
Remnant of Unrelated Amniote Sex Chromosomal Linkage Sharing on the Same Chromosome in House Gecko Lizards, Providing a Better Understanding of the Ancestral Super-Sex Chromosome





 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimen Collection, Cell Culture, Chromosome Preparation
2.2. Meiotic Chromosome Preparation
2.3. C-Bandings
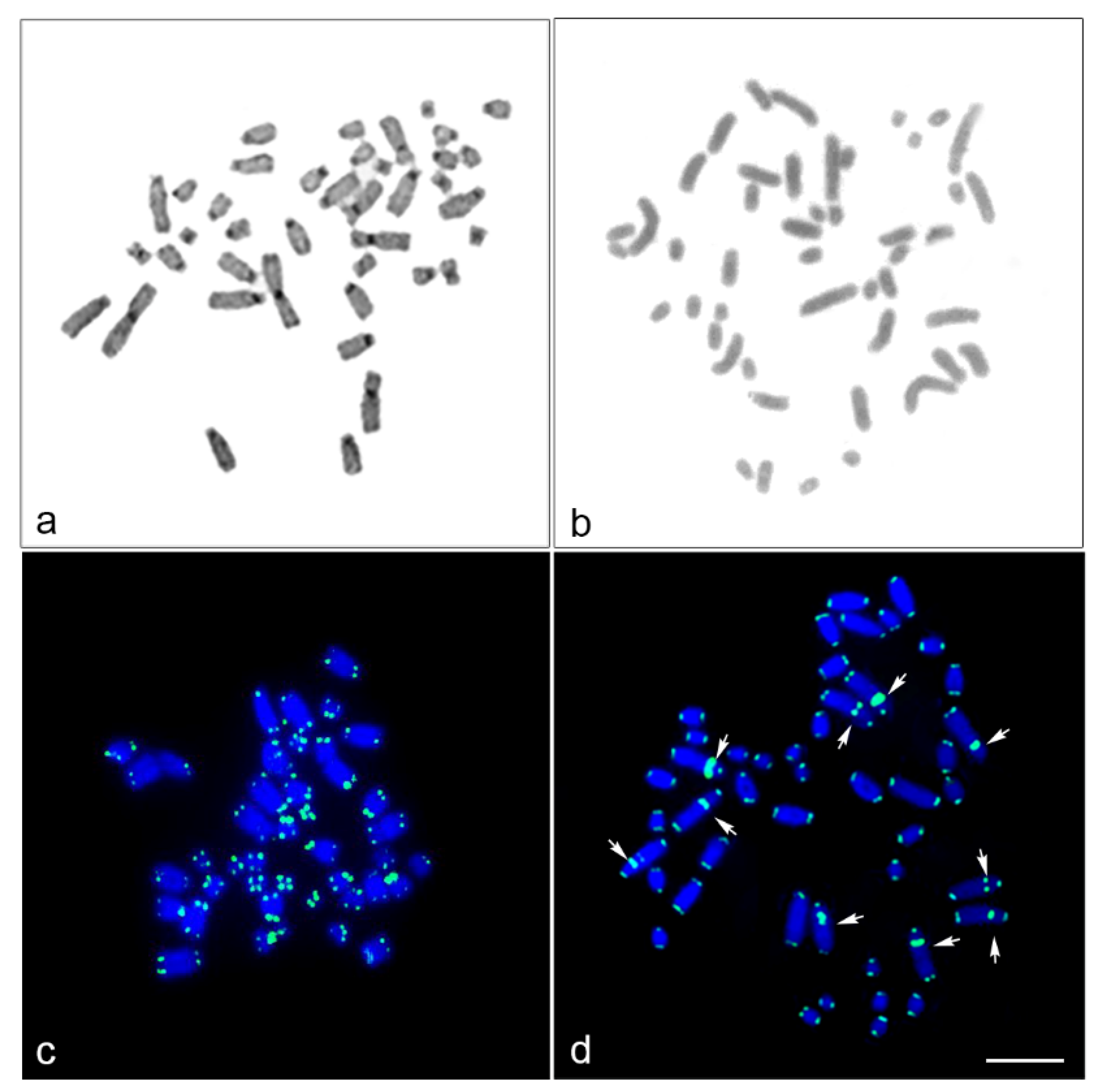
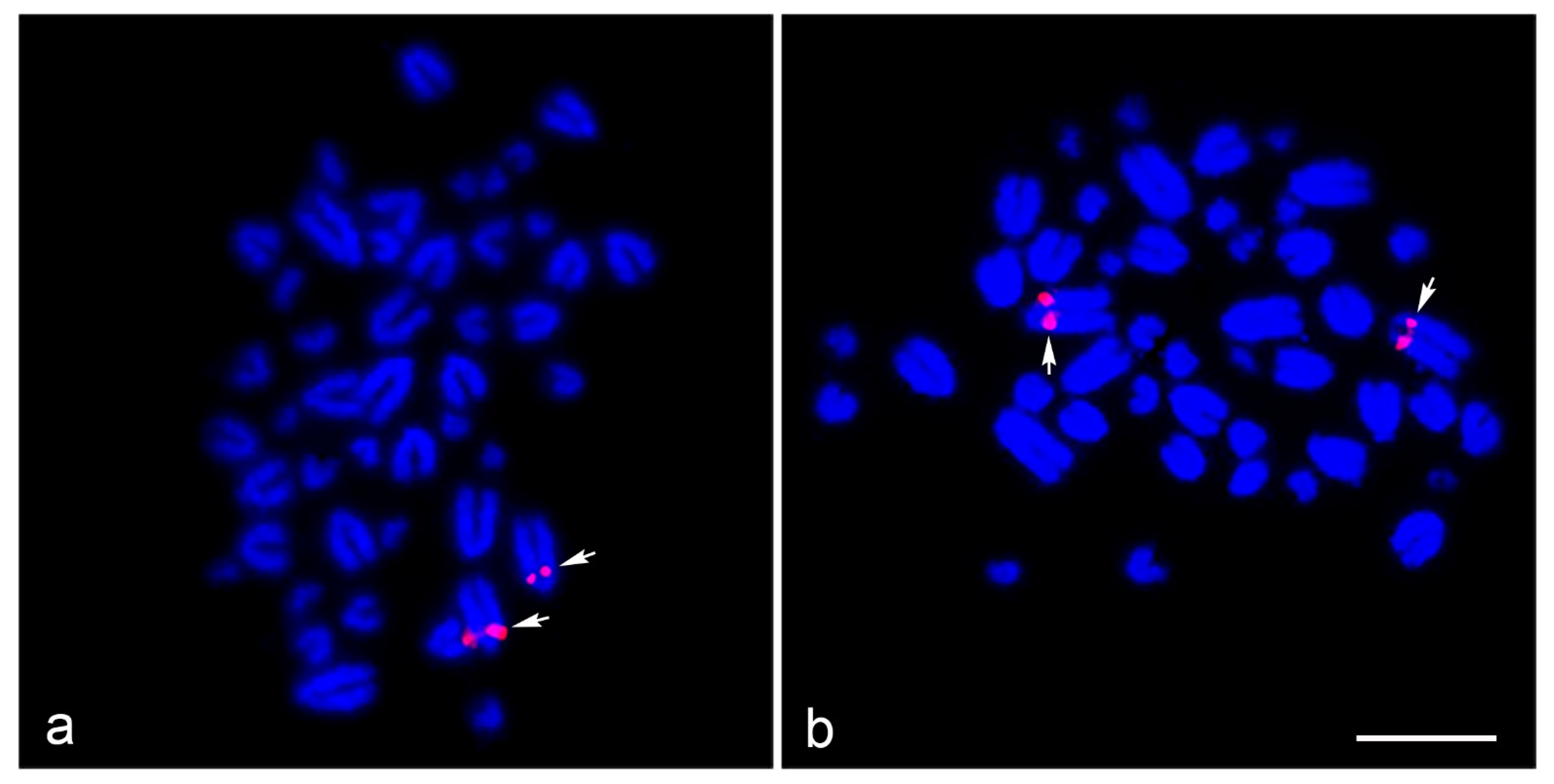
2.4. Fluorescence In Situ Hybridization Mapping of Telomeric Repeat and Microsatellite Repeat Motifs
2.5. Isolation, Amplification and Labeling of Chicken and Zebra Finch BACs
2.6. Cross-Species BAC FISH Mapping
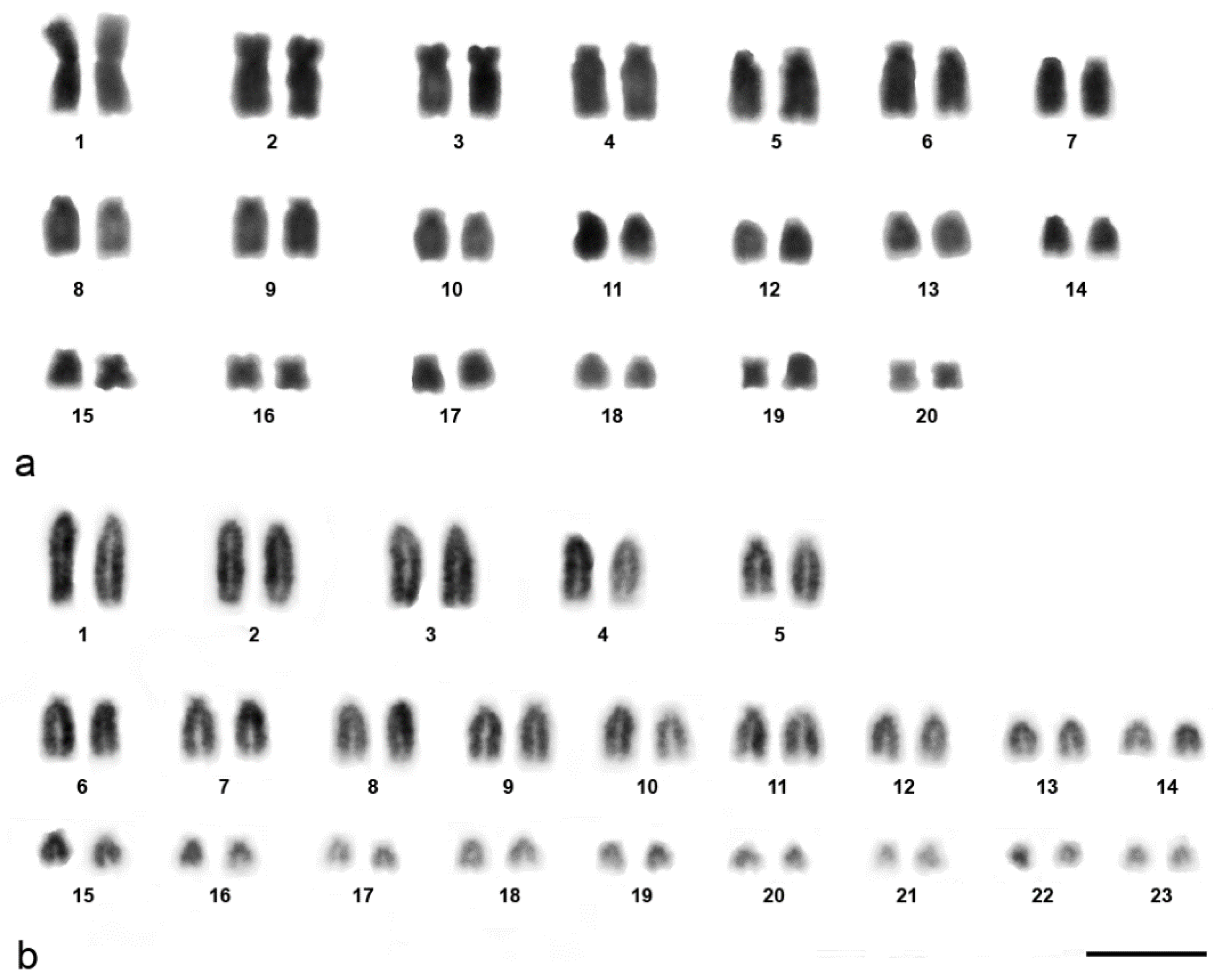
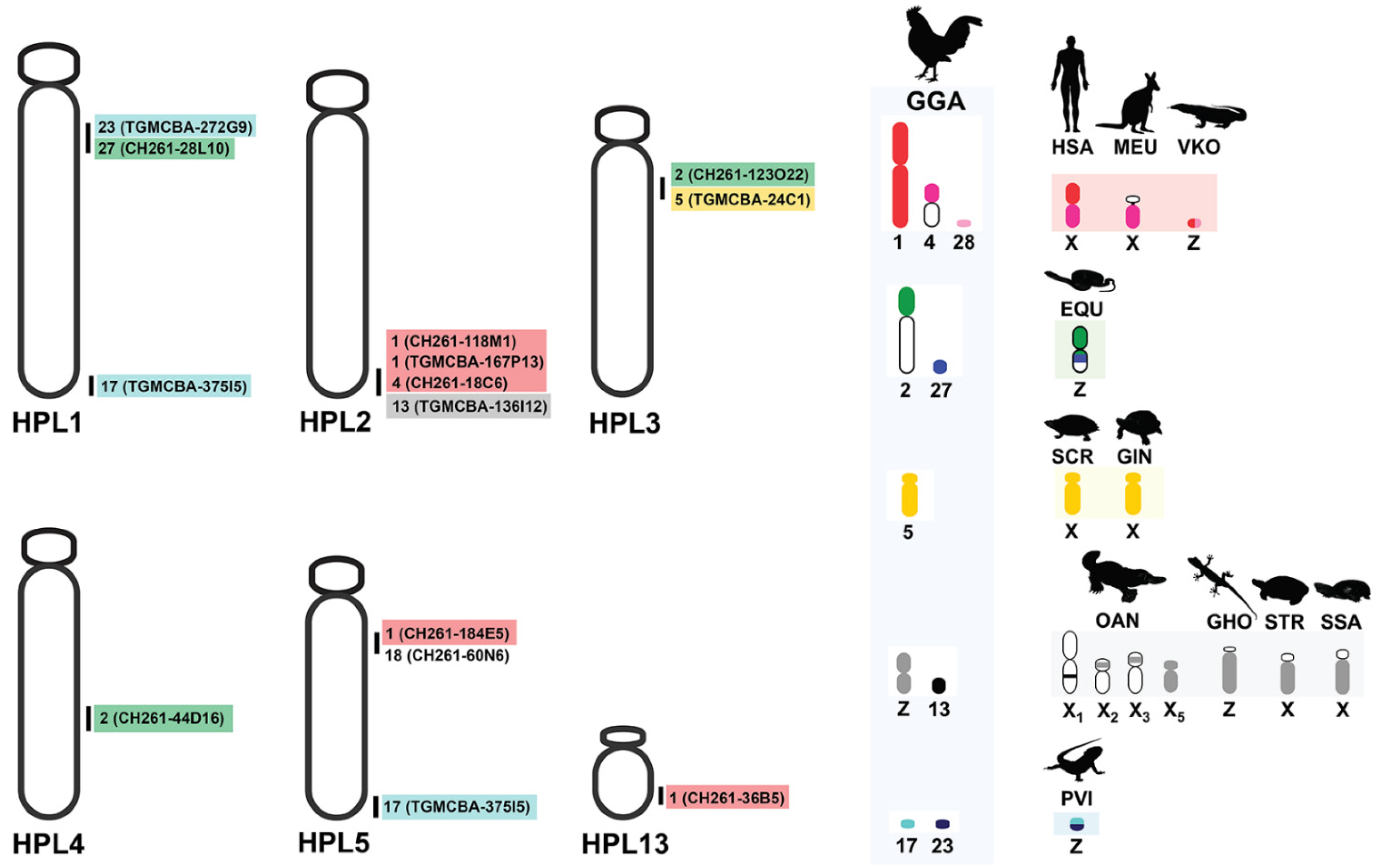
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deng, Z.; Delwart, E. ContigExtender: A new approach to improving de novo sequence assembly for viral metagenomics data. BMC Bioinform. 2021, 22, 119. [Google Scholar] [CrossRef] [PubMed]
- International Chicken Genome Sequencing Consortium. Sequence and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution. Nature 2004, 432, 695–716. [Google Scholar] [CrossRef] [PubMed]
- Schook, L.B.; Beever, J.E.; Rogers, J.; Humphray, S.; Archibald, A.; Chardon, P.; Milan, D.; Rohrer, G.; Eversole, K. Swine Genome Sequencing Consortium (SGSC): A strategic roadmap for sequencing the pig genome. Comp. Funct. Genomics 2005, 6, 251–255. [Google Scholar] [CrossRef] [Green Version]
- Snelling, W.M.; Chiu, R.; Schein, J.E.; Hobbs, M.; Abbey, C.A.; Adelson, D.L.; Aerts, J.; Bennett, G.L.; Bosdet, I.E.; Boussaha, M.; et al. A physical map of the bovine genome. Genome Biol. 2007, 8, R165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghurye, J.; Rhie, A.; Walenz, B.P.; Schmitt, A.; Selvaraj, S.; Pop, M.; Phillippy, A.M.; Koren, S. Integrating Hi-C links with assembly graphs for chromosome-scale assembly. PLoS Comput. Biol. 2019, 15, e1007273. [Google Scholar] [CrossRef] [Green Version]
