
The Inflammatory Lung Microenvironment; a Key Mediator in MSC Licensing

 and
and
Abstract
:1. Introduction
2. The Progress in the Clinical Translation of MSCs for Inflammatory Lung Disease
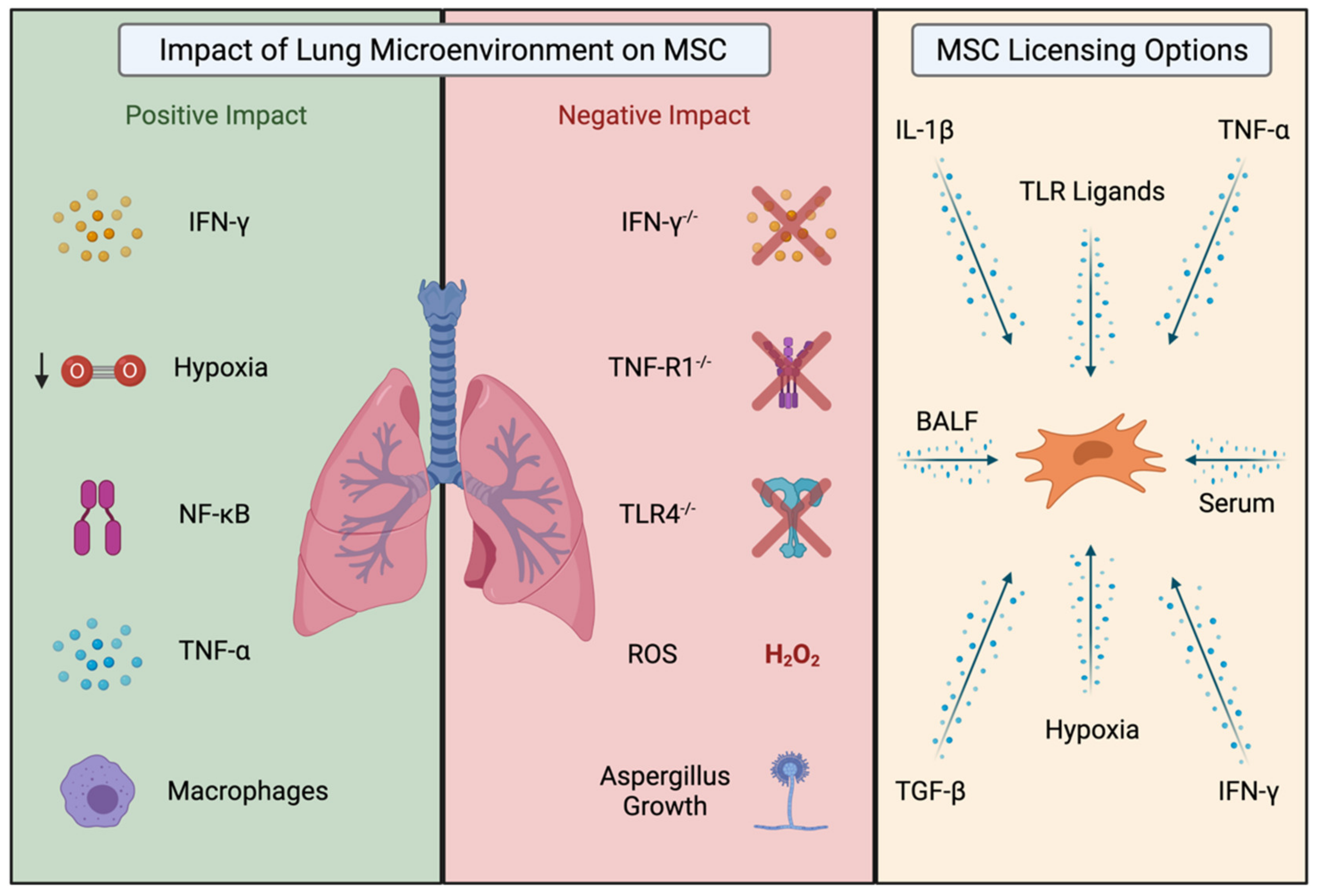
3. The Importance of the Inflammatory Disease Microenvironment on MSC Therapeutic Efficacy
4. Licensing of MSCs Enhances Their Therapeutic Efficacy
5. Exogenous Licensing of MSCs
6. Endogenous Licensing of MSCs
7. Patient Stratification to Identify Responders to MSC-Based Therapy
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cruz, F.F.; Rocco, P.R.M. The potential of mesenchymal stem cell therapy for chronic lung disease. Expert Rev. Respir. Med. 2019, 14, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Abreu, S.C.; Xisto, D.G.; De Oliveira, T.B.; Blanco, N.G.; De Castro, L.L.; Kitoko, J.; Olsen, P.; Lopes-Pacheco, M.; Morales, M.M.; Weiss, D.J.; et al. Serum from Asthmatic Mice Potentiates the Therapeutic Effects of Mesenchymal Stromal Cells in Experimental Allergic Asthma. Stem Cells Transl. Med. 2018, 8, 301–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, F.F.; Borg, Z.D.; Goodwin, M.; Sokocevic, D.; Wagner, D.E.; Coffey, A.; Antunes, M.; Robinson, K.L.; Mitsialis, S.A.; Kourembanas, S.; et al. Systemic Administration of Human Bone Marrow-Derived Mesenchymal Stromal Cell Extracellular Vesicles AmelioratesAspergillusHyphal Extract-Induced Allergic Airway Inflammation in Immunocompetent Mice. Stem Cells Transl. Med. 2015, 4, 1302–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cahill, E.F.; Kennelly, H.; Carty, F.; Mahon, B.P.; English, K. Hepatocyte Growth Factor Is Required for Mesenchymal Stromal Cell Protection Against Bleomycin-Induced Pulmonary Fibrosis. Stem Cells Transl. Med. 2016, 5, 1307–1318. [Google Scholar] [CrossRef] [PubMed]
- Cargnoni, A.; Romele, P.; Signoroni, P.B.; Farigu, S.; Magatti, M.; Vertua, E.; Toschi, I.; Cesari, V.; Silini, A.R.; Stefani, F.R.; et al. Amniotic MSCs reduce pulmonary fibrosis by hampering lung B-cell recruitment, retention, and maturation. Stem Cells Transl. Med. 2020, 9, 1023–1035. [Google Scholar] [CrossRef] [PubMed]
- Moroncini, G.; Paolini, C.; Orlando, F.; Capelli, C.; Grieco, A.; Tonnini, C.; Agarbati, S.; Mondini, E.; Saccomanno, S.; Goteri, G.; et al. Mesenchymal stromal cells from human umbilical cord prevent the development of lung fibrosis in immunocompetent mice. PLoS ONE 2018, 13, e0196048. [Google Scholar] [CrossRef]
- Antunes, M.A.; Abreu, S.C.; Cruz, F.F.; Teixeira, A.C.; Lopes-Pacheco, M.; Bandeira, E.; Olsen, P.; Diaz, B.L.; Takyia, C.M.; Freitas, I.P.R.G.; et al. Effects of different mesenchymal stromal cell sources and delivery routes in experimental emphysema. Respir. Res. 2014, 15, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennelly, H.; Mahon, B.; English, K. Human mesenchymal stromal cells exert HGF dependent cytoprotective effects in a human relevant pre-clinical model of COPD. Sci. Rep. 2016, 6, 38207. [Google Scholar] [CrossRef]
- Devaney, J.; Horie, S.; Masterson, C.; Elliman, S.; Barry, F.; O’Brien, T.; Curley, G.; O’Toole, D.; Laffey, J. Human mesenchymal stromal cells decrease the severity of acute lung injury induced by E. coli in the rat. Thorax 2015, 70, 625–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, N.; Sinha, R.; Krasnodembskaya, A.; Xu, X.; Nizet, V.; Matthay, M.A.; Griffin, J.H. The TLR4-PAR1 Axis Regulates Bone Marrow Mesenchymal Stromal Cell Survival and Therapeutic Capacity in Experimental Bacterial Pneumonia. Stem Cells 2018, 36, 796–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, M.V.; Morrison, T.J.; Doherty, D.F.; McAuley, D.; Matthay, M.A.; Kissenpfennig, A.; O’Kane, C.; Krasnodembskaya, A.D. Mitochondrial Transfer via Tunneling Nanotubes is an Important Mechanism by Which Mesenchymal Stem Cells Enhance Macrophage Phagocytosis in the In Vitro and In Vivo Models of ARDS. Stem Cells 2016, 34, 2210–2223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curley, G.; Jerkic, M.; Dixon, S.; Hogan, G.; Masterson, C.; O’Toole, D.; Devaney, J.; Laffey, J. Cryopreserved, Xeno-Free Human Umbilical Cord Mesenchymal Stromal Cells Reduce Lung Injury Severity and Bacterial Burden in Rodent Escherichia coli–Induced Acute Respiratory Distress Syndrome. Crit. Care Med. 2017, 45, e202–e212. [Google Scholar] [CrossRef] [PubMed]
- Horie, S.; Gonzalez, H.E.; Laffey, J.G.; Masterson, C.H. Cell therapy in acute respiratory distress syndrome. J. Thorac. Dis. 2018, 10, 5607–5620. [Google Scholar] [CrossRef] [PubMed]
- Masterson, C.; Devaney, J.; Horie, S.; O’Flynn, L.; Deedigan, L.; Elliman, S.; Barry, F.; O’Brien, T.; O’Toole, D.; Laffey, J.G. Syndecan-2–positive, Bone Marrow–derived Human Mesenchymal Stromal Cells Attenuate Bacterial-induced Acute Lung Injury and Enhance Resolution of Ventilator-induced Lung Injury in Rats. Anesthesiology 2018, 129, 502–516. [Google Scholar] [CrossRef] [PubMed]
- Glassberg, M.K.; Minkiewicz, J.; Toonkel, R.L.; Simonet, E.S.; Rubio, G.A.; DiFede, D.; Shafazand, S.; Khan, A.; Pujol, M.V.; LaRussa, V.F.; et al. Allogeneic Human Mesenchymal Stem Cells in Patients With Idiopathic Pulmonary Fibrosis via Intravenous Delivery (AETHER). Chest 2017, 151, 971–981. [Google Scholar] [CrossRef] [PubMed]
- Armitage, J.D.; Tan, D.B.; Sturm, M.; Moodley, Y.P. Transcriptional profiling of circulating mononuclear cells from patients with chronic obstructive pulmonary disease receiving mesenchymal stromal cell infusions. Stem Cells Transl. Med. 2021, 10, 1470–1481. [Google Scholar] [CrossRef]
- Weiss, D.J.; Casaburi, R.; Flannery, R.; LeRoux-Williams, M.; Tashkin, D.P. A Placebo-Controlled, Randomized Trial of Mesenchymal Stem Cells in COPD. Chest 2013, 143, 1590–1598. [Google Scholar] [CrossRef] [Green Version]
- Weiss, D.J.; Segal, K.; Casaburi, R.; Hayes, J.; Tashkin, D. Effect of mesenchymal stromal cell infusions on lung function in COPD patients with high CRP levels. Respir. Res. 2021, 22, 1–11. [Google Scholar] [CrossRef]
