
Zn2+-Dependent Nuclease Is Involved in Nuclear Degradation during the Programmed Cell Death of Secretory Cavity Formation in Citrus grandis ‘Tomentosa’ Fruits



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Sampling
2.2. Quantitative Real-Time PCR Analysis of CgENDO1
2.3. Cloning and Sequence Analysis of CgENDO1
2.4. In Vitro Expression of CgENDO1 in Escherichia coli and Enzyme Activity Assay
2.5. Morphological and Developmental Observation and Immunocytochemical Localization
2.6. CgENDO1 Expression Analysis by In Situ Hybridization
2.7. Zn2+ Ions Subcellular Localization
3. Results
3.1. Zn2+-Dependent Nuclease CgENDO1 Degrades DNA
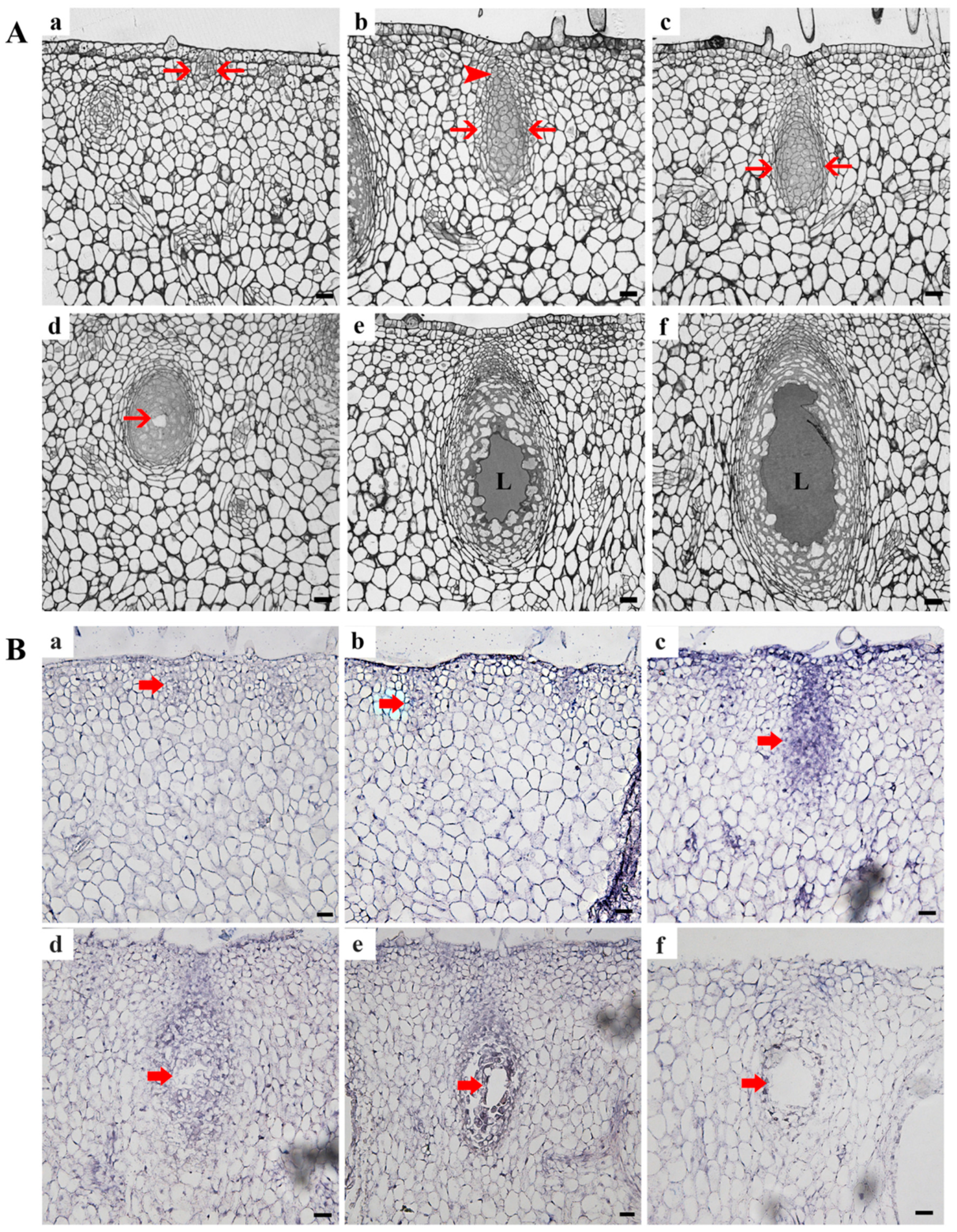
3.2. Molecular and Cytological Characteristics of Zn2+-Dependent Nuclease CgENDO1 in the Secretory Cavity Cells PCD in C. grandis ‘Tomentosa’ Fruits
4. Discussion
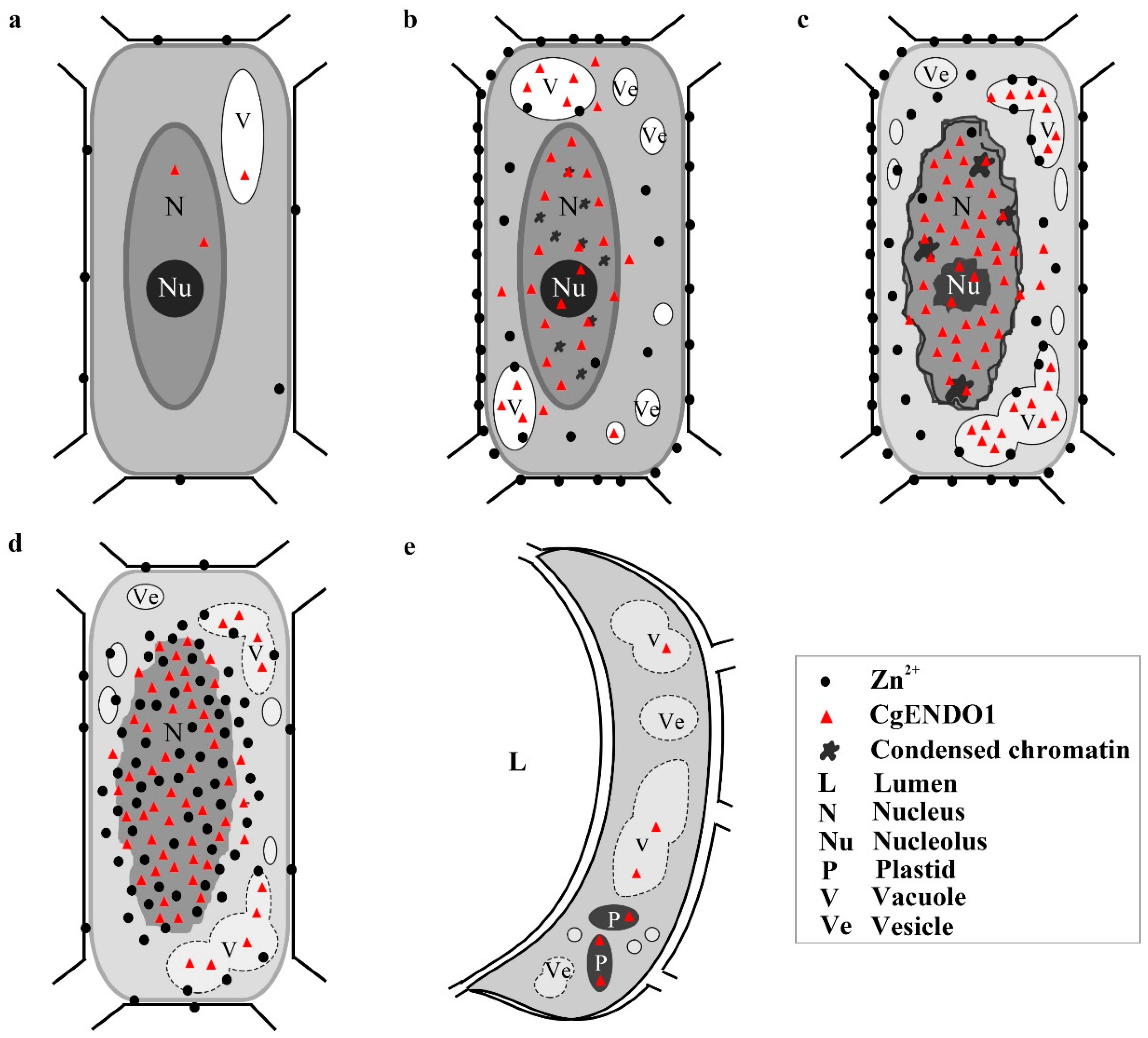
4.1. Zn2+-Dependent Nuclease CgENDO1 Is Involved in the Nuclear DNA Degradation Process of Secretory Cavity Cell PCD
4.2. The Zn2+-Dependent Nuclease CgENDO1 and the Ca2+-Dependent Nuclease CgCAN May Play a Synergistic Role in the Process of Nuclear DNA Degradation
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Pennell, R.I.; Lamb, C. Programmed cell death in plants. Plant Cell. 1997, 9, 1157–1168. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.M. Programmed cell death in development and defense. Plant Physiol. 2001, 125, 94–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filonova, L.H.; von Arnold, S.; Daniel, G.; Bozhkov, P.V. Programmed cell death eliminates all but one embryo in a polyembryonic plant seed. Cell Death Differ. 2002, 9, 1057–1062. [Google Scholar] [CrossRef]
- Gavrieli, Y.; Sherman, Y.; Ben-Sasson, S.A. Identification of programmed cell death in situ via specific labeling of nuclear DNA fragmentation. J. Cell Biol. 1992, 119, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Van Doorn, W.G. Classes of programmed cell death in plants, compared to those in animals. J. Exp. Bot. 2011, 62, 4749–4761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kihlmark, M.; Imreh, G.; Hallberg, E. Sequential degradation of proteins from the nuclear envelope during apoptosis. J. Cell Sci. 2001, 114, 3643–3653. [Google Scholar] [CrossRef]
