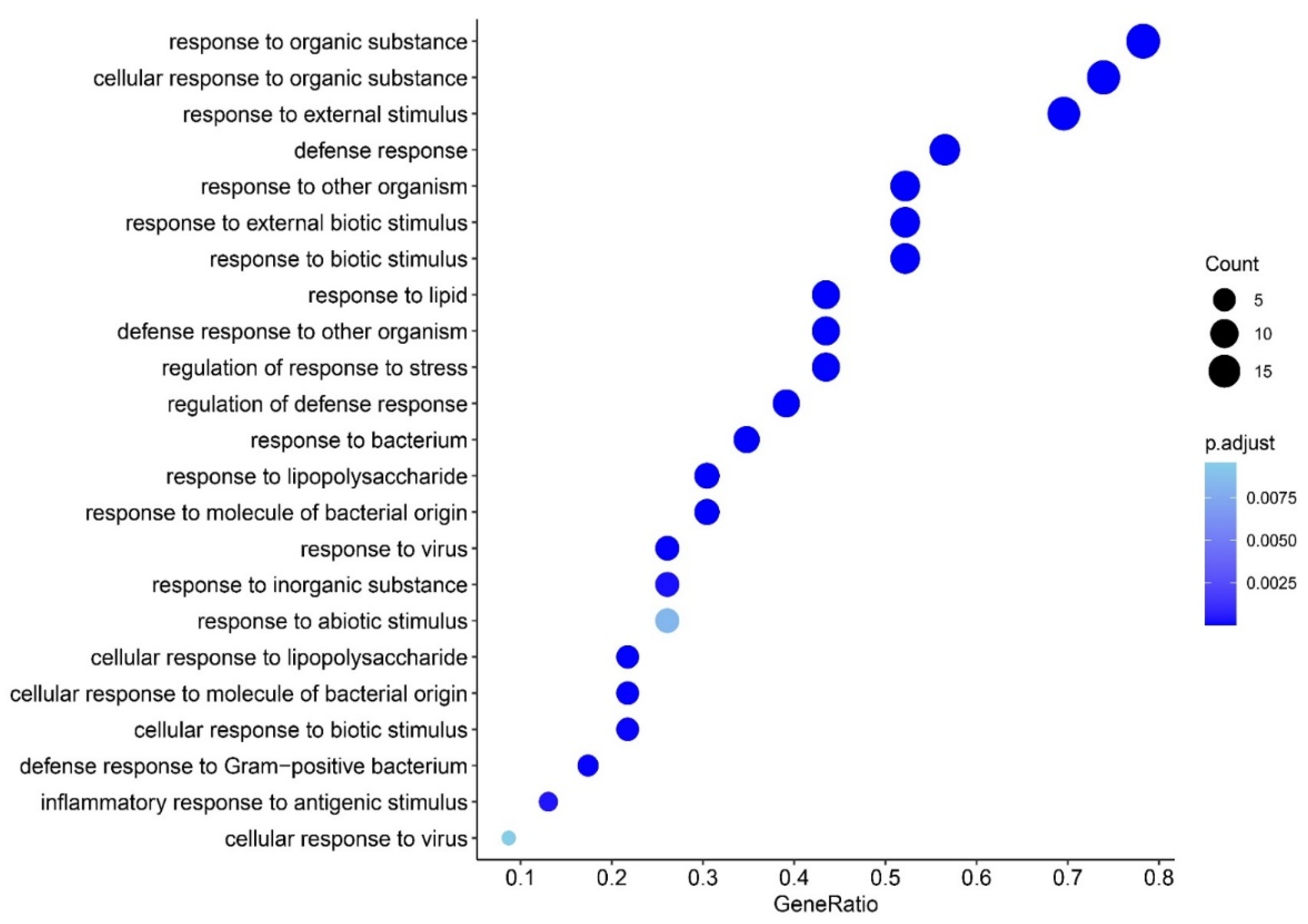
3.2. Enrichment Analysis in FEP
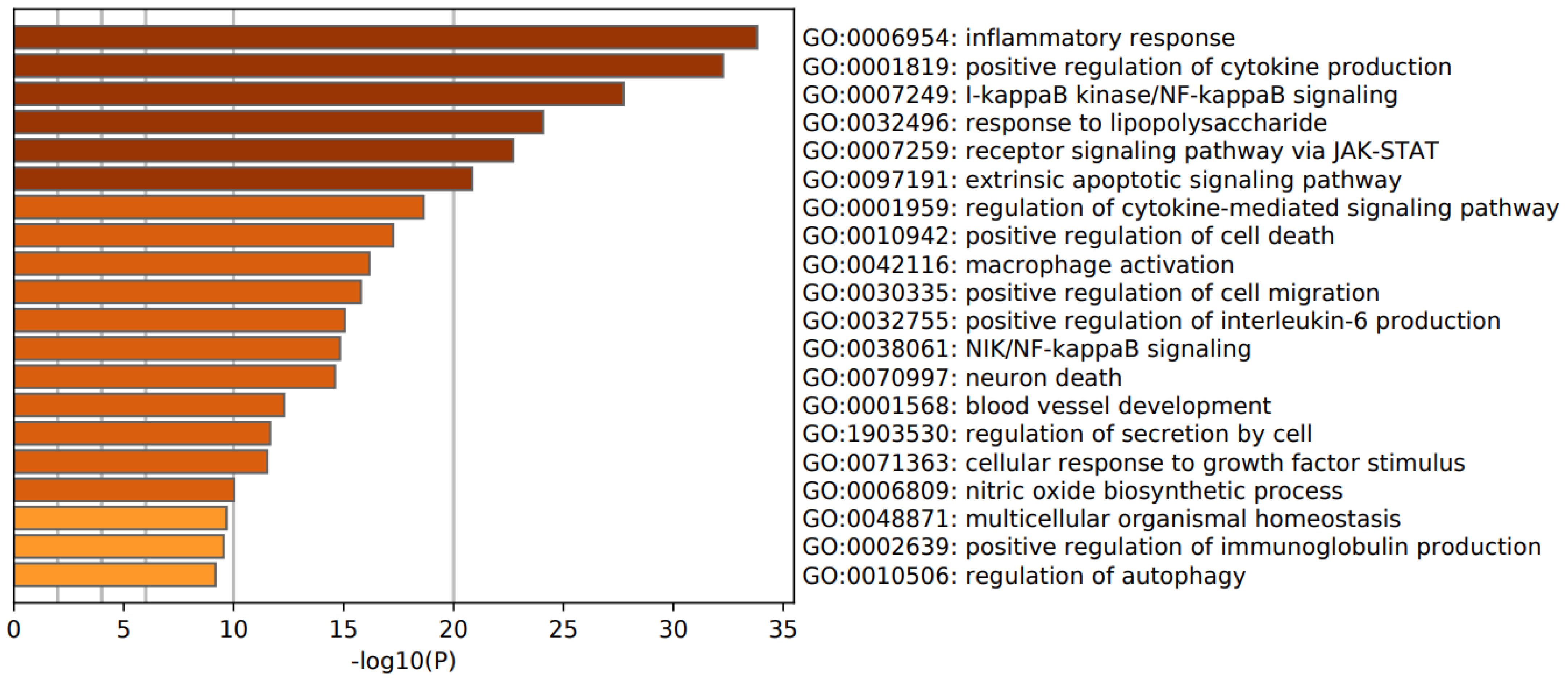
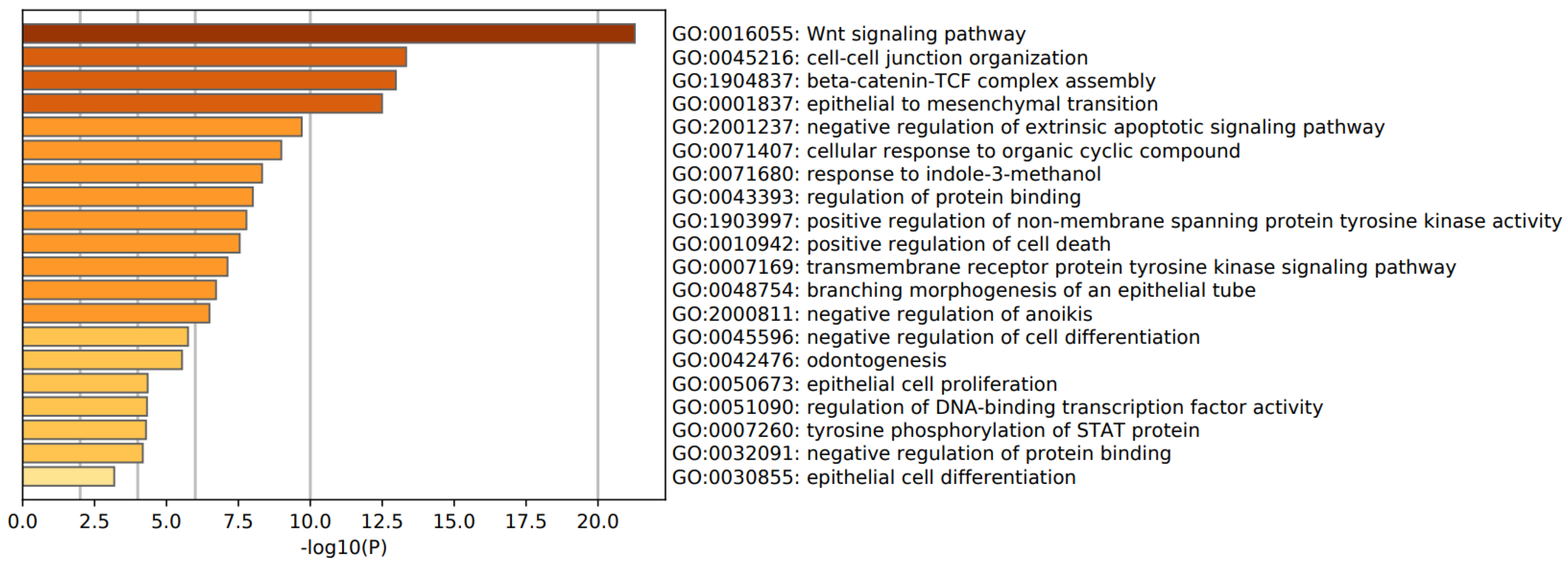
3.3. Enrichment Analysis in FEP/FES
3.4. Annotation Analysis and Visualization
4. Discussion
4.1. The PPI Network of FEP/FES
4.2. Trigger Factors in FEP and FEP/FES
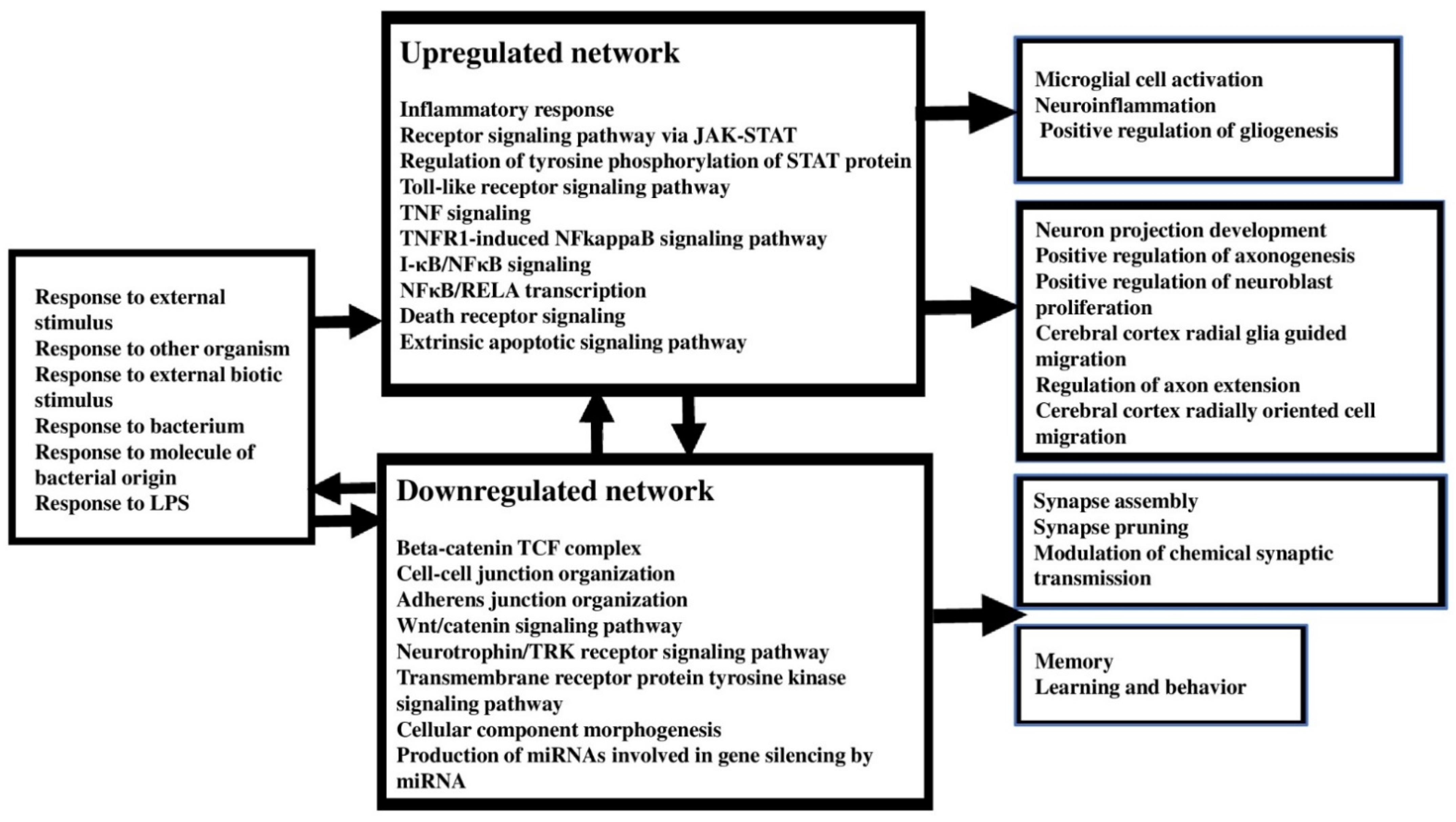
4.3. Upregulated Pathways, Molecular and Cellular Processes in FEP
4.4. Downregulated Pathways, Molecular and Cellular Processes in FEP
4.5. Pathways, Molecular and Cellular Processes in FEP/FES
4.6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smith, R.; Maes, M. The macrophage-T-lymphocyte theory of schizophrenia: Additional evidence. Med. Hypotheses 1995, 45, 135–141. [Google Scholar] [CrossRef]
- Maes, M.; Delange, J.; Ranjan, R.; Meltzer, H.Y.; Desnyder, R.; Cooremans, W.; Scharpé, S. Acute phase proteins in schizophrenia, mania and major depression: Modulation by psychotropic drugs. Psychiatry Res. 1997, 66, 1–11. [Google Scholar] [CrossRef]
- Maes, M.; Meltzer, H.Y.; Bosmans, E. Immune-inflammatory markers in schizophrenia: Comparison to normal controls and effects of clozapine. Acta Psychiatr. Scand. 1994, 89, 346–351. [Google Scholar] [CrossRef]
- Roomruangwong, C.; Noto, C.; Kanchanatawan, B.; Anderson, G.; Kubera, M.; Carvalho, A.F.; Maes, M. The Role of Aberrations in the Immune-Inflammatory Response System (IRS) and the Compensatory Immune-Regulatory Reflex System (CIRS) in Different Phenotypes of Schizophrenia: The IRS-CIRS Theory of Schizophrenia. Mol. Neurobiol. 2020, 57, 778–797. [Google Scholar] [CrossRef]
- Comer, A.L.; Carrier, M.; Tremblay, M.-È.; Cruz-Martín, A. The Inflamed Brain in Schizophrenia: The Convergence of Genetic and Environmental Risk Factors That Lead to Uncontrolled Neuroinflammation. Front. Cell. Neurosci. 2020, 14. [Google Scholar] [CrossRef] [PubMed]
- Boin, F.; Zanardini, R.; Pioli, R.; Altamura, C.A.; Maes, M.; Gennarelli, M. Association between −G308A tumor necrosis factor alpha gene polymorphism and schizophrenia. Mol. Psychiatry 2001, 6, 79–82. [Google Scholar] [CrossRef] [Green Version]