- Matthay, M.A.; Calfee, C.S.; Zhuo, H.; Thompson, B.T.; Wilson, J.G.; Levitt, J.E.; Rogers, A.J.; Gotts, J.E.; Wiener-Kronish, J.P.; Bajwa, E.K.; et al. Treatment with allogeneic mesenchymal stromal cells for moderate to severe acute respiratory distress syndrome (START study): A randomised phase 2a safety trial. Lancet Respir. Med. 2018, 7, 154–162. [Google Scholar] [CrossRef]
- Wilson, J.G.; Liu, K.D.; Zhuo, H.; Caballero, L.; McMillan, M.; Fang, X.; Cosgrove, K.; Vojnik, R.; Calfee, C.S.; Lee, J.-W.; et al. Mesenchymal stem (stromal) cells for treatment of ARDS: A phase 1 clinical trial. Lancet Respir. Med. 2014, 3, 24–32. [Google Scholar] [CrossRef] [Green Version]
- Lanzoni, G.; Linetsky, E.; Correa, D.; Cayetano, S.M.; Alvarez, R.A.; Kouroupis, D.; Gil, A.A.; Poggioli, R.; Ruiz, P.; Marttos, A.C.; et al. Umbilical cord mesenchymal stem cells for COVID-19 acute respiratory distress syndrome: A double-blind, phase 1/2a, randomized controlled trial. Stem Cells Transl. Med. 2021, 10, 660–673. [Google Scholar] [CrossRef] [PubMed]
- Galipeau, J. Mesenchymal Stromal Cells for Graft-versus-Host Disease: A Trilogy. Biol. Blood Marrow Transplant. 2020, 26, e89–e91. [Google Scholar] [CrossRef] [PubMed]
- Galipeau, J.; Sensébé, L. Mesenchymal Stromal Cells: Clinical Challenges and Therapeutic Opportunities. Cell Stem Cell 2018, 22, 824–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pittenger, M.F.; Discher, D.E.; Péault, B.M.; Phinney, D.G.; Hare, J.M.; Caplan, A.I. Mesenchymal stem cell perspective: Cell biology to clinical progress. NPJ Regen. Med. 2019, 4, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Z.W.; Baumgart, D.C. Darvadstrocel for the treatment of perianal fistulas in Crohn’s disease. Expert Rev. Gastroenterol. Hepatol. 2020, 14, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Panés, J.; García-Olmo, D.; Van Assche, G.; Colombel, J.F.; Reinisch, W.; Baumgart, D.C.; Dignass, A.; Nachury, M.; Ferrante, M.; Kazemi-Shirazi, L.; et al. Expanded allogeneic adipose-derived mesenchymal stem cells (Cx601) for complex perianal fistulas in Crohn’s disease: A phase 3 randomised, double-blind controlled trial. Lancet 2016, 388, 1281–1290. [Google Scholar] [CrossRef]
- Enes, S.R.; Krasnodembskaya, A.D.; English, K.; Dos Santos, C.C.; Weiss, D.J. Research Progress on Strategies that can Enhance the Therapeutic Benefits of Mesenchymal Stromal Cells in Respiratory Diseases With a Specific Focus on Acute Respiratory Distress Syndrome and Other Inflammatory Lung Diseases. Front. Pharmacol. 2021, 12, 811. [Google Scholar] [CrossRef]
- English, K. Mechanisms of mesenchymal stromal cell immunomodulation. Immunol. Cell Biol. 2012, 91, 19–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, D.J.; English, K.; Krasnodembskaya, A.; Isaza-Correa, J.M.; Hawthorne, I.J.; Mahon, B.P. The Necrobiology of Mesenchymal Stromal Cells Affects Therapeutic Efficacy. Front. Immunol. 2019, 10, 1228. [Google Scholar] [CrossRef]
- English, K.; French, A.; Wood, K.J. Mesenchymal Stromal Cells: Facilitators of Successful Transplantation? Cell Stem Cell 2010, 7, 431–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, P.; Calfee, C.S. Phenotypes in acute respiratory distress syndrome. Curr. Opin. Crit. Care 2019, 25, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Desai, D.; Brightling, C. Cytokine and anti-cytokine therapy in asthma: Ready for the clinic? Clin. Exp. Immunol. 2009, 158, 10–19. [Google Scholar] [CrossRef]
- Wick, K.D.; Leligdowicz, A.; Zhuo, H.; Ware, L.B.; Matthay, M.A. Mesenchymal stromal cells reduce evidence of lung injury in patients with ARDS. JCI Insight 2021, 6. [Google Scholar] [CrossRef]
- Yip, H.-K.; Fang, W.-F.; Li, Y.-C.; Lee, F.-Y.; Lee, C.-H.; Pei, S.-N.; Ma, M.-C.; Chen, K.-H.; Sung, P.-H.; Lee, M.S. Human Umbilical Cord-Derived Mesenchymal Stem Cells for Acute Respiratory Distress Syndrome. Crit. Care Med. 2020, 48, e391–e399. [Google Scholar] [CrossRef]
- Bellingan, G.; Jacono, F.; Bannard-Smith, J.; Brealey, D.; Meyer, N.; Thickett, D.; Young, D.; Bentley, A.; McVerry, B.; Wunderink, R.; et al. Primary Analysis of a Phase 1/2 Study to Assess MultiStem® Cell Therapy, a Regenerative Advanced Therapy Medicinal Product (ATMP), in Acute Respiratory Distress Syndrome (MUST-ARDS). Am. J. Respir. Crit. Care Med. 2019, 199, A7353. [Google Scholar] [CrossRef]
- Dilogo, I.H.; Aditianingsih, D.; Sugiarto, A.; Burhan, E.; Damayanti, T.; Sitompul, P.A.; Mariana, N.; Antarianto, R.D.; Liem, I.K.; Kispa, T.; et al. Umbilical cord mesenchymal stromal cells as critical COVID-19 adjuvant therapy: A randomized controlled trial. Stem Cells Transl. Med. 2021, 10, 1279–1287. [Google Scholar] [CrossRef] [PubMed]
- Ilic, D.; Liovic, M. Industry updates from the field of stem cell research and regenerative medicine in November 2020. Regen. Med. 2021, 16, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Matthay, M.A.; Arabi, Y.M.; Siegel, E.R.; Ware, L.B.; Bos, L.D.J.; Sinha, P.; Beitler, J.R.; Wick, K.D.; Curley, M.A.Q.; Constantin, J.-M.; et al. Phenotypes and personalized medicine in the acute respiratory distress syndrome. Intensiv. Care Med. 2020, 46, 2136–2152. [Google Scholar] [CrossRef]
- Vasquez, C.R.; Gupta, S.; Miano, T.A.; Roche, M.; Hsu, J.; Yang, W.; Holena, D.N.; Reilly, J.P.; Schrauben, S.J.; Leaf, D.E.; et al. Identification of Distinct Clinical Subphenotypes in Critically Ill Patients With COVID-19. Chest 2021, 160, 929–943. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.; Calfee, C.S.; McAuley, D.F. Acute Respiratory Distress Syndrome Subphenotypes beyond the Syndrome: A Step toward Treatable Traits? Am. J. Respir. Crit. Care Med. 2021, 203, 1449–1451. [Google Scholar] [CrossRef]
- Polchert, D.; Sobinsky, J.; Douglas, G.W.; Kidd, M.; Moadsiri, A.; Reina, E.; Genrich, K.; Mehrotra, S.; Setty, S.; Smith, B.J.; et al. IFN-γ activation of mesenchymal stem cells for treatment and prevention of graft versus host disease. Eur. J. Immunol. 2008, 38, 1745–1755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudres, M.; Norol, F.; Trenado, A.; Grégoire, S.; Charlotte, F.; Levacher, B.; Lataillade, J.-J.; Bourin, P.; Holy, X.; Vernant, J.-P.; et al. Bone Marrow Mesenchymal Stem Cells Suppress Lymphocyte Proliferation In Vitro but Fail to Prevent Graft-versus-Host Disease in Mice. J. Immunol. 2006, 176, 7761–7767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tobin, L.M.; Healy, M.E.; English, K.; Mahon, B.P. Human mesenchymal stem cells suppress donor CD4+T cell proliferation and reduce pathology in a humanized mouse model of acute graft-versus-host disease. Clin. Exp. Immunol. 2012, 172, 333–348. [Google Scholar] [CrossRef] [PubMed]