- Suryamohan, K.; Krishnankutty, S.P.; Guillory, J.; Jevit, M.; Schröder, M.S.; Wu, M.; Kuriakose, B.; Mathew, O.K.; Perumal, R.C.; Koludarov, I.; et al. The Indian cobra reference genome and transcriptome enables comprehensive identification of venom toxins. Nat. Genet. 2020, 52, 106–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Z.; Druzhinina, I.S.; Wang, X.; Zhang, X.; Peng, L.; Labbé, J. Insight into a highly polymorphic endophyte isolated from the roots of the halophytic seepweed Suaeda salsa: Laburnicola rhizohalophila sp. nov. (Didymosphaeriaceae, Pleosporales). Fungal Biol. 2020, 124, 327–337. [Google Scholar] [CrossRef]
- Singchat, W.; O’Connor, R.E.; Tawichasri, P.; Suntronpong, A.; Sillapaprayoon, S.; Suntrarachun, S.; Muangmai, N.; Baicharoen, S.; Peyachoknagul, S.; Chanhome, L.; et al. Chromosome map of the Siamese cobra: Did partial synteny of sex chromosomes in the amniote represent “a hypothetical ancestral super-sex chromosome” or random distribution? BMC Genom. 2018, 19, 1–16. [Google Scholar] [CrossRef]
- Singchat, W.; Sillapaprayoon, S.; Muangmai, N.; Baicharoen, S.; Indananda, C.; Duengkae, P.; Peyachoknagul, S.; O’Connor, R.E.; Griffin, D.K.; Srikulnath, K. Do sex chromosomes of snakes, monitor lizards, and iguanian lizards result from multiple fission of an “ancestral amniote super-sex chromosome”? Chromosome Res. 2020, 28, 209–228. [Google Scholar] [CrossRef]
- Singchat, W.; Ahmad, S.F.; Sillapaprayoon, S.; Muangmai, N.; Duengkae, P.; Peyachoknagul, S.; O’Connor, R.E.; Griffin, D.K.; Srikulnath, K. Partial amniote sex chromosomal linkage homologies shared on snake W sex chromosomes support the ancestral super-sex chromosome evolution in amniotes. Front. Genet. 2020, 11, 948. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Shi, M.; Holmes, E.C. Using metagenomics to characterize an expanding virosphere. Cell 2018, 172, 1168–1172. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.; Wang, Q.; Khurshid, H.; Raza, G.; Han, J.; Wang, B.; Wang, K. Chromosome painting provides insights into the genome structure and evolution of sugarcane. Front. Plant Sci. 2021, 12, 731664. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Hou, Y.; Xu, Y.; Luan, Y.; Zhou, H.; Qi, X.; Hu, M.; Wang, D.; Wang, Z.; Fu, Y.; et al. A compendium and comparative epigenomics analysis of cis-regulatory elements in the pig genome. Nat. Commun. 2021, 12, 2217. [Google Scholar] [CrossRef] [PubMed]
- Deakin, J.E.; Ezaz, T. Understanding the evolution of reptile chromosomes through applications of combined cytogenetics and genomics approaches. Cytogenet. Genome Res. 2019, 157, 7–20. [Google Scholar] [CrossRef]
- Olmo, E. Rate of chromosome changes and speciation in reptiles. Genetica 2005, 125, 185–203. [Google Scholar] [CrossRef]
- Gamble, T.; Bauer, A.M.; Greenbaum, E.; Jackman, T.R. Out of the blue: A novel, trans-atlantic clade of geckos (Gekkota, Squamata). Zool. Scr. 2008, 37, 355–366. [Google Scholar] [CrossRef]
- Pyron, R.A.; Burbrink, F.T.; Wiens, J.J. A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes. BMC Evol. Biol. 2013, 13, 93. [Google Scholar] [CrossRef] [Green Version]
- Pincheira-Donoso, D.; Bauer, A.M.; Meiri, S.; Uetz, P. Global taxonomic diversity of living reptiles. PLoS ONE 2013, 8, e59741. [Google Scholar] [CrossRef] [Green Version]
- Uetz, P.; Freed, P.; Aguilar, R.; Hošek, J. (Eds.) The Reptile Database. 2021. Available online: http://www.reptile-database.org (accessed on 30 August 2021).
- Olmo, E.; Signorino, G. Chromorep: A Reptile Chromosomes Database. 2005. Available online: http://chromorep.univpm.it (accessed on 30 August 2021).