- Maes, M.; Vojdani, A.; Sirivichayakul, S.; Barbosa, D.S.; Kanchanatawan, B. Inflammatory and Oxidative Pathways Are New Drug Targets in Multiple Episode Schizophrenia and Leaky Gut, Klebsiella pneumoniae, and C1q Immune Complexes Are Additional Drug Targets in First Episode Schizophrenia. Mol. Neurobiol. 2021, 58, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Noto, M.N.; Maes, M.; Nunes, S.O.V.; Ota, V.K.; Rossaneis, A.C.; Verri, W.A., Jr.; Cordeiro, Q.; Belangero, S.I.; Gadelha, A.; Bressan, R.A.; et al. Activation of the immune-inflammatory response system and the compensatory immune-regulatory system in antipsychotic naive first episode psychosis. Eur. Neuropsychopharmacol. 2019, 29, 416–431. [Google Scholar] [CrossRef] [PubMed]
- Noto, C.; Ota, V.K.; Santoro, M.L.; Gouvea, E.S.; Silva, P.N.; Spindola, L.; Cordeiro, Q.; Bressan, R.; Gadelha, A.; Brietzke, E.; et al. Depression, Cytokine, and Cytokine by Treatment Interactions Modulate Gene Expression in Antipsychotic Naïve First Episode Psychosis. Mol. Neurobiol. 2016, 53, 5701–5709. [Google Scholar] [CrossRef] [PubMed]
- Noto, M.N.; Maes, M.; Nunes, S.O.V.; Ota, V.K.; Cavalcante, D.; Oliveira, G.; Rossaneis, A.C.; Verri, W.A.; Cordeiro, Q.; Belangero, S.I.; et al. BDNF in antipsychotic naive first episode psychosis: Effects of risperidone and the immune-inflammatory response system. J. Psychiatr. Res. 2021, 141, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Sirivichayakul, S.; Matsumoto, A.K.; Michelin, A.P.; Semeão, L.D.O.; Pedrão, J.V.D.L.; Moreira, E.G.; Barbosa, D.S.; Carvalho, A.F.; Solmi, M.; et al. Lowered Antioxidant Defenses and Increased Oxidative Toxicity Are Hallmarks of Deficit Schizophrenia: A Nomothetic Network Psychiatry Approach. Mol. Neurobiol. 2020, 57, 4578–4597. [Google Scholar] [CrossRef] [PubMed]