- Toné, S.; Sugimoto, K.; Tanda, K.; Suda, T.; Uehira, K.; Kanouchi, H.; Samejima, K.; Minatogawa, Y.; Earnshaw, W.C. Three distinct stages of apoptotic nuclear condensation revealed by time-lapse imaging, biochemical and electron microscopy analysis of cell-free apoptosis. Exp. Cell Res. 2007, 313, 3635–3644. [Google Scholar] [CrossRef] [Green Version]
- Shiokawa, D.; Ohyama, H.; Yamada, T.; Takahashi, K.; Tanuma, S. Identification of an endonuclease responsible for apoptosis in rat thymocytes. Eur. J. Biochem. 1994, 226, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, P.; Widlak, P.; Zou, H.; Luo, X.; Garrard, W.T.; Wang, X. The 40-kDa subunit of DNA fragmentation factor induces DNA fragmentation and chromatin condensation during apoptosis. Proc. Natl. Acad. Sci. USA 1998, 95, 8461–8466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minina, E.A.; Coll, N.S.; Tuominen, H.; Bozhkov, P.V. Metacaspases versus caspases in development and cell fate regulation. Cell Death Differ. 2017, 24, 1314–1325. [Google Scholar] [CrossRef] [PubMed]
- Elbaz, M.; Avni, A.; Weil, M. Constitutive caspase-like machinery executes programmed cell death in plant cells. Cell Death Differ. 2002, 9, 726–733. [Google Scholar] [CrossRef]
- Huai, B.; Bai, M.; Tong, P.P.; He, H.J.; Liang, M.J.; Chen, C.Y.; Wu, H. CgPBA1 may be involved in nuclear degradation during secretory cavity formation by programmed cell death in Citrus grandis ‘Tomentosa’ fruits. Plant Physiol. Biochem. 2021, 160, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Ito, J.; Fukuda, H. ZEN1 is a key enzyme in the degradation of nuclear DNA during programmed cell death of tracheary elements. Plant Cell. 2002, 14, 3201–3211. [Google Scholar] [CrossRef] [Green Version]
- Bai, M.; Liang, M.; Huai, B.; Gao, H.; Tong, P.; Shen, R.; He, H.; Wu, H. Ca2+-dependent nuclease is involved in DNA degradation during the formation of the secretory cavity by programmed cell death in fruit of Citrus grandis ‘Tomentosa’. J. Exp. Bot. 2020, 71, 4812–4827. [Google Scholar] [CrossRef]
- Sugiyama, M.; Ito, J.; Aoyagi, S.; Fukuda, H. Endonucleases. Plant Mol. Biol. 2000, 44, 387–397. [Google Scholar] [CrossRef]
- Maret, W. Zinc biochemistry: From a single zinc enzyme to a key element of life. Adv. Nutr. 2013, 4, 82–91. [Google Scholar] [CrossRef] [Green Version]