- Kawai, A.; Ishijima, J.; Nishida, C.; Kosaka, A.; Ota, H.; Kohno, S.; Matsuda, Y. The ZW sex chromosomes of Gekko hokouensis (Gekkonidae, Squamata) represent highly conserved homology with those of avian species. Chromosoma 2009, 118, 43–51. [Google Scholar] [CrossRef]
- Trifonov, V.A.; Giovannotti, M.; O’Brien, P.C.; Wallduck, M.; Lovell, F.; Rens, W.; Parise-Maltempi, P.P.; Caputo, V.; Ferguson-Smith, M.A. Chromosomal evolution in Gekkonidae. I. Chromosome painting between Gekko and Hemidactylus species reveals phylogenetic relationships within the group. Chromosome Res. 2011, 19, 843–855. [Google Scholar] [CrossRef]
- Srikulnath, K.; Uno, Y.; Nishida-Umehara, C.; Ota, H.; Matsuda, Y. Karyotype reorganization in the Hokou Gecko (Gekko hokouensis, Gekkonidae): The process of microchromosome disappearance in Gekkota. PLoS ONE 2015, 10, e0134829. [Google Scholar] [CrossRef] [PubMed]
- Srikulnath, K.; Ahmad, S.F.; Singchat, W.; Panthum, T. Why do some vertebrates have microchromosomes? Cells 2021, 10, 2182. [Google Scholar] [CrossRef] [PubMed]
- Schmid, M.; Feichtinger, W.; Nanda, I.; Schakowski, R.; Visbal Garcia, R.; Manzanilla Puppo, J.; Fernández Badillo, A. An extraordinarily low diploid chromosome number in the reptile Gonatodes taniae (Squamata, Gekkonidae). J. Hered. 1994, 85, 255–260. [Google Scholar] [CrossRef]
- De Smet, W.H.O. Description of the orcein stained karyotypes of 36 lizard species (Lacertilia, Reptilia) belonging to the families Teiidae, Scincidae, Lacertidae, Cordylidae and Varanidae (Autrachoglossa). Acta. Zool. Pathol. Antverp. 1981, 76, 37–118. [Google Scholar]
- Gorman, G.C. The chromosome of the Reptilia, a cytotaxonomic interpretation. In Cytotaxonomy and Vertebrate Evolution; Chiarelli, A.B., Cappana, E., Eds.; Academic Press: New York, NY, USA, 1973; pp. 349–424. [Google Scholar]
- Olmo, E. A Reptilia. In Animal cytogenetics 4, Chordata 3; John, B., Ed.; Gebrueder Borntraeger: Berlin, Germany, 1986; pp. 1–100. [Google Scholar]
- Odierna, G.; Aprea, G.; Capriglione, T.; Caputo, V.; Olmo, E. Chromosomal diversification in the gekkonid Tarentola mauritanica (Reptilia, Gekkonidae). Ital. J. Zool. 1994, 61, 325–330. [Google Scholar]
- Trifonov, V.A.; Paoletti, A.; Caputo Barucchi, V.; Kalinina, T.; O’Brien, P.C.; Ferguson-Smith, M.A.; Giovannotti, M. Comparative chromosome painting and NOR distribution suggest a complex hybrid origin of triploid Lepidodactylus lugubris (Gekkonidae). PLoS ONE 2015, 10, e0132380. [Google Scholar] [CrossRef]
- Olmo, E. Trends in the evolution of reptilian chromosomes. Integr. Comp. Biol. 2008, 48, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Srikulnath, K.; Matsubara, K.; Uno, Y.; Nishida, C.; Olsson, M.; Matsuda, Y. Identification of the linkage group of the Z sex chromosomes of the sand lizard (Lacerta agilis, Lacertidae) and elucidation of karyotype evolution in lacertid lizards. Chromosoma 2014, 123, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Chadov, B.F.; Chadova, E.V.; Khotskina, E.A.; Artemova, E.V.; Fedorova, N.B. The main effect of chromosomal rearrangement is changing the action of regulatory genes. Genetika 2004, 40, 893–902. [Google Scholar] [CrossRef]
- Harewood, L.; Fraser, P. The impact of chromosomal rearrangements on regulation of gene expression. Hum. Mol. Genet. 2014, 23, R76–R82. [Google Scholar] [CrossRef] [Green Version]
- Stewart, N.B.; Rogers, R.L. Chromosomal rearrangements as a source of new gene formation in Drosophila yakuba. PLoS Genet. 2019, 15, e1008314. [Google Scholar] [CrossRef] [PubMed]
- Gamble, T.; Coryell, J.; Ezaz, T.; Lynch, J.; Scantlebury, D.P.; Zarkower, D. Restriction Site-Associated DNA Sequencing (RAD-seq) reveals an extraordinary number of transitions among gecko sex-determining systems. Mol. Biol. Evol. 2015, 32, 1296–1309. [Google Scholar] [CrossRef] [Green Version]
- Seipp, R.; Henkel, F.W. Rhacodactylus: Biology. In Natural History & Husbandry; Chimaira: Frankfurt, Germany, 2000. [Google Scholar]
- de Vosjoli, P.; Fast, F.; Repashy, A. Rhacodactylus: The Complete Guide to Their Selection and Care; Advanced Visions, Inc.: Vista, CA, USA, 2003. [Google Scholar]
- Gamble, T. A review of sex determining mechanisms in geckos (Gekkota: Squamata). Sex Dev. 2010, 4, 88–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorman, G.C.; Gress, F. Sex chromosomes of a pygopodid lizard, Lialis burtonis. Experientia 1970, 26, 206–207. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, K.; Knopp, T.; Sarre, S.D.; Georges, A.; Ezaz, T. Karyotypic analysis and FISH mapping of microsatellite motifs reveal highly differentiated XX/XY sex chromosomes in the pink-tailed worm-lizard (Aprasia parapulchella, Pygopodidae, Squamata). Mol. Cytogenet. 2013, 6, 60. [Google Scholar] [CrossRef] [Green Version]
- Pokorná, M.; Kratochvíl, L.; Kejnovský, E. Microsatellite distribution on sex chromosomes at different stages of heteromorphism and heterochromatinization in two lizard species (Squamata: Eublepharidae: Coleonyx elegans and Lacertidae: Eremias velox). BMC Genet. 2011, 12, 90. [Google Scholar] [CrossRef] [Green Version]
- Matsubara, K.; Sarre, S.D.; Georges, A.; Matsuda, Y.; Marshall Graves, J.A.; Ezaz, T. Highly differentiated ZW sex microchromosomes in the Australian varanus species evolved through rapid amplification of repetitive sequences. PLoS ONE 2014, 9, e95226. [Google Scholar] [CrossRef]
- Rovatsos, M.; Kratochvíl, L. Molecular sexing applicable in 4000 species of lizards and snakes? From dream to real possibility. Methods Ecol Evol. 2017, 8, 902–906. [Google Scholar] [CrossRef] [Green Version]
- Mazzoleni, S.; Němec, P.; Albrecht, T.; Lymberakis, P.; Kratochvíl, L.; Rovatsos, M. Long-term stability of sex chromosome gene content allows accurate qPCR-based molecular sexing across birds. Mol. Ecol. Resour. 2021, 21, 2013–2021. [Google Scholar] [CrossRef]
- Pokorna, M.; Kratochvil, L. Phylogeny of sex-determining mechanisms in squamate reptiles: Are sex chromosomes an evolutionary trap? Zool. J Linn. Soc. 2009, 156, 168–183. [Google Scholar] [CrossRef] [Green Version]