- Jindal, R.D.; Pillai, A.K.; Mahadik, S.P.; Eklund, K.; Montrose, D.M.; Keshavan, M.S. Decreased BDNF in patients with antipsychotic naïve first episode schizophrenia. Schizophr. Res. 2010, 119, 47–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maes, M.; Kanchanatawan, B. In (deficit) schizophrenia, a general cognitive decline partly mediates the effects of neuro-immune and neuro-oxidative toxicity on the symptomatome and quality of life. CNS Spectrums 2021, 12, 1–10. [Google Scholar] [CrossRef]
- Chen, S.J.; Liao, D.L.; Chen, C.H.; Wang, T.Y.; Chen, K.C. Construction and Analysis of Protein-Protein Interaction Network of Heroin Use Disorder. Sci Rep 2019, 9, 4980. [Google Scholar] [CrossRef]
- Brinholi, F.F.; Noto, C.; Maes, M.; Bonifácio, K.L.; Brietzke, E.; Ota, V.K.; Gadelha, A.; Cordeiro, Q.; Belangero, S.; Bressan, R.; et al. Lowered paraoxonase 1 (PON1) activity is associated with increased cytokine levels in drug naïve first episode psychosis. Schizophr. Res. 2015, 166, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Noto, C.; Ota, V.K.; Gadelha, A.; Noto, M.N.; Barbosa, D.S.; Bonifácio, K.L.; Nunes, S.O.; Cordeiro, Q.; Belangero, S.; Bressan, R.; et al. Oxidative stress in drug naïve first episode psychosis and antioxidant effects of risperidone. J. Psychiatr. Res. 2015, 68, 210–216. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Kanchanatawan, B.; Sirivichayakul, S.; Carvalho, A.F. In Schizophrenia, Increased Plasma IgM/IgA Responses to Gut Commensal Bacteria Are Associated with Negative Symptoms, Neurocognitive Impairments, and the Deficit Phenotype. Neurotox. Res. 2018, 35, 684–698. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.; Kershaw, N.J.; Babon, J.J. The molecular details of cytokine signaling via the JAK/STAT pathway. Protein Sci. 2018, 27, 1984–2009. [Google Scholar] [CrossRef] [Green Version]
- Murray, P.J. The JAK-STAT Signaling Pathway: Input and Output Integration. J. Immunol. 2007, 178, 2623–2629. [Google Scholar] [CrossRef] [Green Version]
- Gong, M.; Zhuo, X.; Ma, A. STAT6 Upregulation Promotes M2 Macrophage Polarization to Suppress Atherosclerosis. Med. Sci. Monit. Basic Res. 2017, 23, 240–249. [Google Scholar] [CrossRef]