- Ankrum, J.; Ong, J.F.; Karp, J.M. Mesenchymal stem cells: Immune evasive, not immune privileged. Nat. Biotechnol. 2014, 32, 252–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorronsoro, A.; Ferrin, I.; Salcedo, J.M.; Jakobsson, E.; Fernández-Rueda, J.; Lang, V.; Sepulveda, P.; Fechter, K.; Pennington, D.; Trigueros, C. Human mesenchymal stromal cells modulate T-cell responses through TNF-α-mediated activation of NF-κB. Eur. J. Immunol. 2013, 44, 480–488. [Google Scholar] [CrossRef]
- Zhang, H.; Li, Y.; Slutsky, A.S. Precision medicine for cell therapy in acute respiratory distress syndrome. Lancet Respir. Med. 2019, 7, e13. [Google Scholar] [CrossRef] [Green Version]
- Islam, D.; Huang, Y.; Fanelli, V.; Delsedime, L.; Wu, S.; Khang, J.; Han, B.; Grassi, A.; Li, M.; Xu, Y.; et al. Identification and Modulation of Microenvironment Is Crucial for Effective Mesenchymal Stromal Cell Therapy in Acute Lung Injury. Am. J. Respir. Crit. Care Med. 2019, 199, 1214–1224. [Google Scholar] [CrossRef] [PubMed]
- Luz-Crawford, P.; Ipseiz, N.; Espinosa-Carrasco, G.; Caicedo, A.; Tejedor, G.; Toupet, K.; Loriau, J.; Scholtysek, C.; Stoll, C.; Khoury, M.; et al. PPARβ/δ directs the therapeutic potential of mesenchymal stem cells in arthritis. Ann. Rheum. Dis. 2016, 75, 2166–2174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carty, F.; Dunbar, H.; Hawthorne, I.J.; Ting, A.E.; Stubblefield, S.R.; Hof, W.V.; English, K. IFN-γ and PPARδ influence the efficacy and retention of multipotent adult progenitor cells in graft vs host disease. Stem Cells Transl. Med. 2021, 10, 1561–1574. [Google Scholar] [CrossRef]
- Haskova, Z.; Hoang, B.; Luo, G.; Morgan, L.A.; Billin, A.; Barone, F.C.; Shearer, B.G.; Barton, M.E.; Kilgore, K.S. Modulation of LPS-induced pulmonary neutrophil infiltration and cytokine production by the selective PPARβ/δ ligand GW0742. Inflamm. Res. 2008, 57, 314–321. [Google Scholar] [CrossRef]
- Kapoor, A.; Shintani, Y.; Collino, M.; Osuchowski, M.F.; Busch, D.; Patel, N.S.A.; Sepodes, B.; Castiglia, S.; Fantozzi, R.; Bishop-Bailey, D.; et al. Protective Role of Peroxisome Proliferator–activated Receptor-β/δ in Septic Shock. Am. J. Respir. Crit. Care Med. 2010, 182, 1506–1515. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Li, S.; Zhao, Y.; Qian, P.; Ji, F.; Qian, L.; Wu, X.; Qian, G. Silencing Angiopoietin-Like Protein 4 (ANGPTL4) Protects Against Lipopolysaccharide-Induced Acute Lung Injury Via Regulating SIRT1 /NF-kB Pathway. J. Cell. Physiol. 2015, 230, 2390–2402. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Lan, Y.; Wang, H.; Zeng, N.; Wen, F. Angiopoietin like 4 as a clinical biomarker for acute respiratory distress syndrome. Eur. Respir. J. 2019, 54. [Google Scholar] [CrossRef]
- Corbett, J.M.; Hawthorne, I.; Dunbar, H.; Coulter, I.; Ni Chonghaile, M.; Flynn, C.M.; English, K. Cyclosporine A and IFNγ licencing enhances human mesenchymal stromal cell potency in a humanised mouse model of acute graft versus host disease. Stem Cell Res. Ther. 2021, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- English, K.; Barry, F.P.; Field-Corbett, C.P.; Mahon, B. IFN-γ and TNF-α differentially regulate immunomodulation by murine mesenchymal stem cells. Immunol. Lett. 2007, 110, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Krampera, M.; Cosmi, L.; Angeli, R.; Pasini, A.; Liotta, F.; Andreini, A.; Santarlasci, V.; Mazzinghi, B.; Pizzolo, G.; Vinante, F.; et al. Role for Interferon-γ in the Immunomodulatory Activity of Human Bone Marrow Mesenchymal Stem Cells. Stem Cells 2006, 24, 386–398. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Su, J.; Zhang, L.; Zhao, X.; Ling, W.; L’Huillie, A.; Zhang, J.; Lu, Y.; Roberts, A.I.; Ji, W.; et al. Species Variation in the Mechanisms of Mesenchymal Stem Cell-Mediated Immunosuppression. Stem Cells 2009, 27, 1954–1962. [Google Scholar] [CrossRef] [PubMed]
- Duijvestein, M.; Wildenberg, M.E.; Welling, M.M.; Hennink, S.; Molendijk, I.; van Zuylen, V.L.; Bosse, T.; Vos, A.C.W.; de Jonge-Muller, E.S.M.; Roelofs, H.; et al. Pretreatment with Interferon-γ Enhances the Therapeutic Activity of Mesenchymal Stromal Cells in Animal Models of Colitis. Stem Cells 2011, 29, 1549–1558. [Google Scholar] [CrossRef]
- Rabani, R.; Volchuk, A.; Jerkic, M.; Ormesher, L.; Garces-Ramirez, L.; Canton, J.; Masterson, C.; Gagnon, S.; Tatham, K.C.; Marshall, J.; et al. Mesenchymal stem cells enhance NOX2-dependent reactive oxygen species production and bacterial killing in macrophages during sepsis. Eur. Respir. J. 2018, 51, 1702021. [Google Scholar] [CrossRef]
- Boyt, D.T.; Boland, L.K.; Burand, A.J.; Brown, A.J.; Ankrum, J.A. Dose and duration of interferon γ pre-licensing interact with donor characteristics to influence the expression and function of indoleamine-2,3-dioxygenase in mesenchymal stromal cells. J. R. Soc. Interface. 2020, 17, 20190815. [Google Scholar] [CrossRef]
- Wang, G.; Cao, K.; Liu, K.; Xue, Y.; Roberts, A.I.; Li, F.; Han, Y.; Rabson, A.B.; Wang, Y.; Shi, Y. Kynurenic acid, an IDO metabolite, controls TSG-6-mediated immunosuppression of human mesenchymal stem cells. Cell Death Differ. 2017, 25, 1209–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, A.S.; Sousa, M.R.R.; Alencar-Silva, T.; Carvalho, J.L.; Saldanha-Araujo, F. Mesenchymal stem cells immunomodulation: The road to IFN-γ licensing and the path ahead. Cytokine Growth Factor Rev. 2019, 47, 32–42. [Google Scholar] [CrossRef]