- Keating, S.E.; Blumer, M.; Grismer, L.L.; Lin, A.; Nielsen, S.V.; Thura, M.K.; Wood, P.L.; Quah, E., Jr.; Gamble, T. Sex chromosome turnover in bent-toed geckos (Cyrtodactylus). Genes 2021, 12, 116. [Google Scholar] [CrossRef] [PubMed]
- Ezaz, T.; Srikulnath, K.; Graves, J.A. Origin of amniote sex chromosomes: An ancestral super-sex chromosome, or common requirements? J. Hered. 2017, 108, 94–105. [Google Scholar] [CrossRef]
- Singchat, W.; Ahmad, S.F.; Laopichienpong, N.; Suntronpong, A.; Panthum, T.; Griffin, D.K.; Srikulnath, K. Snake W sex chromosome: The shadow of ancestral amniote super-sex chromosome. Cells 2020, 9, 2386. [Google Scholar] [CrossRef]
- Matsubara, K.; O’Meally, D.; Sarre, S.D.; Georges, A.; Srikulnath, K.; Ezaz, T. ZW sex chromosomes in Australian dragon lizards (Agamidae) originated from a combination of duplication and translocation in the nucleolar organising region. Genes 2019, 10, 861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, S.F.; Singchat, W.; Jehangir, M.; Panthum, T.; Srikulnath, K. Consequence of paradigm shift with repeat landscapes in reptiles: Powerful facilitators of chromosomal rearrangements for diversity and evolution. Genes 2020, 11, 827. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.F.; Singchat, W.; Panthum, T.; Srikulnath, K. Impact of repetitive DNA elements on snake genome biology and evolution. Cells 2021, 10, 1707. [Google Scholar] [CrossRef]
- Pope, C.H. Four new snakes and a new lizard from South China. American Museum Novitates 1928, 325, 1–4. [Google Scholar]
- Linnaeus, C. Systema Naturæ per Regna tria Naturæ, Secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentiis, Synonymis, Locis. Tomus I. Editio Decima, Reformata, 10th ed.; Laurentii Salvii: Holmiæ, Cilicia, 1758; p. 824. [Google Scholar]
- Koomgun, T.; Laopichienpong, N.; Singchat, W.; Panthum, T.; Phatcharakullawarawat, R.; Kraichak, E.; Sillapaprayoon, S.; Ahmad, S.F.; Muangmai, N.; Peyachoknagul, S.; et al. Genome complexity reduction high-throughput genome sequencing of green iguana (Iguana iguana) reveal a paradigm shift in understanding sex-chromosomal linkages on homomorphic X and Y sex chromosomes. Front. Genet. 2020, 11, 556267. [Google Scholar] [CrossRef]
- Laopichienpong, N.; Kraichak, E.; Singchat, W.; Sillapaprayoon, S.; Muangmai, N.; Suntrarachun, S.; Baicharoen, S.; Peyachoknagul, S.; Chanhome, L.; Ezaz, T.; et al. Genome-wide SNP analysis of Siamese cobra (Naja kaouthia) reveals the molecular basis of transitions between Z and W sex chromosomes and supports the presence of an ancestral super-sex chromosome in amniotes. Genomics 2021, 113, 624–636. [Google Scholar] [CrossRef]
- Matsuda, Y.; Nishida-Umehara, C.; Tarui, H.; Kuroiwa, A.; Yamada, K.; Isobe, T.; Ando, J.; Fujiwara, A.; Hirao, Y.; Nishimura, O.; et al. Highly conserved linkage homology between birds and turtles: Bird and turtle chromosomes are precise counterparts of each other. Chromosome Res. 2005, 13, 601–615. [Google Scholar] [CrossRef]
- Grützner, F.; Rens, W.; Tsend-Ayush, E.; El-Mogharbel, N.; O’Brien, P.C.; Jones, R.C.; Ferguson-Smith, M.A.; Marshall Graves, J.A. In the platypus a meiotic chain of ten sex chromosomes shares genes with the bird Z and mammal X chromosomes. Nature 2004, 432, 913–917. [Google Scholar] [CrossRef]
- Veyrunes, F.; Waters, P.D.; Miethke, P.; Rens, W.; McMillan, D.; Alsop, A.E.; Grützner, F.; Deakin, J.E.; Whittington, C.M.; Schatzkamer, K.; et al. Bird-like sex chromosomes of platypus imply recent origin of mammal sex chromosomes. Genome Res. 2008, 18, 965–973. [Google Scholar] [CrossRef] [Green Version]
- Alföldi, J.; Palma, F.D.; Grabherr, M. The genome of the green anole lizard and a comparative analysis with birds and mammals. Nature 2011, 477, 587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezaz, T.; Azad, B.; O’Meally, D.; Young, M.J.; Matsubara, K.; Edwards, M.J.; Zhang, X.; Holleley, C.E.; Deakin, J.E.; Marshall Graves, J.A.; et al. Sequence and gene content of a large fragment of a lizard sex chromosome and evaluation of candidate sex differentiating gene R-spondin 1. BMC Genom. 2013, 14, 899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srikulnath, K.; Matsubara, K.; Uno, Y.; Thongpan, A.; Suputtitada, S.; Apisitwanich, S.; Matsuda, Y.; Nishida, C. Karyological characterization of the butterfly lizard (Leiolepis reevesii rubritaeniata, Agamidae, Squamata) by molecular cytogenetic approach. Cytogenet. Genome Res. 2009, 125, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Srikulnath, K.; Nishida, C.; Matsubara, K.; Uno, Y.; Thongpan, A.; Suputtitada, S.; Apisitwanich, S.; Matsuda, Y. Karyotypic evolution in squamate reptiles: Comparative gene mapping revealed highly conserved linkage homology between the butterfly lizard (Leiolepis reevesii rubritaeniata, Agamidae, Lacertilia) and the Japanese four-striped rat snake (Elaphe quadrivirgata, Colubridae, Serpentes). Chromosome Res. 2009, 17, 975–986. [Google Scholar] [CrossRef]
- Srikulnath, K.; Uno, Y.; Nishida, C.; Matsuda, Y. Karyotype evolution in monitor lizards: Cross-species chromosome mapping of cDNA reveals highly conserved synteny and gene order in the Toxicofera clade. Chromosome Res. 2013, 21, 805–819. [Google Scholar] [CrossRef]
- Lind, A.L.; Lai, Y.; Mostovoy, Y.; Holloway, A.K.; Iannucci, A.; Mak, A.; Fondi, M.; Orlandini, V.; Eckalbar, W.L.; Milan, M.; et al. Genome of the Komodo dragon reveals adaptations in the cardiovascular and chemosensory systems of monitor lizards. Nat. Ecol. Evol. 2019, 3, 1241–1252. [Google Scholar] [CrossRef] [Green Version]