- Sharma, R.P.; Rosen, C.; Melbourne, J.K.; Feiner, B.; Chase, K.A. Activated Phosphorylated STAT1 Levels as a Biologically Relevant Immune Signal in Schizophrenia. Neuroimmunomodulation 2016, 23, 224–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- UniProtB. UniProtKB—Q04206 (TF65_HUMAN). RELA—Transcription Factor p65—Homo Sapiens (Human)—RELA Gene & Protein. Available online: Uniprot.org (accessed on 23 July 2021).
- Volk, D.W.; Moroco, A.E.; Roman, K.M.; Edelson, J.R.; Lewis, D.A. The Role of the Nuclear Factor-κB Transcriptional Complex in Cortical Immune Activation in Schizophrenia. Biol. Psychiatry 2019, 85, 25–34. [Google Scholar] [CrossRef]
- Hashimoto, R.; Ohi, K.; Yasuda, Y.; Fukumoto, M.; Yamamori, H.; Takahashi, H.; Iwase, M.; Okochi, T.; Kazui, H.; Saitoh, O.; et al. Variants of the RELA Gene are Associated with Schizophrenia and their Startle Responses. Neuropsychopharmacology 2011, 36, 1921–1931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wajant, H.; Scheurich, P. TNFR1-induced activation of the classical NF-κB pathway. FEBS J. 2011, 278, 862–876. [Google Scholar] [CrossRef]
- Maes, M.; Sirivichayakul, S.; Matsumoto, A.K.; Maes, A.; Michelin, A.P.; de Oliveira Semeão, L.; de Lima Pedrão, J.V.; Moreira, E.G.; Barbosa, D.S.; Geffard, M.; et al. Increased Levels of Plasma Tumor Necrosis Factor-α Mediate Schizophrenia Symptom Dimensions and Neurocognitive Impairments and Are Inversely Associated with Natural IgM Directed to Malondialdehyde and Paraoxonase 1 Activity. Mol. Neurobiol. 2020, 57, 2333–2345. [Google Scholar] [CrossRef] [PubMed]
- Leopold, A.; Chernov, K.G.; Shemetov, A.A.; Verkhusha, V.V. Neurotrophin receptor tyrosine kinases regulated with near-infrared light. Nat. Commun. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correll, P.H.; Morrison, A.C.; Lutz, M.A. Receptor tyrosine kinases and the regulation of macrophage activation. J. Leukoc. Biol. 2004, 75, 731–737. [Google Scholar] [CrossRef]
- Creative Diagnostics. Neurotrophin Signaling Pathway. Neurotrophin Signaling Pathway—Creative Diagnostics. Available online: Crea-tive-diagnostics.com (accessed on 18 July 2021).