- De Witte, S.F.H.; Franquesa, M.; Baan, C.; Hoogduijn, M.J. Toward Development of iMesenchymal Stem Cells for Immunomodulatory Therapy. Front. Immunol. 2016, 6, 648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, R.H.; Pulin, A.A.; Seo, M.J.; Kota, D.J.; Ylostalo, J.; Larson, B.L.; Semprun-Prieto, L.; Delafontaine, P.; Prockop, D.J. Intravenous hMSCs Improve Myocardial Infarction in Mice because Cells Embolized in Lung Are Activated to Secrete the Anti-inflammatory Protein TSG-6. Cell Stem Cell 2009, 5, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Prasanna, S.J.; Gopalakrishnan, D.; Shankar, S.R.; Vasandan, A.B. Pro-Inflammatory Cytokines, IFNγ and TNFα, Influence Immune Properties of Human Bone Marrow and Wharton Jelly Mesenchymal Stem Cells Differentially. PLoS ONE 2010, 5, e9016. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Zheng, D.; Xu, R.-H. Critical Role of Tumor Necrosis Factor Signaling in Mesenchymal Stem Cell-Based Therapy for Autoimmune and Inflammatory Diseases. Front. Immunol. 2018, 9, 1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, Y.-W.; Choo, K.-B.; Chen, C.-M.; Chung-Hsing, H.; Chen, Y.-B.; Hsieh, C.-H.; Kuo, H.-P.; Chong, K.-Y. Hypoxia-preconditioned mesenchymal stem cells attenuate bleomycin-induced pulmonary fibrosis. Stem Cell Res. Ther. 2015, 6, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.-Y.; Chiang, C.-H.; Hung, S.-C.; Chian, C.-F.; Tsai, C.-L.; Chen, W.-C.; Zhang, H. Hypoxia-preconditioned mesenchymal stem cells ameliorate ischemia/reperfusion-induced lung injury. PLoS ONE 2017, 12, e0187637. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Liu, Q.; Qi, L.; Dai, X.; Liu, H.; Wang, Y. Low levels of TGF-β1 enhance human umbilical cord-derived mesenchymal stem cell fibronectin production and extend survival time in a rat model of lipopolysaccharide-induced acute lung injury. Mol. Med. Rep. 2016, 14, 1681–1692. [Google Scholar] [CrossRef]
- Varkouhi, A.K.; Jerkic, M.; Ormesher, L.; Gagnon, S.; Goyal, S.; Rabani, R.; Masterson, C.; Spring, C.; Chen, P.Z.; Gu, F.X.; et al. Extracellular Vesicles from Interferon-γ–primed Human Umbilical Cord Mesenchymal Stromal Cells Reduce Escherichia coli-induced Acute Lung Injury in Rats. Anesthesiology 2019, 130, 778–790. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, P.N.; Falcones, B.; Farre, R.; Artigas, A.; Almendros, I.; Navajas, D. Biophysically Preconditioning Mesenchymal Stem Cells Improves Treatment of Ventilator-Induced Lung Injury. Arch. Bronconeumol. 2019, 56, 179–181. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Yuan, X.; Zhang, J.; Lu, T.; Yao, J.; Zheng, J.; Cai, J.; Xiao, J.; Chen, H.; Xie, S.; et al. Heat shock preconditioning mesenchymal stem cells attenuate acute lung injury via reducing NLRP3 inflammasome activation in macrophages. Stem Cell Res. Ther. 2021, 12, 1–16. [Google Scholar] [CrossRef]
- Bustos, M.L.; Huleihel, L.; Meyer, E.M.; Donnenberg, A.D.; Donnenberg, V.S.; Sciurba, J.D.; Mroz, L.; McVerry, B.J.; Ellis, B.M.; Kaminski, N.; et al. Activation of Human Mesenchymal Stem Cells Impacts Their Therapeutic Abilities in Lung Injury by Increasing Interleukin (IL)-10 and IL-1RN Levels. Stem Cells Transl. Med. 2013, 2, 884–895. [Google Scholar] [CrossRef]
- Morrison, T.J.; Jackson, M.V.; Cunningham, E.K.; Kissenpfennig, A.; McAuley, D.; O’Kane, C.; Krasnodembskaya, A.D. Mesenchymal Stromal Cells Modulate Macrophages in Clinically Relevant Lung Injury Models by Extracellular Vesicle Mitochondrial Transfer. Am. J. Respir. Crit. Care Med. 2017, 196, 1275–1286. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.D.; De Castro, L.L.; Braga, C.L.; Oliveira, G.P.; Trivelin, S.A.; Barbosa-Junior, C.M.; Morales, M.M.; Dos Santos, C.C.; Weiss, D.J.; Lopes-Pacheco, M.; et al. Mesenchymal Stromal Cells Are More Effective Than Their Extracellular Vesicles at Reducing Lung Injury Regardless of Acute Respiratory Distress Syndrome Etiology. Stem Cells Int. 2019, 2019, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galleu, A.; Riffo-Vasquez, Y.; Trento, C.; Lomas, C.; Dolcetti, L.; Cheung, T.S.; von Bonin, M.; Barbieri, L.; Halai, K.; Ward, S.; et al. Apoptosis in mesenchymal stromal cells induces in vivo recipient-mediated immunomodulation. Sci. Transl. Med. 2017, 9, 158–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, K.; Treacy, O.; Chen, X.; Murphy, N.; Lohan, P.; Islam, N.; Donohoe, E.; Griffin, M.D.; Watson, L.; McLoughlin, S.; et al. TGF-β1-Licensed Murine MSCs Show Superior Therapeutic Efficacy in Modulating Corneal Allograft Immune Rejection In Vivo. Mol. Ther. 2020, 28, 2023–2043. [Google Scholar] [CrossRef] [PubMed]
- Murphy, N.; Treacy, O.; Lynch, K.; Morcos, M.; Lohan, P.; Howard, L.; Fahy, G.; Griffin, M.D.; Ryan, A.E.; Ritter, T. TNF-α/IL-1β—Licensed mesenchymal stromal cells promote corneal allograft survival via myeloid cell-mediated induction of Foxp3 + regulatory T cells in the lung. FASEB J. 2019, 33, 9404–9421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Németh, K.; Leelahavanichkul, A.; Yuen, P.; Mayer, B.; Parmelee, A.; Doi, K.; Robey, P.; Leelahavanichkul, K.; Koller, B.H.; Brown, J.M.; et al. Bone marrow stromal cells attenuate sepsis via prostaglandin E2–dependent reprogramming of host macrophages to increase their interleukin-10 production. Nat. Med. 2008, 15, 42–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abreu, S.C.; Weiss, D.J.; Rocco, P.R.M. Extracellular vesicles derived from mesenchymal stromal cells: A therapeutic option in respiratory diseases? Stem Cell Res. Ther. 2016, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.J.W.; Matthay, M.M.A. Is a Part Better than the Whole for Cell-based Therapy for Acute Respiratory Distress Syndrome? Anesthesiology 2019, 130, 683–685. [Google Scholar] [CrossRef] [Green Version]
- Phinney, D.; Di Giuseppe, M.; Njah, J.; Sala-Llinas, E.; Shiva, S.; Croix, C.M.S.; Stolz, D.B.; Watkins, S.; Di, Y.P.; Leikauf, G.; et al. Mesenchymal stem cells use extracellular vesicles to outsource mitophagy and shuttle microRNAs. Nat. Commun. 2015, 6, 8472. [Google Scholar] [CrossRef] [PubMed]