- Matsubara, K.; Tarui, H.; Toriba, M.; Yamada, K.; Nishida-Umehara, C.; Agata, K.; Matsuda, Y. Evidence for different origin of sex chromosomes in snakes, birds, and mammals and step-wise differentiation of snake sex chromosomes. Proc. Natl. Acad. Sci. USA 2006, 103, 18190–18195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawagoshi, T.; Nishida, C.; and Matsuda, Y. The origin and differentiation process of X and Y chromosomes of the black marsh turtle (Siebenrockiella crassicollis, Geoemydidae, Testudines). Chromosome Res. 2012, 20, 95–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montiel, E.E.; Badenhorst, D.; Tamplin, J.; Burke, R.L.; Valenzuela, N. Discovery of the youngest sex chromosomes reveals first case of convergent co-option of ancestral autosomes in turtles. Chromosoma 2016, 126, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Kawagoshi, T.; Uno, Y.; Nishida, C.; Matsuda, Y. The Staurotypus turtles and aves share the same origin of sex chromosomes but evolved different types of heterogametic sex determination. PLoS ONE 2014, 9, e105315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badenhorst, D.; Stanyon, R.; Engstrom, T.; Valenzuela, N. A ZZ/ZW microchromosome system in the spiny softshell turtle, Apalone spinifera, reveals an intriguing sex chromosome conservation in Trionychidae. Chromosome Res. 2013, 21, 137–147. [Google Scholar] [CrossRef]
- Kawagoshi, T.; Uno, Y.; Matsubara, K.; Matsuda, Y.; Nishida, C. The ZW micro-sex chromosomes of the Chinese soft-shelled turtle (Pelodiscus sinensis, Trionychidae, Testudines) have the same origin as chicken chromosome 15. Cytogenet. Genome Res. 2009, 125, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Duméril, A.M.C.; Bibron, G. Erpetologie Générale ou Histoire Naturelle Complete des Reptiles; Roret: Paris, France, 1836; Volume 3, p. 528. [Google Scholar]
- Schneider, J.G. Amphibiorum physiologiae specimen alterum historiam et species generis stellionum seu geckonum sistens. Traiecti ad Viadrum Frankfurt/Oder CLF Aitzi. 1792, 2, 54. [Google Scholar]
- Damas, J.; O’Connor, R.; Farré, M.; Lenis, V.P.E.; Martell, H.J.; Mandawala, A.; Fowler, K.; Joseph, S.; Swain, M.T.; Griffin, D.K.; et al. Upgrading short-read animal genome assemblies to chromosome level using comparative genomics and a universal probe set. Genome Res. 2017, 27, 875–884. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, R.E.; Farré, M.; Joseph, S.; Damas, J.; Kiazim, L.; Jennings, R.; Bennett, S.; Slack, E.A.; Allanson, E.; Larkin, D.M.; et al. Chromosome-level assembly reveals extensive rearrangement in saker falcon and budgerigar, but not ostrich, genomes. Genome Biol. 2018, 19, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connor, R.E.; Romanov, M.N.; Kiazim, L.G.; Barrett, P.M.; Farré, M.; Damas, J.; Ferguson-Smith, M.; Valenzuela, N.; Larkin, D.M.; Griffin, D.K. Reconstruction of the diapsid ancestral genome permits chromosome evolution tracing in avian and non-avian dinosaurs. Nat. Commun. 2018, 9, 1883. [Google Scholar] [CrossRef] [Green Version]
- Ezaz, T.; Deakin, J.E. Repetitive sequence and sex chromosome evolution in vertebrates. Adv. Evol. Biol. 2014, 2014, 104683. [Google Scholar] [CrossRef] [Green Version]
- Chalopin, D.; Naville, M.; Plard, F.; Galiana, D.; Volff, J.N. Comparative analysis of transposable elements highlights mobilome diversity and evolution in vertebrates. Genome Biol. Evol. 2015, 7, 567–580. [Google Scholar] [CrossRef]
- Uno, Y.; Nishida, C.; Tarui, H.; Ishishita, S.; Takagi, C.; Nishimura, O.; Ishijima, J.; Ota, H.; Kosaka, A.; Matsubara, K.; et al. Inference of the protokaryotypes of amniotes and tetrapods and the evolutionary processes of microchromosomes from comparative gene mapping. PLoS ONE 2012, 7, e53027. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.K.P.; Lim, F.L.K. A Guide to the Amphibians and Reptiles of Singapore; Singapore Science Centre: Singapore, 1992; pp. 1–160. [Google Scholar]
- Cox, M.J.; van Dijk, P.P.; Nabhitabhata, J.; Thirakhupt, K. A Photographic Guide to Snakes and other Reptiles of Peninsular Malaysia, Singapore and Thailand; New Holland Publishers: London, UK; Cape Town, South Africa; Sydney, Australia; Singapore, 1998; pp. 1–144. [Google Scholar]
- Levan, A.; Fredga, K.; Sandberg, A. Nomenclature for centromeric position on chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- Turpin, R.; Lejeune, J. Les Chromosomes Humains; Gauthier-Villars: Paris, France, 1965. [Google Scholar]
- Imai, H.T.; Matsuda, Y.; Shiroishi, T.; Moriwaki, K. High frequency fo X-Y chromosome dissociation in primary spermatocytes of F1 hybrids between Japanese wild mice (Mus musculus molossinus) and inbred laboratory mice. Cytogenet. Cell Genet. 1981, 29, 166–175. [Google Scholar] [CrossRef]
- Sumner, A.T. A simple technique for demonstrating centromeric heterochromatin. Exp Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef]
- Matsuda, Y.; Chapman, V.M. Application of fluorescence in situ hybridization in genome analysis of the mouse. Electrophoresis 1995, 16, 261–272. [Google Scholar] [CrossRef]
- Griffin, D.K.; Robertson, L.B.; Tempest, H.G.; Vignal, A.; Fillon, V.; Crooijmans, R.P.; Groenen, M.A.; Deryusheva, S.; Gaginskaya, E.; Carré, W.; et al. Whole genome comparative studies between chicken and turkey and their implications for avian genome evolution. BMC Genom. 2008, 9, 168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skinner, B.M.; Robertson, L.B.; Tempest, H.G.; Langley, E.J.; Ioannou, D.; Fowler, K.E.; Crooijmans, R.P.; Hall, A.D.; Griffin, D.K.; Völker, M. Comparative genomics in chicken and Pekin duck using FISH mapping and microarray analysis. BMC Genom. 2009, 10, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lithgow, P.E.; O’Connor, R.; Smith, D.; Fonseka, G.; Al Mutery, A.; Rathje, C.; Frodsham, R.; O’Brien, P.; Kasai, F.; Ferguson-Smith, M.A.; et al. Novel tools for characterising inter and intra chromosomal rearrangements in avian microchromosomes. Chromosome Res. 2014, 22, 85–97. [Google Scholar] [CrossRef]
- King, M. Chromosomal evolution in the Diplodactylinae (Gekkonidae: Reptilia). I. Evolutionary relationships and patterns of change. Aust. J Zool. 1987, 35, 507–531. [Google Scholar] [CrossRef]
- Taeniopygia guttata (Vieillot, 1817) in GBIF Secretariat. GBIF Backbone Taxonomy. Checklist Dataset. 2021. Available online: https://doi.org/10.15468/39omei (accessed on 3 September 2021).