- Xu, D.; Lian, D.; Wu, J.; Liu, Y.; Zhu, M.; Sun, J.; He, D.; Li, L. Brain-derived neurotrophic factor reduces inflammation and hippocampal apoptosis in experimental Streptococcus pneumoniae meningitis. J. Neuroinflammation 2017, 14, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.-C.; Yao, W.; Hashimoto, K. Brain-derived Neurotrophic Factor (BDNF)-TrkB Signaling in Inflammation-related Depression and Potential Therapeutic Targets. Curr. Neuropharmacol. 2016, 14, 721–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, M.J.; Matheson, S.L.; Shepherd, A.; Weickert, C.S.; Carr, V.J. Brain-derived neurotrophic factor levels in schizophrenia: A systematic review with meta-analysis. Mol. Psychiatry 2011, 16, 960–972. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Su, Y.; Zhang, C.; Xing, M.; Ding, W.; Liao, L.; Guan, Y.; Li, Z.; Cui, N. The Interaction of BDNF and NTRK2 Gene Increases the Susceptibility of Paranoid Schizophrenia. PLoS ONE 2013, 8, e74264. [Google Scholar] [CrossRef] [Green Version]
- Hennah, W.; Thomson, P.; Peltonen, L.; Porteous, D. Genes and Schizophrenia: Beyond Schizophrenia: The Role of DISC1 in Major Mental Illness. Schizophr. Bull. 2006, 32, 409–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- UniProt. UniProtKB—Q9NRI5 (DISC1_HUMAN). DISC1—Disrupted in Schizophrenia 1 Protein—Homo Sapiens (Human)—DISC1 Gene & Protein. Available online: Uniprot.org (accessed on 18 July 2021).
- Chen, C.; Liu, H.-Y.; Hsueh, Y. TLR 3 downregulates expression of schizophrenia gene Disc1 via MYD 88 to control neuronal morphology. EMBO Rep. 2017, 18, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Aharoni, R.; Tobi, D. Dynamical comparison between Drosha and Dicer reveals functional motion similarities and dissimilarities. PLoS ONE 2019, 14, e0226147. [Google Scholar] [CrossRef]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in Control of Gene Expression: An Overview of Nuclear Functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tahamtan, A.; Teymoori-Rad, M.; Nakstad, B.; Salimi, V. Anti-Inflammatory MicroRNAs and Their Potential for Inflammatory Diseases Treatment. Front. Immunol. 2018, 9, 1377. [Google Scholar] [CrossRef] [Green Version]
- Caputo, V.; Ciolfi, A.; Macri, S.; Pizzuti, A. The emerging role of MicroRNA in schizophrenia. CNS Neurol. Disord.-Drug Targets 2015, 14, 208–221. [Google Scholar] [CrossRef]
- Cao, T.; Zhen, X.-C. Dysregulation of miRNA and its potential therapeutic application in schizophrenia. CNS Neurosci. Ther. 2018, 24, 586–597. [Google Scholar] [CrossRef]
- Topol, A.; Zhu, S.; Hartley, B.J.; English, J.; Hauberg, M.E.; Tran, N.; Rittenhouse, C.A.; Simone, A.; Ruderfer, D.; Johnson, J.; et al. Dysregulation of miRNA-9 in a Subset of Schizophrenia Patient-Derived Neural Progenitor Cells. Cell Rep. 2016, 15, 1024–1036. [Google Scholar] [CrossRef] [Green Version]
- Miller, B.H.; Zeier, Z.; Xi, L.; Lanz, T.; Deng, S.; Strathmann, J.; Willoughby, D.; Kenny, P.J.; Elsworth, J.D.; Lawrence, M.S.; et al. MicroRNA-132 dysregulation in schizophrenia has implications for both neurodevelopment and adult brain function. Proc. Natl. Acad. Sci. USA 2012, 109, 3125–3130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weigelt, K.; Bergink, V.; Burgerhout, K.M.; Pescatori, M.; Wijkhuijs, A.; Drexhage, H.A. Down-regulation of inflammation-protective microRNAs 146a and 212 in monocytes of patients with postpartum psychosis. Brain, Behav. Immun. 2013, 29, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.L.; Kao, P.F.; Itriago, E.; Zhan, Y.; Kozubek, J.A.; Hoss, A.G.; Banigan, M.G.; Vanderburg, C.R.; Rezvani, A.H.; Latourelle, J.C.; et al. miR-149 and miR-29c as candidates for bipolar disorder biomarkers. Am. J. Med Genet. Part B Neuropsychiatr. Genet. 2017, 174, 315–323. [Google Scholar] [CrossRef]
- van Rees, G.F. The Involvement of Microglial Activation in Schizophrenia; Queens College, University of Cambridge: Cambridge, UK, 2017. [Google Scholar]
- Rusznák, Z.; Henskens, W.; Schofield, E.; Kim, W.S.; Fu, Y. Adult Neurogenesis and Gliogenesis: Possible Mechanisms for Neurorestoration. Exp. Neurobiol. 2016, 25, 103–112. [Google Scholar] [CrossRef] [Green Version]
- García-Bueno, B.; Gassó, P.; MacDowell, K.S.; Callado, L.F.; Mas, S.; Bernardo, M.; Lafuente, A.; Meana, J.J.; Leza, J.C. Evidence of activation of the Toll-like receptor-4 proinflammatory pathway in patients with schizophrenia. J. Psychiatry Neurosci. 2016, 41, E46–E55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, X.; Liu, Z.; Niu, B.; Zhang, J.; Tan, T.K.; Lee, S.R.; Zhao, Y.; Harris, D.C.H.; Zheng, G. E-Cadherin/β-Catenin Complex and the Epithelial Barrier. J. Biomed. Biotechnol. 2011, 2011, 567305. [Google Scholar] [CrossRef] [Green Version]
- van Roy, F.; Berx, G. The cell-cell adhesion molecule E-cadherin. Cell Mol. Life Sci. 2008, 65, 3756–3788. [Google Scholar] [CrossRef] [PubMed]