- Harting, M.T.; Srivastava, A.; Zhaorigetu, S.; Bair, H.; Prabhakara, K.S.; Furman, N.E.T.; Vykoukal, J.V.; Ruppert, K.A.; Cox, C.S.; Olson, S.D. Inflammation-Stimulated Mesenchymal Stromal Cell-Derived Extracellular Vesicles Attenuate Inflammation. Stem Cells 2017, 36, 79–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Fu, L.; Liang, Y.; Guo, Z.; Wang, L.; Ma, C.; Wang, H. Exosomes originating from MSCs stimulated with TGF-β and IFN-γ promote Treg differentiation. J. Cell. Physiol. 2018, 233, 6832–6840. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.; Baveja, R.; Liang, O.D.; Fernandez-Gonzalez, A.; Lee, C.; Mitsialis, S.A.; Kourembanas, S. Bone Marrow Stromal Cells Attenuate Lung Injury in a Murine Model of Neonatal Chronic Lung Disease. Am. J. Respir. Crit. Care Med. 2009, 180, 1122–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, Q.; Gudapati, V.; Monsel, A.; Park, J.H.; Hu, S.; Kato, H.; Lee, J.H.; Zhou, L.; He, H. Mesenchymal Stem Cell–Derived Extracellular Vesicles Decrease Lung Injury in Mice. J. Immunol. 2019, 203, 1961–1972. [Google Scholar] [CrossRef] [PubMed]
- Monsel, A.; Zhu, Y.-G.; Gennai, S.; Hao, Q.; Hu, S.; Rouby, J.-J.; Rosenzwajg, M.; Matthay, M.A.; Lee, J.W. Therapeutic Effects of Human Mesenchymal Stem Cell–derived Microvesicles in Severe Pneumonia in Mice. Am. J. Respir. Crit. Care Med. 2015, 192, 324–336. [Google Scholar] [CrossRef] [Green Version]
- De Castro, L.L.; Xisto, D.G.; Kitoko, J.Z.; Cruz, F.F.; Olsen, P.C.; Redondo, P.A.G.; Ferreira, T.P.T.; Weiss, D.J.; Martins, M.A.; Morales, M.M.; et al. Human adipose tissue mesenchymal stromal cells and their extracellular vesicles act differentially on lung mechanics and inflammation in experimental allergic asthma. Stem Cell Res. Ther. 2017, 8, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, J.D.; Su, Y.; Calfee, C.S.; Delucchi, K.L.; Weiss, D.; McAuley, D.F.; O’Kane, C.; Krasnodembskaya, A.D. Mesenchymal stromal cell extracellular vesicles rescue mitochondrial dysfunction and improve barrier integrity in clinically relevant models of ARDS. Eur. Respir. J. 2020, 58, 2002978. [Google Scholar] [CrossRef] [PubMed]
- Lan, Y.-W.; Theng, S.-M.; Huang, T.-T.; Choo, K.-B.; Chen, C.-M.; Kuo, H.-P.; Chong, K.Y. Oncostatin M-Preconditioned Mesenchymal Stem Cells Alleviate Bleomycin-Induced Pulmonary Fibrosis Through Paracrine Effects of the Hepatocyte Growth Factor. Stem Cells Transl. Med. 2016, 6, 1006–1017. [Google Scholar] [CrossRef]
- Hackel, A.; Aksamit, A.; Bruderek, K.; Lang, S.; Brandau, S. TNF-α and IL-1β sensitize human MSC for IFN-γ signaling and enhance neutrophil recruitment. Eur. J. Immunol. 2020, 51, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Reading, J.L.; Vaes, B.; Hull, C.; Sabbah, S.; Hayday, T.; Wang, N.S.; DiPiero, A.; A Lehman, N.; Taggart, J.M.; Carty, F.; et al. Suppression of IL-7-dependent Effector T-cell Expansion by Multipotent Adult Progenitor Cells and PGE2. Mol. Ther. 2015, 23, 1783–1793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterman, R.S.; Tomchuck, S.L.; Henkle, S.L.; Betancourt, A.M. A New Mesenchymal Stem Cell (MSC) Paradigm: Polarization into a Pro-Inflammatory MSC1 or an Immunosuppressive MSC2 Phenotype. PLoS ONE 2010, 5, e10088. [Google Scholar] [CrossRef]
- Qiu, Y.; Guo, J.; Mao, R.; Chao, K.; Chen, B.-L.; He, Y.; Zeng, Z.-R.; Zhang, S.-H.; Chen, M.-H. TLR3 preconditioning enhances the therapeutic efficacy of umbilical cord mesenchymal stem cells in TNBS-induced colitis via the TLR3-Jagged-1-Notch-1 pathway. Mucosal Immunol. 2016, 10, 727–742. [Google Scholar] [CrossRef] [PubMed]
- Mastri, M.; Shah, Z.; McLaughlin, T.; Greene, C.J.; Baum, L.; Suzuki, G.; Lee, T. Activation of Toll-like receptor 3 amplifies mesenchymal stem cell trophic factors and enhances therapeutic potency. Am. J. Physiol. Physiol. 2012, 303, C1021–C1033. [Google Scholar] [CrossRef] [PubMed]
- Park, A.; Park, H.; Yoon, J.; Kang, D.; Kang, M.-H.; Park, Y.-Y.; Suh, N.; Yu, J. Priming with Toll-like receptor 3 agonist or interferon-gamma enhances the therapeutic effects of human mesenchymal stem cells in a murine model of atopic dermatitis. Stem Cell Res. Ther. 2019, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Kim, S.; Lim, H.; Liu, A.; Hu, S.; Lee, J.; Zhuo, H.; Hao, Q.; Matthay, M.A.; Lee, J.-W. Therapeutic effects of human mesenchymal stem cell microvesicles in an ex vivo perfused human lung injured with severe E. coli pneumonia. Thorax 2018, 74, 43–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurte, M.; Vega-Letter, A.M.; Luz-Crawford, P.; Djouad, F.; Noël, D.; Khoury, M.; Carrión, F. Time-dependent LPS exposure commands MSC immunoplasticity through TLR4 activation leading to opposite therapeutic outcome in EAE. Stem Cell Res. Ther. 2020, 11, 1–14. [Google Scholar] [CrossRef]
- Ti, D.; Hao, H.; Tong, C.; Liu, J.; Dong, L.; Zheng, J.; Zhao, Y.; Liu, H.; Fu, X.; Han, W. LPS-preconditioned mesenchymal stromal cells modify macrophage polarization for resolution of chronic inflammation via exosome-shuttled let-7b. J. Transl. Med. 2015, 13, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.-C.; Chiang, B.-L. Toll-like receptor 2 ligation of mesenchymal stem cells alleviates asthmatic airway inflammation. J. Allergy Clin. Immunol. 2018, 142, 284–287.e5. [Google Scholar] [CrossRef] [Green Version]
- Kota, D.J.; Dicarlo, B.; Hetz, R.A.; Smith, P.; Cox, C.S.; Olson, S.D. Differential MSC activation leads to distinct mononuclear leukocyte binding mechanisms. Sci. Rep. 2014, 4, 4565. [Google Scholar] [CrossRef]
- Najar, M.; Krayem, M.; Meuleman, N.; Bron, D.; Lagneaux, L. Mesenchymal Stromal Cells and Toll-Like Receptor Priming: A Critical Review. Immune Netw. 2017, 17, 89–102. [Google Scholar] [CrossRef] [Green Version]
- Lei, J.; Wang, Z.; Hui, D.; Yu, W.; Zhou, D.; Xia, W.; Chen, C.; Zhang, Q.; Wang, Z.; Zhang, Q.; et al. Ligation of TLR2 and TLR4 on murine bone marrow-derived mesenchymal stem cells triggers differential effects on their immunosuppressive activity. Cell. Immunol. 2011, 271, 147–156. [Google Scholar] [CrossRef]
- Liu, C.; Zhang, Y.-S.; Chen, F.; Wu, X.-Y.; Zhang, B.-B.; Wu, Z.-D.; Lei, J.-X. Immunopathology in schistosomiasis is regulated by TLR2,4- and IFN-γ-activated MSC through modulating Th1/Th2 responses. Stem Cell Res. Ther. 2020, 11, 217. [Google Scholar] [CrossRef] [PubMed]