- Stejneger, L.H. Herpetology of Japan and adjacent territory. Bull. US Natl. Mus. 1907, 58, 1–577. [Google Scholar] [CrossRef]
- Hara, Y.; Takeuchi, M.; Kageyama, Y.; Tatsumi, K.; Hibi, M.; Kiyonari, H.; Kuraku, S. Madagascar ground gecko genome analysis characterizes asymmetric fates of duplicated genes. BMC Biol. 2018, 16, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pensabene, E.; Kratochvíl, L.; Rovatsos, M. Independent evolution of sex chromosomes in Eublepharid geckos, a lineage with environmental and genotypic sex determination. Life 2020, 10, 342. [Google Scholar] [CrossRef] [PubMed]
- Augstenová, B.; Pensabene, E.; Veselý, M.; Kratochvíl, L.; Rovatsos, M. Are geckos special in sex determination? Independently evolved differentiated ZZ/ZW sex chromosomes in carphodactylid geckos. Genome Biol. Evol. 2021, 13, evab119. [Google Scholar] [CrossRef] [PubMed]
- Ahl, E. Neue Eidechsen und Amphibien. Zool. Anz. 1926, 67, 186–192. [Google Scholar]
- Young, M.J.; O’Meally, D.; Sarre, S.D.; Georges, A.; Ezaz, T. Molecular cytogenetic map of the central bearded dragon, Pogona vitticeps (Squamata: Agamidae). Chromosome Res. 2013, 21, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, K.; Uno, Y.; Srikulnath, K.; Matsuda, Y.; Miller, E.; Olsson, M. No interstitial telomeres on autosomes but remarkable amplification of telomeric repeats on the W sex chromosome in the sand lizard (Lacerta agilis). J. Hered. 2015, 106, 753–757. [Google Scholar] [CrossRef] [Green Version]
- Matsubara, K.; O’Meally, D.; Azad, B.; Georges, A.; Sarre, S.D.; Graves, J.A.; Matsuda, Y.; Ezaz, T. Amplification of microsatellite repeat motifs is associated with the evolutionary differentiation and heterochromatinization of sex chromosomes in Sauropsida. Chromosoma 2016, 125, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, K.; Nishida, C.; Matsuda, Y.; Kumazawa, Y. Sex chromosome evolution in snakes inferred from divergence patterns of two gametologous genes and chromosome distribution of sex chromosome-linked repetitive sequences. Zool. Lett. 2016, 2, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rovatsos, M.; Pokorná, J.M.; Kratochvíl, L. Differentiation of sex chromosomes and karyotype characterisation in the dragonsnake Xenodermus javanicus (Squamata: Xenodermatidae). Cytogenet. Genome Res. 2015, 147, 48–54. [Google Scholar] [CrossRef]
- Augstenová, B.; Mazzoleni, S.; Kratochvíl, L.; Rovatsos, M. Evolutionary dynamics of the W chromosome in caenophidian snakes. Genes 2018, 9, 5. [Google Scholar] [CrossRef] [Green Version]
- Perry, B.W.; Card, D.C.; McGlothlin, J.W.; Pasquesi, G.; Adams, R.H.; Schield, D.R.; Hales, N.R.; Corbin, A.B.; Demuth, J.P.; Hoffmann, F.G.; et al. Molecular adaptations for sensing and securing prey and insight into amniote genome diversity from the garter snake genome. Genome Biol. Evol. 2018, 10, 2110–2129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamble, T.; Castoe, T.A.; Nielsen, S.V.; Banks, J.L.; Card, D.C.; Schield, D.R.; Schuett, G.W.; Booth, W. The discovery of XY sex chromosomes in a Boa and Python. Curr. Biol. 2017, 27, 2148–2153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graves, J.A.M.; Peichel, C.L. Are homologies in vertebrate sex determination due to shared ancestry or to limited options? Genome Biol. 2010, 11, 205. [Google Scholar] [CrossRef]
- O’Meally, D.; Ezaz, T.; Georges, A.; Sarre, S.D.; Graves, J.A. Are some chromosomes particularly good at sex? Insights from amniotes. Chromosome Res. 2012, 20, 7–19. [Google Scholar] [CrossRef]
- Rovatsos, M.; Farkačová, K.; Altmanová, M.; Johnson Pokorná, M.; Kratochvíl, L. The rise and fall of differentiated sex chromosomes in geckos. Mol. Ecol. 2019, 28, 3042–3052. [Google Scholar] [CrossRef] [PubMed]
- Pokorná, J.M.; Kratochvíl, L. What was the ancestral sex-determining mechanism in amniote vertebrates? Biol. Rev. Camb. Philos. Soc. 2016, 91, 1–12. [Google Scholar] [CrossRef]
- Kostmann, A.; Kratochvíl, L.; Rovatsos, M. Poorly differentiated XX/XY sex chromosomes are widely shared across skink radiation. BioRxiv. 2020. [Google Scholar] [CrossRef]
- Vicoso, B.; Emerson, J.J.; Zektser, Y.; Mahajan, S.; Bachtrog, D. Comparative sex chromosome genomics in snakes: Differentiation, evolutionary strata, and lack of global dosage compensation. PLoS Biol. 2013, 11, e1001643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamble, T.; Geneva, A.J.; Glor, R.E.; Zarkower, D. Anolis sex chromosomes are derived from a single ancestral pair. Evolution 2014, 68, 1027–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charlesworth, B. The evolution of sex chromosomes. Science 1991, 251, 1030–1033. [Google Scholar] [CrossRef]
- Hillis, D.M.; Green, D.M. Evolutionary changes of heterogametic sex in the phylogenetic history of amphibians. J. Evol. Biol. 1990, 3, 49–64. [Google Scholar] [CrossRef]
- Mank, J.E.; Promislow, D.E.L.; Avise, J.C. Evolution of alternative sex-determining mechanisms in teleost fishes. Biol. J. Linn. Soc. 2006, 87, 83–93. [Google Scholar] [CrossRef] [Green Version]
- Takehana, Y.; Naruse, K.; Hamaguchi, S.; Sakaizumi, M. Evolution of ZZ/ZW and XX/XY sex-determination systems in the closely related medaka species, Oryzias hubbsi and O. dancena. Chromosoma 2007, 116, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Volff, J.N.; Nanda, I.; Schmid, M.; Schartl, M. Governing sex determination in fish: Regulatory putsches and ephemeral dictators. Sex. Dev. 2007, 1, 85–99. [Google Scholar] [CrossRef]
- Miura, I. An evolutionary witness: The frog rana rugosa underwent change of heterogametic sex from XY male to ZW female. Sex. Dev. 2007, 1, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.A.; Urton, J.R.; Boland, J.; Shapiro, M.D.; Peichel, C.L. Turnover of sex chromosomes in the stickleback fishes (gasterosteidae). PLoS Genet. 2009, 5, e1000391. [Google Scholar] [CrossRef] [Green Version]
- Graves, J.A.M. Weird animal genomes and the evolution of vertebrate sex and sex chromosomes. Annu. Rev. Genet. 2008, 42, 565–586. [Google Scholar] [CrossRef]