- Oliva, C.A.; Vargas, J.Y.; Inestrosa, N.C. Wnts in adult brain: From synaptic plasticity to cognitive deficiencies. Front. Cell. Neurosci. 2013, 7, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jridi, I.; Canté-Barrett, K.; Pike-Overzet, K.; Staal, F.J.T. Inflammation and Wnt Signaling: Target for Immunomodulatory Therapy? Front. Cell Dev. Biol. 2021, 8, 615131. [Google Scholar] [CrossRef]
- Li, X.; Xiang, Y.; Li, F.; Yin, C.; Li, B.; Ke, X. WNT/β-Catenin Signaling Pathway Regulating T Cell-Inflammation in the Tumor Microenvironment. Front. Immunol. 2019, 10, 2293. [Google Scholar] [CrossRef] [Green Version]
- Silva-García, O.; Valdez-Alarcón, J.J.; Baizabal-Aguirre, V.M. The Wnt/β-Catenin Signaling Pathway Controls the Inflammatory Response in Infections Caused by Pathogenic Bacteria. Mediat. Inflamm. 2014, 2014, 310183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devaux, C.A.; Mezouar, S.; Mege, J.-L. The E-Cadherin Cleavage Associated to Pathogenic Bacteria Infections Can Favor Bacterial Invasion and Transmigration, Dysregulation of the Immune Response and Cancer Induction in Humans. Front. Microbiol. 2019, 10, 2598. [Google Scholar] [CrossRef] [PubMed]
- Mehta, S.; Nijhuis, A.; Kumagai, T.; Lindsay, J.; Silver, A. Defects in the adherens junction complex (E-cadherin/ β-catenin) in inflammatory bowel disease. Cell Tissue Res. 2015, 360, 749–760. [Google Scholar] [CrossRef]
- Yi, H.; Hu, J.; Qian, J.; Hackam, A.S. Expression of brain-derived neurotrophic factor is regulated by the Wnt signaling pathway. NeuroReport 2012, 23, 189–194. [Google Scholar] [CrossRef] [Green Version]
- Mallei, A.; Ieraci, A.; Corna, S.; Tardito, D.; Lee, F.S.; Popoli, M. Global epigenetic analysis of BDNF Val66Met mice hippocampus reveals changes in dendrite and spine remodeling genes. Hippocampus 2018, 28, 783–795. [Google Scholar] [CrossRef]
- Bamji, S.X.; Rico, B.; Kimes, N.; Reichardt, L.F. BDNF mobilizes synaptic vesicles and enhances synapse formation by disrupting cadherin–β-catenin interactions. J. Cell Biol. 2006, 174, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Hoseth, E.Z.; Krull, F.; Dieset, I.; Mørch, R.H.; Hope, S.; Gardsjord, E.S.; Steen, N.E.; Melle, I.; Brattbakk, H.-R.; Steen, V.M.; et al. Exploring the Wnt signaling pathway in schizophrenia and bipolar disorder. Transl. Psychiatry 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Dujaili, A.H.; Mousa, R.F.; Al-Hakeim, H.K.; Maes, M. High Mobility Group Protein 1 and Dickkopf-Related Protein 1 in Schizophrenia and Treatment-Resistant Schizophrenia: Associations with Interleukin-6, Symptom Domains, and Neurocognitive Impairments. Schizophr. Bull. 2021, 47, 530–541. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Delanghe, J.; Chiavetto, L.B.; Bignotti, S.; Tura, G.B.; Pioli, R.; Zanardini, R.; Altamura, C.A. Haptoglobin polymorphism and schizophrenia: Genetic variation on chromosome 16. Psychiatry Res. 2001, 104, 1–9. [Google Scholar] [CrossRef]
- Maes, M.; Vojdani, A.; Galecki, P.; Kanchanatawan, B. How to Construct a Bottom-Up Nomothetic Network Model and Disclose Novel Nosological Classes by Integrating Risk Resilience and Adverse Outcome Pathways with the Phenome of Schizophrenia. Brain Sci. 2020, 10, 645. [Google Scholar] [CrossRef]
- Levchenko, A.; Davtian, S.; Freylichman, O.; Zagrivnaya, M.; Kostareva, A.; Malashichev, Y. Beta-catenin in schizophrenia: Possibly deleterious novel mutation. Psychiatry Res. 2015, 228, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Dong, F.; Jiang, J.; McSweeney, C.; Zou, D.; Liu, L.; Mao, Y. Deletion of CTNNB1 in inhibitory circuitry contributes to autism-associated behavioral defects. Hum. Mol. Genet. 2016, 25, 2738–2751. [Google Scholar] [CrossRef] [Green Version]
- Redies, C.; Hertel, N.; Hübner, C.A. Cadherins and neuropsychiatric disorders. Brain Res. 2012, 1470, 130–144. [Google Scholar] [CrossRef]
- Hawi, Z.; Tong, J.; Dark, C.; Yates, H.; Johnson, B.; Bellgrove, M.A. The role of cadherin genes in five major psychiatric disorders: A literature update. Am. J. Med Genet. Part B Neuropsychiatr. Genet. 2018, 177, 168–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connor, L.; Gilmour, J.; Bonifer, C. The Role of the Ubiquitously Expressed Transcription Factor Sp1 in Tissue-specific Tran-scriptional Regulation and in Disease. Yale J. Biol. Med. 2016, 89, 513–525. [Google Scholar]
- Mao, X.R.; Moerman-Herzog, A.M.; Chen, Y.; Barger, S.W. Unique aspects of transcriptional regulation in neurons nuances in NFkappaB and Sp1-related factors. J. Neuroinflammation 2009, 6, 16. [Google Scholar] [CrossRef] [Green Version]
- Sif, S.; Gilmore, T.D. Interaction of the v-Rel oncoprotein with cellular transcription factor Sp1. J. Virol. 1994, 68, 7131–7138. [Google Scholar] [CrossRef] [Green Version]
- Ryu, H.; Lee, J.; Zaman, K.; Kubilis, J.; Ferrante, R.J.; Ross, B.D.; Neve, R.; Ratan, R.R. Sp1 and Sp3 Are Oxidative Stress-Inducible, Antideath Transcription Factors in Cortical Neurons. J. Neurosci. 2003, 23, 3597–3606. [Google Scholar] [CrossRef] [PubMed]