- Vega-Letter, A.M.; Kurte, M.; Fernández-O’Ryan, C.; Gauthier-Abeliuk, M.; Fuenzalida, P.; Moya-Uribe, I.; Altamirano, C.; Figueroa, F.; Irarrázabal, C.; Carrión, F. Differential TLR activation of murine mesenchymal stem cells generates distinct immunomodulatory effects in EAE. Stem Cell Res. Ther. 2016, 7, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amiri, F.; Jahanian-Najafabadi, A.; Roudkenar, M.H. In vitro augmentation of mesenchymal stem cells viability in stressful microenvironments. Cell Stress Chaperon- 2014, 20, 237–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boregowda, S.V.; Krishnappa, V.; Chambers, J.; Lograsso, P.V.; Lai, W.-T.; Ortiz, L.; Phinney, D.G. Atmospheric Oxygen Inhibits Growth and Differentiation of Marrow-Derived Mouse Mesenchymal Stem Cells via a p53-Dependent Mechanism: Implications for Long-Term Culture Expansion. Stem Cells 2012, 30, 975–987. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Yu, S.P.; Fraser, J.L.; Lu, Z.; Ogle, M.E.; Wang, J.-A.; Wei, L. Transplantation of hypoxia-preconditioned mesenchymal stem cells improves infarcted heart function via enhanced survival of implanted cells and angiogenesis. J. Thorac. Cardiovasc. Surg. 2008, 135, 799–808. [Google Scholar] [CrossRef] [Green Version]
- Levy, O.; Kuai, R.; Siren, E.M.J.; Bhere, D.; Milton, Y.; Nissar, N.; De Biasio, M.; Heinelt, M.; Reeve, B.; Abdi, R.; et al. Shattering barriers toward clinically meaningful MSC therapies. Sci. Adv. 2020, 6, eaba6884. [Google Scholar] [CrossRef] [PubMed]
- Bader, A.M.; Klose, K.; Bieback, K.; Korinth, D.; Schneider, M.; Seifert, M.; Choi, Y.-H.; Kurtz, A.; Falk, V.; Stamm, C. Hypoxic Preconditioning Increases Survival and Pro-Angiogenic Capacity of Human Cord Blood Mesenchymal Stromal Cells In Vitro. PLoS ONE 2015, 10, e0138477. [Google Scholar] [CrossRef]
- Lee, J.H.; Yoon, Y.M.; Lee, S.H. Hypoxic Preconditioning Promotes the Bioactivities of Mesenchymal Stem Cells via the HIF-1α-GRP78-Akt Axis. Int. J. Mol. Sci. 2017, 18, 1320. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Wei, Z.Z.; Gu, X.; Zhang, J.Y.; Zhang, Y.; Li, J.; Wei, L. Intranasal delivery of hypoxia-preconditioned bone marrow-derived mesenchymal stem cells enhanced regenerative effects after intracerebral hemorrhagic stroke in mice. Exp. Neurol. 2015, 272, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Yin, S.; Zhang, W.; Gao, F.; Liu, Y.; Chen, Z.; Zhang, M.; He, J.; Zheng, S. Hypoxia preconditioned bone marrow mesenchymal stem cells promote liver regeneration in a rat massive hepatectomy model. Stem Cell Res. Ther. 2013, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Brahimi-Horn, M.C.; Pouysségur, J. Oxygen, a source of life and stress. FEBS Lett. 2007, 581, 3582–3591. [Google Scholar] [CrossRef] [PubMed]
- Noronha, N.C.; Mizukami, A.; Caliári-Oliveira, C.; Cominal, J.G.; Rocha, J.L.M.; Covas, D.T.; Swiech, K.; Malmegrim, K.C.R. Priming approaches to improve the efficacy of mesenchymal stromal cell-based therapies. Stem Cell Res. Ther. 2019, 10, 1–21. [Google Scholar]
- Da Silva, L.H.A.; Antunes, M.A.; Dos Santos, C.C.; Weiss, D.J.; Cruz, F.F.; Rocco, P.R.M. Strategies to improve the therapeutic effects of mesenchymal stromal cells in respiratory diseases. Stem Cell Res. Ther. 2018, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Waszak, P.; Alphonse, R.; Vadivel, A.; Ionescu, L.; Eaton, F.; Thébaud, B. Preconditioning Enhances the Paracrine Effect of Mesenchymal Stem Cells in Preventing Oxygen-Induced Neonatal Lung Injury in Rats. Stem Cells Dev. 2012, 21, 2789–2797. [Google Scholar] [CrossRef]
- Block, G.J.; Ohkouchi, S.; Fung, F.; Frenkel, J.; Gregory, C.; Pochampally, R.; DiMattia, G.; Sullivan, D.E.; Prockop, D.J. Multipotent stromal cells are activated to reduce apoptosis in part by upregulation and secretion of stanniocalcin-1. Stem Cells 2009, 27, 670–681. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, X.-B.; Huang, S.; Bi, X.-Y.; Wang, H.-X.; Xie, L.-X.; Wang, Y.-Q.; Cao, X.-F.; Lv, J.; Xiao, F.-J.; et al. Microvesicles Derived from Human Umbilical Cord Mesenchymal Stem Cells Stimulated by Hypoxia Promote Angiogenesis Both In Vitro and In Vivo. Stem Cells Dev. 2012, 21, 3289–3297. [Google Scholar] [CrossRef]
- Li, L.; Jin, S.; Zhang, Y. Ischemic preconditioning potentiates the protective effect of mesenchymal stem cells on endotoxin-induced acute lung injury in mice through secretion of exosome. Int. J. Clin. Exp. Med. 2015, 8, 3825–3832. [Google Scholar] [PubMed]
- Abreu, S.C.; Enes, S.R.; Dearborn, J.; Goodwin, M.; Coffey, A.; Borg, Z.D.; Dos Santos, C.C.; Wargo, M.J.; Cruz, F.F.; Loi, R.; et al. Lung inflammatory environments differentially alter mesenchymal stromal cell behavior. Am. J. Physiol. Cell. Mol. Physiol. 2019, 317, L823–L831. [Google Scholar] [CrossRef] [PubMed]
- Xu, A.; Ii, L.A.R.; Walker, K.P.; Mohammadipoor, A.; Kamucheka, R.M.; Cancio, L.C.; Batchinsky, A.I.; Antebi, B. Mesenchymal Stem Cells Reconditioned in Their Own Serum Exhibit Augmented Therapeutic Properties in the Setting of Acute Respiratory Distress Syndrome. Stem Cells Transl. Med. 2019, 8, 1092–1106. [Google Scholar] [CrossRef] [Green Version]
- Abreu, S.C.; Hampton, T.H.; Hoffman, E.; Dearborn, J.; Ashare, A.; Singh Sidhu, K.; Matthews, D.E.; McKenna, D.H.; Amiel, E.; Barua, J.; et al. Differential effects of the cystic fibrosis lung inflammatory environment on mesenchymal stromal cells. Am. J. Physiol. Lung Cell Mol. Physiol. 2020, 319, 908–925. [Google Scholar] [CrossRef] [PubMed]
- Enes, S.R.; Hampton, T.H.; Barua, J.; McKenna, D.H.; dos Santos, C.C.; Amiel, E.; Ashare, A.; Liu, K.D.; Krasnodembskaya, A.D.; English, K.; et al. Healthy versus inflamed lung environments differentially affect mesenchymal stromal cells. Eur. Respir. J. 2021, 58, 2004149. [Google Scholar] [CrossRef]
- Goodwin, M.; Sueblinvong, V.; Eisenhauer, P.; Ziats, N.P.; LeClair, L.; Poynter, M.E.; Steele, C.; Rincon, M.; Weiss, D.J. Bone marrow-derived mesenchymal stromal cells inhibit Th2-mediated allergic airways inflammation in mice. Stem Cells 2011, 29, 1137–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danchuk, S.; Ylostalo, J.H.; Hossain, F.; Sorge, R.; Ramsey, A.; Bonvillain, R.W.; Lasky, J.A.; Bunnell, B.A.; Welsh, D.A.; Prockop, D.J.; et al. Human multipotent stromal cells attenuate lipopolysaccharide-induced acute lung injury in mice via secretion of tumor necrosis factor-α-induced protein 6. Stem Cell Res. Ther. 2011, 2, 27. [Google Scholar] [CrossRef] [Green Version]