- Evans, B.J.; Pyron, R.A.; Wiens, J.J. Polyploidization and sex chromosome evolution in amphibians. In Polyploidy and Genome Evolution; Soltis, P.S., Soltis, D.E., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 385–410. [Google Scholar]
- Blaser, O.; Neuenschwander, S.; Perrin, N. Sex-chromosome turnovers: The hot-potato model. Am. Nat. 2014, 183, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Bertho, S.; Herpin, A.; Branthonne, A.; Jouanno, E.; Yano, A.; Nicol, B.; Muller, T.; Pannetier, M.; Pailhoux, E.; Miwa, M.; et al. The unusual rainbow trout sex determination gene hijacked the canonical vertebrate gonadal differentiation pathway. Proc. Natl. Acad. Sci. USA 2018, 115, 12781–12786. [Google Scholar] [CrossRef] [Green Version]
- Moore, E.C.; Roberts, R.B. Polygenic sex determination. Curr. Biol. 2013, 23, R510–R512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qvarnström, A.; Bailey, R.I. Speciation through evolution of sex-linked genes. Heredity 2009, 102, 4–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, J.E. Catalogue of the Specimens of Lizards in the Collection of the British Museum; Trustees of die British Museum/Edward Newman: London, UK, 1845; p. 289. [Google Scholar]
- Lambert, M.R.; Smylie, M.S.; Roman, A.J.; Freidenburg, L.K.; Skelly, D.K. Sexual and somatic development of wood frog tadpoles along a thermal gradient. J. Exp. Zool. A Ecol. Integr. Physiol. 2018, 329, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Sarre, S.D.; Georges, A.; Quinn, A. The ends of a continuum: Genetic and temperature-dependent sex determination in reptiles. Bioessays 2004, 26, 639–645. [Google Scholar] [CrossRef]
- Graves, J.A.M. Did sex chromosome turnover promote divergence of the major mammal groups? De novo sex chromosomes and drastic rearrangements may have posed reproductive barriers between monotremes, marsupials and placental mammals. BioEssays 2016, 38, 734–743. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Chromosome Number | Sex Determination | Collection Locality | Number of Animals Used (Female + Male) | Reference |
|---|---|---|---|---|---|
| Hemidactylus frenatus | 2n = 40 | ZZ/ZW | Bangkok | 1 + 0 | [22,36,90] |
| Hemidactylus platyurus | 2n = 46 | - | Bangkok | 1 unknown | [22] |
| GGA | Name | Result | GJA | |
|---|---|---|---|---|
| HFR | HPL | |||
| 1q | CH261-107E2 | - | - | |
| 1q | CH261-118M1 | - | 2 | 4 |
| 1q | CH261-168O17 | - | - | |
| 1q | CH261-184E5 | - | 5 | 2 |
| 1q | CH261-18J16 | - | - | |
| 1q | CH261-58K12 | - | - | |
| 1 | CH261-36B5 | - | 13 | 2p, 11, 13 |
| 1 | TGMCBA-167P13 | - | 2 | 4 |
| 1p | CH261-89C18 | - | - | |
| 1q | CH261-98G4 | - | - | |
| 2p | CH261-123O22 | - | 3 | 1q |
| 2p | CH261-177K1 | - | - | |
| 2p | CH261-169N6 | - | - | |
| 2q | CH261-44D16 | - | 4 | 8 |
| 4 | CH261-18C6 | - | 2 | 4 |
| 4 | CH261-71L6 | - | - | |
| 4p | CH261-83E1 | - | - | |
| 4q | CH261-89P6 | - | - | |
| 5 | CH261-122F8 | - | - | |
| 5 | CH261-2I23 | - | - | |
| 5p | CH261-49B22 | - | - | |
| 5q | CH261-78F13 | - | - | |
| 5 | TGMCBA-145C6 | - | - | |
| 5 | TGMCBA-24C1 | - | 3 | 1q |
| 6q | CH261-49F3 | - | - | |
| 6p | TGMCBA-382J4 | - | - | |
| 9p | CH261-183N19 | - | - | |
| 9q | CH261-187M16 | - | - | |
| 9 | CH261-68O18 | - | - | |
| 9 | CH261-95N3 | - | - | |
| 9 | TGMCBA-150E19 | - | - | |
| 9 | TGMCBA-217A3 | - | - | |
| 9 | TGMCBA-321L6 | - | - | |
| 13p | CH261-115I12 | - | - | |
| 13 | CH261-11H24 | - | - | |
| 13 | CH261-59M8 | - | - | |
| 13 | TGMCBA-136I12 | - | 2 | 4 |
| 13 | TGMCBA-266O5 | - | - | |
| 13q | TGMCBA-321B13 | - | - | |
| 15 | CH261-40D6 | - | - | |
| 15 | CH261-48M1 | - | - | |
| 15p | CH261-90P23 | - | - | |
| 15 | TGMCBA-231D20 | - | - | |
| 15q | TGMCBA-266G23 | - | - | |
| 17 | CH261-113A7 | - | - | |
| 17q | CH261-42P16 | - | - | |
| 17 | CH261-69M11 | - | - | |
| 17 | TGMCBA-185B22 | - | - | |
| 17 | TGMCBA-197G19 | - | - | |
| 17p | TGMCBA-375I5 | - | 1 and 5 | 3 and 2q |
| 17 | TGMCBA-67H23 | - | - | |
| 18p | CH261-60N6 | 5 | 2q | |
| 18q | CH261-72B18 | |||
| 23 | CH261-105P1 | - | - | |
| 23p | CH261-191G17 | - | - | |
| 23 | CH261-49G9 | - | - | |
| 23q | CH261-90K11 | - | - | |
| 23 | TGMCBA-173N15 | - | - | |
| 23 | TGMCBA-227A15 | - | - | |
| 23 | TGMCBA-272G9 | - | 1 | 3 |
| 23 | TGMCBA-48O8 | - | - | |
| 27 | CH261-100E5 | - | - | |
| 27q | CH261-28L10 | - | 1 | 3 |
| 27p | CH261-66M16 | - | - | |
| 27 | TGMCBA-23C5 | - | - | |
| 27 | TGMCBA-324P4 | - | - | |
| 28 | CH261-101C8 | - | - | |
| 28 | CH261-186C5 | - | - | |
| 28p | CH261-64A15 | - | - | |
| 28q | CH261-72A10 | - | - | |
| Zp | CH261-129A16 | - | - | |
| Zq | CH261-133M4 | - | - | |
| Z | CH261-137F19 | - | - | |
| Z | TGMCBA-200J22 | - | - | |
| Z | TGMCBA-270I9 | - | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singchat, W.; Panthum, T.; Ahmad, S.F.; Baicharoen, S.; Muangmai, N.; Duengkae, P.; Griffin, D.K.; Srikulnath, K. Remnant of Unrelated Amniote Sex Chromosomal Linkage Sharing on the Same Chromosome in House Gecko Lizards, Providing a Better Understanding of the Ancestral Super-Sex Chromosome. Cells 2021, 10, 2969. https://doi.org/10.3390/cells10112969
Singchat W, Panthum T, Ahmad SF, Baicharoen S, Muangmai N, Duengkae P, Griffin DK, Srikulnath K. Remnant of Unrelated Amniote Sex Chromosomal Linkage Sharing on the Same Chromosome in House Gecko Lizards, Providing a Better Understanding of the Ancestral Super-Sex Chromosome. Cells. 2021; 10(11):2969. https://doi.org/10.3390/cells10112969
Chicago/Turabian StyleSingchat, Worapong, Thitipong Panthum, Syed Farhan Ahmad, Sudarath Baicharoen, Narongrit Muangmai, Prateep Duengkae, Darren K. Griffin, and Kornsorn Srikulnath. 2021. "Remnant of Unrelated Amniote Sex Chromosomal Linkage Sharing on the Same Chromosome in House Gecko Lizards, Providing a Better Understanding of the Ancestral Super-Sex Chromosome" Cells 10, no. 11: 2969. https://doi.org/10.3390/cells10112969
APA StyleSingchat, W., Panthum, T., Ahmad, S. F., Baicharoen, S., Muangmai, N., Duengkae, P., Griffin, D. K., & Srikulnath, K. (2021). Remnant of Unrelated Amniote Sex Chromosomal Linkage Sharing on the Same Chromosome in House Gecko Lizards, Providing a Better Understanding of the Ancestral Super-Sex Chromosome. Cells, 10(11), 2969. https://doi.org/10.3390/cells10112969