- Ravache, M.; Weber, C.; Merienne, K.; Trottier, Y. Transcriptional Activation of REST by Sp1 in Huntington’s Disease Models. PLoS ONE 2010, 5, e14311. [Google Scholar] [CrossRef]
- Zhao, J.-G.; Zhou, L.; Jin, J.-Y.; Zhao, Z.; Lan, J.; Zhang, Y.-B.; Zhang, Q.-Y.; Gui, J.-F. Antimicrobial activity-specific to Gram-negative bacteria and immune modulation-mediated NF-κB and Sp1 of a medaka β-defensin. Dev. Comp. Immunol. 2009, 33, 624–637. [Google Scholar] [CrossRef] [PubMed]
- Gallego, J.A.; Blanco, E.A.; Morell, C.; Lencz, T.; Malhotra, A.K. Complement component C4 levels in the cerebrospinal fluid and plasma of patients with schizophrenia. Neuropsychopharmacology 2021, 46, 1140–1144. [Google Scholar] [CrossRef] [PubMed]
- Rey, R.; Suaud-Chagny, M.-F.; Bohec, A.-L.; Dorey, J.-M.; D’Amato, T.; Tamouza, R.; Leboyer, M. Overexpression of complement component C4 in the dorsolateral prefrontal cortex, parietal cortex, superior temporal gyrus and associative striatum of patients with schizophrenia. Brain Behav. Immun. 2020, 90, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Purves-Tyson, T.D.; Robinson, K.; Brown, A.M.; Boerrigter, D.; Cai, H.Q.; Weissleder, C.; Owens, S.J.; Rothmond, D.A.; Weickert, C.S. Increased Macrophages and C1qA, C3, C4 Transcripts in the Midbrain of People with Schizophrenia. Front. Immunol. 2020, 11, 2002. [Google Scholar] [CrossRef] [PubMed]
- Talukdar, P.M.; Abdul, F.; Maes, M.; Berk, M.; Venkatasubramanian, G.; Kutty, B.M.; Debnath, M. A proof-of-concept study of maternal immune activation mediated induction of Toll-like receptor (TLR) and inflammasome pathways leading to neuroprogressive changes and schizophrenia-like behaviours in offspring. Eur. Neuropsychopharmacol. 2021, 52, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Talukdar, P.M.; Abdul, F.; Maes, M.; Binu, V.; Venkatasubramanian, G.; Kutty, B.M.; Debnath, M. Maternal Immune Activation Causes Schizophrenia-like Behaviors in the Offspring through Activation of Immune-Inflammatory, Oxidative and Apoptotic Pathways, and Lowered Antioxidant Defenses and Neuroprotection. Mol. Neurobiol. 2020, 57, 1–17. [Google Scholar] [CrossRef]
- Simeonova, D.; Ivanovska, M.; Murdjeva, M.; Carvalho, A.F.; Maes, M. Recognizing the Leaky Gut as a Trans-diagnostic Target for Neuroimmune Disorders Using Clinical Chemistry and Molecular Immunology Assays. Curr. Top. Med. Chem. 2018, 18, 1641–1655. [Google Scholar] [CrossRef]
- Simeonova, D.; Stoyanov, D.; Leunis, J.; Carvalho, A.F.; Kubera, M.; Murdjeva, M.; Maes, M. Increased Serum Immunoglobulin Responses to Gut Commensal Gram-Negative Bacteria in Unipolar Major Depression and Bipolar Disorder Type 1, Especially When Melancholia Is Present. Neurotox. Res. 2019, 37, 338–348. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Mihaylova, I.; Leunis, J.-C. Increased serum IgA and IgM against LPS of enterobacteria in chronic fatigue syndrome (CFS): Indication for the involvement of gram-negative enterobacteria in the etiology of CFS and for the presence of an increased gut–intestinal permeability. J. Affect. Disord. 2007, 99, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Krynicki, C.R.; Dazzan, P.; Pariante, C.M.; Barnes, N.M.; Vincent, R.C.; Roberts, A.; Giordano, A.; Watson, A.; Suckling, J.; Barnes, T.R.; et al. Deconstructing depression and negative symptoms of schizophrenia; differential and longitudinal immune correlates, and response to minocycline treatment. Brain, Behav. Immun. 2021, 91, 498–504. [Google Scholar] [CrossRef]
- Maes, M.; Fišar, Z.; Medina, M.; Scapagnini, G.; Nowak, G.; Berk, M. New drug targets in depression: Inflammatory, cell-mediated immune, oxidative and nitrosative stress, mitochondrial, antioxidant, and neuroprogressive pathways. And new drug candidates—Nrf2 activators and GSK-3 inhibitors. Inflammopharmacology 2012, 20, 127–150. [Google Scholar] [CrossRef]
- Maes, M. Nooit Meer Moe; Paperback, Zorrobooks: Brugge, Belgium, 2011; ISBN 9789461680013. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MCODE Components | GO ID | Biological Term | Log10 (p) |
|---|---|---|---|
| Upregulated genes in FEP, MCODE1 (Biological GO terms) | GO:0032496 | response to lipopolysaccharide | −41.6 |
| GO:0006954 | inflammatory response | −40.8 | |
| GO:0002237 | response to molecule of bacterial origin | −40.8 | |
| Upregulated genes in FEP, MCODE2 (GO, PANTHER, KEGG, WikiPaths) | R-HSA-5357956 | TNFR1-induced NFkappaB signaling pathway | −10.1 |
| R-HSA-73887 | death receptor signaling | −9.8 | |
| R-HSA-75893 | TNF signaling | −9.4 | |
| Downregulated genes in FEP, MCODE1 (Biological GO terms) | GO:0048011 | neurotrophin TRK receptor signaling pathway | −22.5 |
| GO:0007169 | transmembrane receptor protein tyrosine kinase signaling pathway | −22.1 | |
| GO:0032989 | cellular component morphogenesis | −21.8 | |
| Downregulated genes in FEP, MCODE2 (Biological GO terms) | GO:0016246 | RNA interference | −15.6 |