- Jahromi, S.H.; Li, Y.; Davies, J.E.; Jahromi, M.S.H.; Li, M.Y. Effect of Tumor Necrosis Factor Alpha Dose and Exposure Time on Tumor Necrosis Factor-Induced Gene-6 Activation by Neonatal and Adult Mesenchymal Stromal Cells. Stem Cells Dev. 2018, 27, 44–54. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, Z.; Jiang, D.; Liang, X.; Liao, S.; Zhang, Z.; Yue, W.; Li, X.; Chiu, S.-M.; Chai, Y.-H.; et al. iPSC-MSCs with High Intrinsic MIRO1 and Sensitivity to TNF-α Yield Efficacious Mitochondrial Transfer to Rescue Anthracycline-Induced Cardiomyopathy. Stem Cell Rep. 2016, 7, 749–763. [Google Scholar] [CrossRef] [Green Version]
- Naftali-Shani, N.; Levin-Kotler, L.-P.; Palevski, D.; Amit, U.; Kain, D.; Landa, N.; Hochhauser, E.; Leor, J. Left Ventricular Dysfunction Switches Mesenchymal Stromal Cells Toward an Inflammatory Phenotype and Impairs Their Reparative Properties Via Toll-Like Receptor-4. Circulation 2017, 135, 2271–2287. [Google Scholar] [CrossRef] [PubMed]
- Carty, F.; Mahon, B.P.; English, K. The influence of macrophages on mesenchymal stromal cell therapy: Passive or aggressive agents? Clin. Exp. Immunol. 2017, 188, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Stevens, H.Y.; Bowles, A.C.; Yeago, C.; Roy, K. Molecular Crosstalk Between Macrophages and Mesenchymal Stromal Cells. Front. Cell Dev. Biol. 2020, 8, 1436. [Google Scholar] [CrossRef] [PubMed]
- Chiossone, L.; Conte, R.; Spaggiari, G.M.; Serra, M.; Romei, C.; Bellora, F.; Becchetti, F.; Andaloro, A.; Moretta, L.; Bottino, C. Mesenchymal Stromal Cells Induce Peculiar Alternatively Activated Macrophages Capable of Dampening Both Innate and Adaptive Immune Responses. Stem Cells 2016, 34, 1909–1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, J.H.; Lee, H.J.; Jeong, H.J.; Kim, M.K.; Wee, W.R.; Yoon, S.-O.; Choi, H.; Prockop, D.J.; Oh, J.Y. Mesenchymal stem/stromal cells precondition lung monocytes/macrophages to produce tolerance against allo- and autoimmunity in the eye. Proc. Natl. Acad. Sci. USA 2015, 113, 158–163. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhang, R.; Yan, K.; Chen, F.; Huang, W.; Lv, B.; Sun, C.; Xu, L.; Li, F.; Jiang, X. Mesenchymal stem cells inhibit lipopolysaccharide-induced inflammatory responses of BV2 microglial cells through TSG-6. J. Neuroinflammation 2014, 11, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luz-Crawford, P.; Djouad, F.; Toupet, K.; Bony, C.; Franquesa, M.; Hoogduijn, M.J.; Jorgensen, C.; Noël, D. Mesenchymal Stem Cell-Derived Interleukin 1 Receptor Antagonist Promotes Macrophage Polarization and Inhibits B Cell Differentiation. Stem Cells 2015, 34, 483–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz, L.A.; DuTreil, M.; Fattman, C.; Pandey, A.C.; Torres, G.; Go, K.; Phinney, D.G. Interleukin 1 receptor antagonist mediates the antiinflammatory and antifibrotic effect of mesenchymal stem cells during lung injury. Proc. Natl. Acad. Sci. USA 2007, 104, 11002–11007. [Google Scholar] [CrossRef] [Green Version]
- Ghanem, L.Y.; Mansour, I.M.; Abulata, N.; Akl, M.M.; Demerdash, Z.A.; El Baz, H.G.; Mahmoud, S.S.; Mohamed, S.H.; Mahmoud, F.S.; Hassan, A.S.M. Liver Macrophage Depletion Ameliorates The Effect of Mesenchymal Stem Cell Transplantation in a Murine Model of Injured Liver. Sci. Rep. 2019, 9, 35. [Google Scholar] [CrossRef] [PubMed]
- Giri, J.; Das, R.; Nylen, E.; Chinnadurai, R.; Galipeau, J. CCL2 and CXCL12 Derived from Mesenchymal Stromal Cells Cooperatively Polarize IL-10+ Tissue Macrophages to Mitigate Gut Injury. Cell Rep. 2020, 30, 1923–1934.e4. [Google Scholar] [CrossRef] [Green Version]
- Braza, F.; Dirou, S.; Forest, V.; Sauzeau, V.; Hassoun, D.; Chesné, J.; Cheminant-Muller, M.-A.; Sagan, C.; Magnan, A.; Lemarchand, P. Mesenchymal Stem Cells Induce Suppressive Macrophages Through Phagocytosis in a Mouse Model of Asthma. Stem Cells 2016, 34, 1836–1845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Witte, S.F.; Luk, F.; Sierra-Parraga, J.M.; Gargesha, M.; Merino, A.; Korevaar, S.S.; Shankar, A.S.; O’Flynn, L.; Elliman, S.J.; Roy, D.; et al. Immunomodulation By Therapeutic Mesenchymal Stromal Cells (MSC) Is Triggered Through Phagocytosis of MSC By Monocytic Cells. Stem Cells 2018, 36, 602–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, J.H.; Kim, H.J.; Jeong, H.J.; Lee, H.J.; Oh, J.Y. Mesenchymal Stem and Stromal Cells Harness Macrophage-Derived Amphiregulin to Maintain Tissue Homeostasis. Cell Rep. 2020, 30, 3806–3820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, H.; Xu, L.; Parrott, R.; Overall, C.C.; Lillich, M.; Rabjohns, E.M.; Rampersad, R.R.; Tarrant, T.K.; Meadows, N.; Fernandez-Castaneda, A.; et al. Mesenchymal stromal cells reprogram monocytes and macrophages with processing bodies. Stem Cells 2021, 39, 115–128. [Google Scholar] [PubMed]
- Weiss, A.R.R.; Lee, O.; Eggenhofer, E.; Geissler, E.; Korevaar, S.S.; Soeder, Y.; Schlitt, H.J.; Geissler, E.K.; Hoogduijn, M.J.; Dahlke, M.H.; et al. Differential effects of heat-inactivated, secretome-deficient MSC and metabolically active MSC in sepsis and allogenic heart transplantation. Stem Cells 2020, 38, 797–807. [Google Scholar] [CrossRef]
- Carty, F.; Corbett, J.M.; da Cunha, J.P.M.C.M.; Reading, J.L.; Tree, T.; Ting, A.; Stubblefield, S.R.; English, K. Multipotent Adult Progenitor Cells Suppress T Cell Activation in In Vivo Models of Homeostatic Proliferation in a Prostaglandin E2-Dependent Manner. Front. Immunol. 2018, 9, 645. [Google Scholar] [CrossRef] [PubMed]
- Armitage, J.; Tan, D.B.; Troedson, R.; Young, P.; Lam, K.-V.; Shaw, K.; Sturm, M.; Weiss, D.J.; Moodley, Y.P. Mesenchymal stromal cell infusion modulates systemic immunological responses in stable COPD patients: A phase I pilot study. Eur. Respir. J. 2018, 51, 1702369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Oliveira, H.G.; Cruz, F.; Antunes, M.A.; Neto, A.V.D.M.; Oliveira, G.A.; Svartman, F.M.; Borgonovo, T.; Rebelatto, C.L.K.; Weiss, D.J.; Brofman, P.R.S.; et al. Combined Bone Marrow-Derived Mesenchymal Stromal Cell Therapy and One-Way Endobronchial Valve Placement in Patients with Pulmonary Emphysema: A Phase I Clinical Trial. Stem Cells Transl. Med. 2016, 6, 962–969. [Google Scholar] [CrossRef] [PubMed]