- Fukada, T.; Kambe, T. Zinc Signals in Cellular Functions and Disorders; Springer: Berlin/Heidelberg, Germany, 2014; ISBN 978-4-431-55114-0. [Google Scholar]
- Peck, E.J., Jr.; Ray, W.J., Jr. Metal complexes of phosphoglucomutase in vivo. Alterations induced by insulin. J. Biol. Chem. 1971, 246, 1160–1167. [Google Scholar] [CrossRef]
- Krężel, A.; Maret, W. The biological inorganic chemistry of zinc ions. Arch. Biochem. Biophys. 2016, 611, 3–19. [Google Scholar] [CrossRef] [Green Version]
- Desai, N.A.; Shankar, V. Single-strand-specific nucleases. FEMS Microbiol. Rev. 2003, 26, 457–491. [Google Scholar] [CrossRef]
- Lesniewicz, K.; Karlowski, W.M.; Pienkowska, J.R.; Krzywkowski, P.; Poreba, E. The plant s1-like nuclease family has evolved a highly diverse range of catalytic capabilities. Plant Cell Physiol. 2013, 54, 1064–1078. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Amador, M.A.; Abler, M.L.; De Rocher, E.J.; Thompson, D.M.; van Hoof, A.; LeBrasseur, N.D.; Lers, A.; Green, P.J. Identification of BFN1, a bifunctional nuclease induced during leaf and stem senescence in Arabidopsis. Plant Physiol. 2000, 122, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Triques, K.; Sturbois, B.; Gallais, S.; Dalmais, M.; Chauvin, S.; Clepet, C.; Aubourg, S.; Rameau, C.; Caboche, M.; Bendahmane, A. Characterization of Arabidopsis thaliana mismatch specific endonucleases: Application to mutation discovery by TILLING in pea. Plant J. 2007, 51, 1116–1125. [Google Scholar] [CrossRef] [PubMed]
- Farage-Barhom, S.; Burd, S.; Sonego, L.; Mett, A.; Belausov, E.; Gidoni, D.; Lers, A. Localization of the Arabidopsis senescence- and cell death-associated BFN1 nuclease: From the ER to fragmented nuclei. Mol. Plant. 2011, 4, 1062–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, C.Y.; Lai, Y.L.; Liu, W.Y.; Lin, C.H.; Chen, Y.T.; Chen, L.F.; Lin, T.Y.; Shaw, J.F. Arabidopsis ENDO2: Its catalytic role and requirement of N-glycosylation for function. J. Agric. Food Chem. 2012, 60, 5169–5179. [Google Scholar] [CrossRef]
- Matallana-Ramirez, L.P.; Rauf, M.; Farage-Barhom, S.; Dortay, H.; Xue, G.P.; Dröge-Laser, W.; Lers, A.; Balazadeh, S.; Mueller-Roeber, B. NAC transcription factor ORE1 and senescence-induced BIFUNCTIONAL NUCLEASE1 (BFN1) constitute a regulatory cascade in Arabidopsis. Mol. Plant. 2013, 6, 1438–1452. [Google Scholar] [CrossRef] [Green Version]