| GO:0035196 | production of miRNAs involved in gene silencing by miRNA | −15.5 | |
| GO:0070918 | production of small RNA involved in gene silencing by RNA | −15.3 | |
| All seed genes in FEP/FES, MCODE1 (Biological GO terms) | GO:0001774 | microglial cell activation | −13.2 |
| GO:0042531 | positive regulation of tyrosine phosphorylation of STAT protein | −12.3 | |
| GO:0042509 | regulation of tyrosine phosphorylation of STAT protein | −11.8 |
| Top Genes | GO Term ID | GO Term Definition | P | p FDR Adjusted | Number of Genes |
|---|---|---|---|---|---|
| Complement factors | |||||
| 1 | GO:0098883 | synapse pruning | 1.00 × 10−10 | 1.41 × 10−6 | 3 |
| 2 | GO:0030449 | regulation of complement activation | 2.02 × 10−10 | 1.11 × 10−3 | 3 |
| 3 | GO:0002920 | regulation of humoral immune response | 3.18 × 10−7 | 1.17 × 10−3 | 3 |
| 4 | GO:0006958 | complement activation, classical pathway | 5.40 × 10−7 | 1.30 × 10−3 | 3 |
| 5 | GO:0002455 | humoral immune response mediated by circulating immunoglobulin | 5.92 × 10−7 | 1.30 × 10−3 | 3 |
| 6 | GO:0006956 | complement activation | 7.29 × 10−7 | 1.34 × 10−3 | 3 |
| 7 | GO:0016064 | immunoglobulin mediated immune response | 1.08 × 10−6 | 1.56 × 10−3 | 3 |
| 8 | GO:0019724 | B cell mediated immunity | 1.13 × 10−6 | 1.56 × 10−3 | 3 |
| 9 | GO:0002449 | lymphocyte mediated immunity | 2.35 × 10−6 | 2.66 × 10−3 | 3 |
| 10 | GO:0050808 | synapse organization | 2.46 × 10−6 | 2.66 × 10−3 | 3 |
| DISC1 | |||||
| 1 | GO:0021799 | cerebral cortex radially oriented cell migration | 1.13 × 10−8 | 1.25 × 10−4 | 3 |
| 2 | GO:0021795 | cerebral cortex cell migration | 3.82 × 10−8 | 1.40 × 10−4 | 3 |
| 3 | GO:0045773 | positive regulation of axon extension | 3.82 × 10−8 | 1.40 × 10−4 | 3 |
| 4 | GO:0022029 | telencephalon cell migration | 9.56 × 10−8 | 2.46 × 10−4 | 3 |
| 5 | GO:0021885 | forebrain cell migration | 1.11 × 10−7 | 2.46 × 10−4 | 3 |
| 6 | GO:0021954 | central nervous system neuron development | 2.54 × 10−7 | 4.67 × 10−4 | 3 |
| 7 | GO:0050772 | positive regulation of axonogenesis | 3.06 × 10−7 | 4.81 × 10−4 | 3 |
| 8 | GO:0030516 | regulation of axon extension | 4.42 × 10−7 | 6.09 × 10−4 | 3 |
| 9 | GO:0061387 | regulation of extent of cell growth | 6.67 × 10−7 | 8.17 × 10−4 | 3 |
| 10 | GO:0021987 | cerebral cortex development | 7.83 × 10−7 | 8.48 × 10−4 | 3 |
| DROSCHA | |||||
| 1 | GO:0031053 | primary miRNA processing | 1.84 × 10−7 | 2.03 × 10−3 | 2 |
| 2 | GO:0010586 | miRNA metabolic process | 1.18 × 10−6 | 5.95 × 10−3 | 2 |
| 3 | GO:0035196 | production of miRNAs involved in gene silencing by miRNA | 2.08 × 10−6 | 5.95 × 10−3 | 2 |
| 4 | GO:0070918 | production of small RNA involved in gene silencing by RNA | 2.70 × 10−6 | 5.95 × 10−3 | 2 |
| 5 | GO:0031050 | dsRNA processing | 2.70 × 10−6 | 5.95 × 10−3 | 2 |
| DOID ID | Disease | Size | Overlap | Enrichment | p-Value |
|---|---|---|---|---|---|
| DOID:2914 | immune system disease | 1.9 k | 89/246 | 3.81 | 2.4 × 10−30 |
| DOID:612 | primary immunodeficiency syndrome | 1.3 k | 70/246 | 4.28 | 2.9 × 10−26 |
| DOID:5295 | intestinal disease | 1.0 k | 61/246 | 4.89 | 9.2 × 10−26 |
| DOID:0060032 | autoimmune disease of the musculoskeletal system | 645 | 48/246 | 6.05 | 4.2x 10−24 |
| DOID:0050589 | inflammatory bowel disease | 306 | 35/246 | 9.30 | 8.7 × 10−24 |
| DOID:417 | autoimmune disease | 1.1 k | 58/246 | 4.42 | 3.0 × 10−22 |
| DOID:9500 | leukocyte disease | 417 | 38/246 | 7.41 | 3.3 × 10−22 |
| DOID:65 | connective tissue disease | 1.8 k | 76/246 | 3.35 | 5.3 × 10−22 |
| DOID:0060180 | Colitis | 237 | 30/246 | 10.29 | 8.8 × 10−22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maes, M.; Plaimas, K.; Suratanee, A.; Noto, C.; Kanchanatawan, B. First Episode Psychosis and Schizophrenia Are Systemic Neuro-Immune Disorders Triggered by a Biotic Stimulus in Individuals with Reduced Immune Regulation and Neuroprotection. Cells 2021, 10, 2929. https://doi.org/10.3390/cells10112929
Maes M, Plaimas K, Suratanee A, Noto C, Kanchanatawan B. First Episode Psychosis and Schizophrenia Are Systemic Neuro-Immune Disorders Triggered by a Biotic Stimulus in Individuals with Reduced Immune Regulation and Neuroprotection. Cells. 2021; 10(11):2929. https://doi.org/10.3390/cells10112929
Chicago/Turabian StyleMaes, Michael, Kitiporn Plaimas, Apichat Suratanee, Cristiano Noto, and Buranee Kanchanatawan. 2021. "First Episode Psychosis and Schizophrenia Are Systemic Neuro-Immune Disorders Triggered by a Biotic Stimulus in Individuals with Reduced Immune Regulation and Neuroprotection" Cells 10, no. 11: 2929. https://doi.org/10.3390/cells10112929
APA StyleMaes, M., Plaimas, K., Suratanee, A., Noto, C., & Kanchanatawan, B. (2021). First Episode Psychosis and Schizophrenia Are Systemic Neuro-Immune Disorders Triggered by a Biotic Stimulus in Individuals with Reduced Immune Regulation and Neuroprotection. Cells, 10(11), 2929. https://doi.org/10.3390/cells10112929