- Stolk, J.; Broekman, W.; Mauad, T.; Zwaginga, J.; Roelofs, H.; Fibbe, W.; Oostendorp, J.; Bajema, I.; Versteegh, M.; Taube, C.; et al. A phase I study for intravenous autologous mesenchymal stromal cell administration to patients with severe emphysema. Qjm: Int. J. Med. 2016, 109, 331–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, D.C.; Enever, D.; Ilic, N.; Sparks, L.; Whitelaw, K.; Ayres, J.; Yerkovich, S.; Khalil, D.; Atkinson, K.M.; Hopkins, P.M. A phase 1b study of placenta-derived mesenchymal stromal cells in patients with idiopathic pulmonary fibrosis. Respirology 2014, 19, 1013–1018. [Google Scholar] [CrossRef]
- Ortiz, L.A.; Gambelli, F.; McBride, C.; Gaupp, D.; Baddoo, M.; Kaminski, N.; Phinney, D. Mesenchymal stem cell engraftment in lung is enhanced in response to bleomycin exposure and ameliorates its fibrotic effects. Proc. Natl. Acad. Sci. USA 2003, 100, 8407–8411. [Google Scholar] [CrossRef] [Green Version]
- Sabry, M.M.; Elkalawy, S.A.-E.; Abo-Elnour, R.K.E.-D.; Abd-El-Maksod, D.F. Histolgical AND Immunohistochemical Study on the Effect of Stem Cell Therapy on Bleomycin Induced Pulmonary Fibrosis in Albino Rat. Int. J. Stem Cells 2014, 7, 33–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laffey, J.G.; Matthay, M.A. FiftyYearsofResearchinARDS.Cell-based Therapy for Acute Respiratory Distress Syndrome. Biology and Potential Therapeutic Value. Am. J. Respir. Crit. Care Med. 2017, 196, 266–273. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Murine Model | MSC Source | Route of Administration | Licensing Method | Licensing Location | Outcome | Ref |
|---|---|---|---|---|---|---|
| IPF (bleomycin) | Mouse BM-MSC | I.T. | Hypoxia (1.5% O2) | in vitro | ↑ HIF1α, HGF, VEGF ↓ IL-6, pro-IL-1β | [67] |
| ALI | Rat BM-MSC | In perfusate | Hypoxia (1% O2) | in vitro | ↑ IL-10, PGE2 ↓ Lung injury score ↓ TNF-α, IL-1β, MIP-2 | [68] |
| ALI | Human UC-MSC | I.V. | TGF-β1 | in vitro | ↑ MSC survival ↑ Expression of RhoA ↓ LPS-induced injury | [69] |
| ALI | Human UC-MSC (EVs) | I.V. | IFN-γ | in vitro | ↑ Animal survival ↑ eNOS ↓ Lung injury score ↓ TNF-α | [70] |
| ALI | Rat lung MSCs | I.V. | Culture on lung ECM-cyclic stretch | in vitro | ↑ Lung elastance ↓ TNF-α, CXCL2 + neutrophils | [71] |
| ALI | Human UC-MSC | I.N. | Heatshock (42 °C for 1 h) | in vitro | ↑ HSP70 expression ↑ IL-10 + PGE2 ↓ NLRP3 inflammasome formation ↓ IL-1β secretion by macrophages | [72] |
| ALI | Human BM-MSC | I.V. | Co-culture w/ serum from ARDS patients | ex vivo | ↑ IL-10 + IL-1RN ↓ IL-6, IL-1α, IL-8, IL-1β, IFN-γ, TGFβ3, TGFβ2 + TNFAIP6 | [73] |
| ALI | Human BM-MSC | AM treated with MSC- EVs-I.V. | Co-culture w/ BALF from ARDS patients | ex vivo | ↑ M2 macrophage marker expression ↑ Phagocytic capacity of human MDMs ↓ Cytokine production | [74] |
| ALI | Human BM-MSC | I.T. | Cco-culture w/ plasma from ARDS patients | ex vivo | ↑ IL-6 production ↑ N-cadherin expression at mRNA + protein levels ↓ CD105 + CD90 marker expression at day 5 | [47] |
| ALI | Mouse BM-MSC | I.V. | Co-culture w/ serum from ALI mice | ex vivo | ↑ Expression of anti-inflammatory mediators (TGF-β + IL-10) in AM in vitro ↓ iNOS + IL-6 | [75] |
| Asthma | Mouse BM-MSC | I.T. | Co-culture w/ serum or BALF from asthmatic mice | ex vivo | ↑ TGF-β, IFN-γ, IL-10,TSG-6, IDO-1, IL-1RN, iNOS, TNF-α, IL-1β + arginase-2 ↑ caspase-3, bax ↓ bcl-2Serum:↓ IL-4, IL-13 + eotaxin | [2] |
| Murine Model | MSC Source | Route of Administration | Licensing Method | Licensing Location | Outcome | Ref |
|---|---|---|---|---|---|---|
| GvHD | Human BM-MSC | I.V. | IFN-γ | in vitro | ↑ Animal survival ↓ Cellular infiltration ↓ Pathology severity in small intestine + liver | [43] |
| GvHD | Human MAPC | I.V. | IFN-γ / PPARδ antagonist | in vitro | ↑ Efficacy on day 0 ↑ Retention in spleen + liver | [49] |
| GvHD | Human BM-MSC | I.V. | Cyclosporine A + IFN-γ | in vitro | ↑ IDO production + activity ↑ Animal survival | [54] |
| GvHD | Human BM-MSC | I.V. | N/A | N/A | ↑ Immunosuppression ↑ IDO production in recipient phagocytes | [76] |
| Corneal allograft | Mouse BM-MSC | I.V. | TGF-β1 | in vitro | ↑ CD73 expression ↑ Treg expansion ↑ Immunosuppression ↑ Allograft survival ↓ Syngeneic T cell proliferation | [77] |
| Corneal allograft | Rat BM-MSC | I.V. | IFN-γ + TNF-α + IL-1β | in vitro | ↑ Nitric oxide production ↑ Suppression of syngeneic lymphocytes ↑ Allograft survival ↑ Myeloid cells in lung ↑ FoxP3+ Treg population in lung + spleen | [78] |
| Sepsis | BM-MSC | I.V. | N/A | N/A | ↑ Animal survival ↑ Organ function ↓ TNF-α + IL-6 | [79] |
| Arthritis | Human BM-MSC | I.V. | PPARβ/δ antagonist orIFN-γ + TNF-α | in vitro | ↑ Immunosuppression ↑ NF-kB activity ↑ p65 binding on iNOS promoter ↓ Severity of disease | [48] |
| Myocardial Infarction | Human BM-MSC | I.V. | TNF-α | in vitro | ↑ TSG-6 mRNA in lung ↓ Infarct size ↓ Pro-inflammatory proteases ↓ MMP9 | [64] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dunbar, H.; Weiss, D.J.; Rolandsson Enes, S.; Laffey, J.G.; English, K. The Inflammatory Lung Microenvironment; a Key Mediator in MSC Licensing. Cells 2021, 10, 2982. https://doi.org/10.3390/cells10112982
Dunbar H, Weiss DJ, Rolandsson Enes S, Laffey JG, English K. The Inflammatory Lung Microenvironment; a Key Mediator in MSC Licensing. Cells. 2021; 10(11):2982. https://doi.org/10.3390/cells10112982
Chicago/Turabian StyleDunbar, Hazel, Daniel J Weiss, Sara Rolandsson Enes, John G Laffey, and Karen English. 2021. "The Inflammatory Lung Microenvironment; a Key Mediator in MSC Licensing" Cells 10, no. 11: 2982. https://doi.org/10.3390/cells10112982
APA StyleDunbar, H., Weiss, D. J., Rolandsson Enes, S., Laffey, J. G., & English, K. (2021). The Inflammatory Lung Microenvironment; a Key Mediator in MSC Licensing. Cells, 10(11), 2982. https://doi.org/10.3390/cells10112982