- Granot, G.; Morgenstern, Y.; Khan, A.; Rapp, Y.G.; Pesok, A.; Nevo, E.; Grafi, G. Internucleosomal DNA fragmentation in wild emmer wheat is catalyzed by S1-type endonucleases translocated to the nucleus upon induction of cell death. Biochim. Biophys. Acta 2015, 1849, 239–246. [Google Scholar] [CrossRef]
- Givaty-Rapp, Y.; Yadav, N.S.; Khan, A.; Grafi, G. S1-Type endonuclease 2 in dedifferentiating Arabidopsis protoplasts: Translocation to the nucleus in senescing protoplasts is associated with De-Glycosylation. PLoS ONE 2017, 12, e0170067. [Google Scholar] [CrossRef] [Green Version]
- Farage-Barhom, S.; Burd, S.; Sonego, L.; Perl-Treves, R.; Lers, A. Expression analysis of the BFN1 nuclease gene promoter during senescence, abscission, and programmed cell death-related processes. J. Exp. Bot. 2008, 59, 3247–3258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belmonte, M.F.; Kirkbride, R.C.; Stone, S.L.; Pelletier, J.M.; Bui, A.Q.; Yeung, E.C.; Hashimoto, M.; Fei, J.; Harada, C.M.; Munoz, M.D.; et al. Comprehensive developmental profiles of gene activity in regions and subregions of the Arabidopsis seed. Proc. Natl. Acad. Sci. USA 2013, 110, E435–E444. [Google Scholar] [CrossRef] [Green Version]
- Doherty, R.; Madhusudan, S. DNA repair endonucleases: Physiological roles and potential as drug targets. J. Biomol. Screen. 2015, 20, 829–841. [Google Scholar] [CrossRef] [Green Version]
- Brown, P.H.; Ho, T.H. Biochemical properties and hormonal regulation of barley nuclease. Eur. J. Biochem. 1987, 168, 357–364. [Google Scholar] [CrossRef]
- Mariani, C.; Beuckeleer, M.; Treuttner, J.; Leemans, J.; Goldberg, R.B. Induction of male-sterility in plants by a chimaeric ribonuclease gene. Nature 1990, 347, 737–741. [Google Scholar] [CrossRef]
- Aoyagi, S.; Sugiyama, M.; Fukuda, H. BEN1 and ZEN1 cDNAs encoding S1-type DNases that are associated with programmed cell death in plants. FEBS Lett. 1998, 429, 134–138. [Google Scholar] [CrossRef] [Green Version]
- Zaina, G.; Morassutti, C.; De Amicis, F.; Fogher, C.; Marchetti, S. Endonuclease genes up-regulated in tissues undergoing programmed cell death are expressed during male gametogenesis in barley. Gene 2003, 315, 43–50. [Google Scholar] [CrossRef]
- Domínguez, F.; Cejudo, F.J. Identification of a nuclear-localized nuclease from wheat cells undergoing programmed cell death that is able to trigger DNA fragmentation and apoptotic morphology on nuclei from human cells. Biochem. J. 2006, 397, 529–536. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z. Study on the relationship between structure and development of medicinal plants and accumulation of main medicinal components. Chin. Wild Plant Resour. 2005, 24, 8–12. [Google Scholar]
- Liang, S.J.; Wang, H.Y.; Yang, M.; Wu, H. Sequential actions of pectinases and cellulases during secretory cavity formation in Citrus fruits. Trees Struc. Funct. 2009, 23, 19–27. [Google Scholar] [CrossRef]
- Tong, P.; Huai, B.; Chen, Y.; Bai, M.; Wu, H. CisPG21 and CisCEL16 are involved in the regulation of the degradation of cell walls during secretory cavity cell programmed cell death in the fruits of Citrus sinensis (L.) Osbeck. Plant Sci. 2020, 297, 110540. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wu, H. Programmed cell death involved in the schizolysigenous formation of the secretory cavity in Citrus sinensis (L.) Osbeck. Chin. Sci. Bull. 2010, 55, 2160–2168. [Google Scholar] [CrossRef]
- Liu, P.W.; Liang, S.J.; Yao, N.; Wu, H. Programmed cell death of secretory cavity cells in fruits of Citrus grandis cv. Tomentosa is associated with activation of caspase 3-like protease. Trees Struc. Funct. 2012, 26, 1821–1835. [Google Scholar] [CrossRef]
- Ren, H.; Zhao, X.; Li, W.; Hussain, J.; Qi, G.; Liu, S. Calcium signaling in plant programmed cell death. Cells. 2021, 10, 1089. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhan, Q.; Shi, T.; Liu, J.; Zhao, K.; Gao, Y. The nuclear transporter SAD2 plays a role in calcium- and H2O2 -mediated cell death in Arabidopsis. Plant J. 2020, 101, 324–333. [Google Scholar] [CrossRef]
- Pihl, E. Ultrastructural localization of heavy metals by a modified sulfide-silver method. Histochemie 1967, 10, 126–139. [Google Scholar] [CrossRef]
- Volbeda, A.; Lahm, A.; Sakiyama, F.; Suck, D. Crystal structure of Penicillium citrinum P1 nuclease at 2.8 A resolution. EMBO J. 1991, 10, 1607–1618. [Google Scholar] [CrossRef]
- Thelen, M.P.; Northcote, D.H. Identification and purification of a nuclease from Zinnia elegans L.: A potential molecular marker for xylogenesis. Planta 1989, 179, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, F.; Cejudo, F.J. A comparison between nuclear dismantling during plant and animal programmed cell death. Plant Sci. 2012, 197, 114–121. [Google Scholar] [CrossRef] [Green Version]
- Leśniewicz, K.; Pieńkowska, J.; Poreba, E. Characterization of nucleases involved in seedling development of cauliflower. J. Plant Physiol. 2010, 167, 1093–1100. [Google Scholar] [CrossRef]
- Sakamoto, W.; Takami, T. Nucleases in higher plants and their possible involvement in DNA degradation during leaf senescence. J. Exp. Bot. 2014, 65, 3835–3843. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Bai, M.; Chen, Y.; Liu, P.W.; Gao, L.; Liang, S.J.; Wu, H. Programmed cell death of secretory cavity cells of Citrus fruits is associated with Ca2+ accumulation in the nucleus. Trees Struct. Funct. 2014, 28, 1137–1144. [Google Scholar] [CrossRef]
- Martin, J.B.; Bligny, R.; Rebeille, F.; Douce, R.; Leguay, J.J.; Mathieu, Y.; Guern, J. A P Nuclear magnetic resonance study of intracellular pH of plant cells cultivated in liquid medium. Plant Physiol. 1982, 70, 1156–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara-Nishimura, I.; Hatsugai, N. The role of vacuole in plant cell death. Cell Death Differ. 2011, 18, 1298–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, M.; Bai, M.; Wu, H. Zn2+-Dependent Nuclease Is Involved in Nuclear Degradation during the Programmed Cell Death of Secretory Cavity Formation in Citrus grandis ‘Tomentosa’ Fruits. Cells 2021, 10, 3222. https://doi.org/10.3390/cells10113222
Liang M, Bai M, Wu H. Zn2+-Dependent Nuclease Is Involved in Nuclear Degradation during the Programmed Cell Death of Secretory Cavity Formation in Citrus grandis ‘Tomentosa’ Fruits. Cells. 2021; 10(11):3222. https://doi.org/10.3390/cells10113222
Chicago/Turabian StyleLiang, Minjian, Mei Bai, and Hong Wu. 2021. "Zn2+-Dependent Nuclease Is Involved in Nuclear Degradation during the Programmed Cell Death of Secretory Cavity Formation in Citrus grandis ‘Tomentosa’ Fruits" Cells 10, no. 11: 3222. https://doi.org/10.3390/cells10113222
APA StyleLiang, M., Bai, M., & Wu, H. (2021). Zn2+-Dependent Nuclease Is Involved in Nuclear Degradation during the Programmed Cell Death of Secretory Cavity Formation in Citrus grandis ‘Tomentosa’ Fruits. Cells, 10(11), 3222. https://doi.org/10.3390/cells10113222