
Neuronal Menin Overexpression Rescues Learning and Memory Phenotype in CA1-Specific α7 nAChRs KD Mice


Abstract
:1. Introduction
2. Methodology
2.1. Animals: Brain Slices and Neuronal Cell Culture
2.2. Immunocytochemistry and Immunohistochemistry
2.3. Quantitative PCR (qPCR)
2.4. Confocal Microscopy
2.5. AAV Production and Transduction of Neuronal Cultures
α7 shRNA AAV and α7 shRNA Scrambled MEN1 Encoding AAV and GFP Only AAV
2.6. Plasmid Construction and AAV Packaging
2.7. Stereotaxic Injections in C57BL/6 Mice
2.8. Contextual Fear Conditioning Behaviour Test
2.9. Experimental Design and Statistical Analysis
3. Results
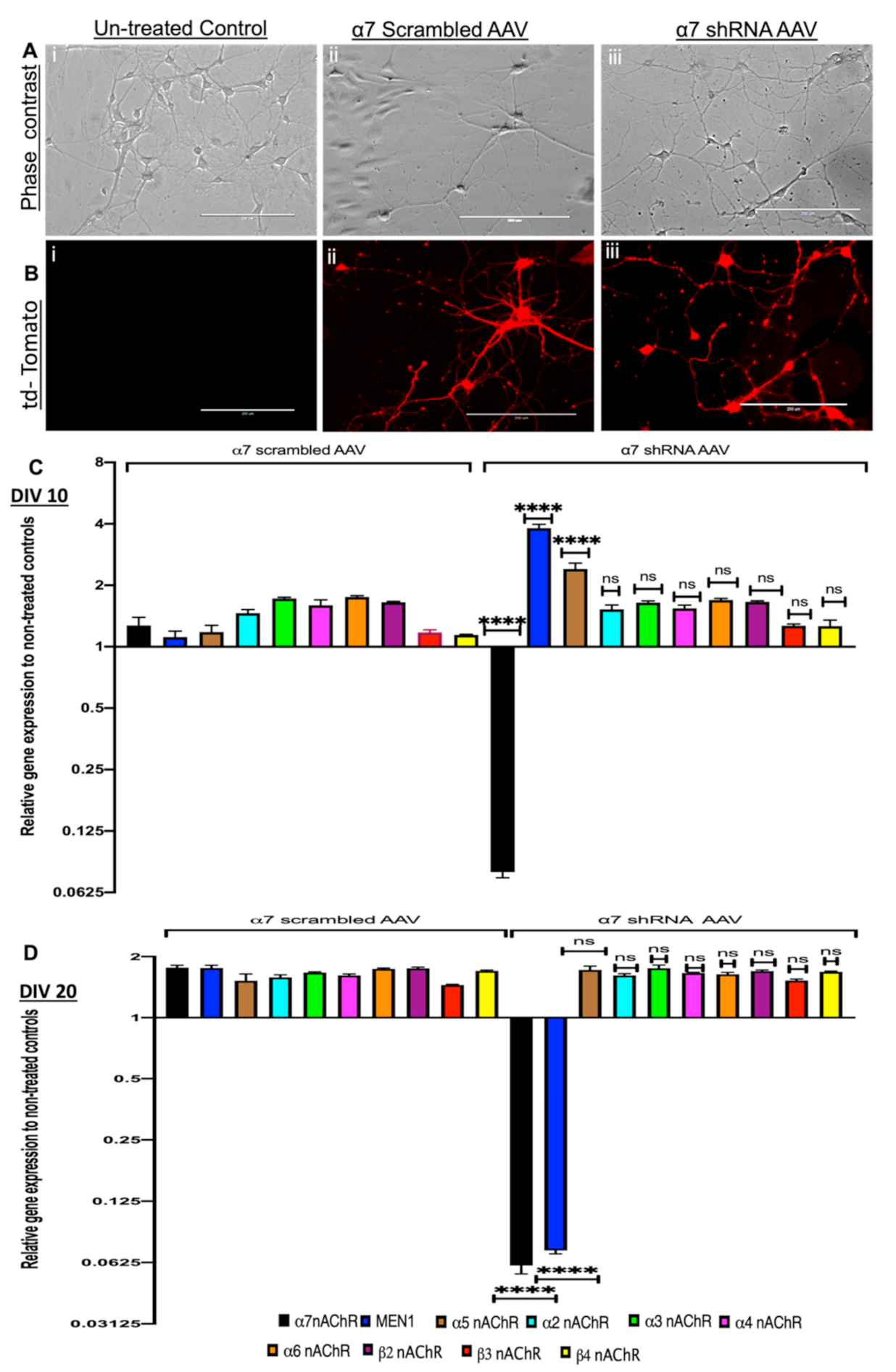
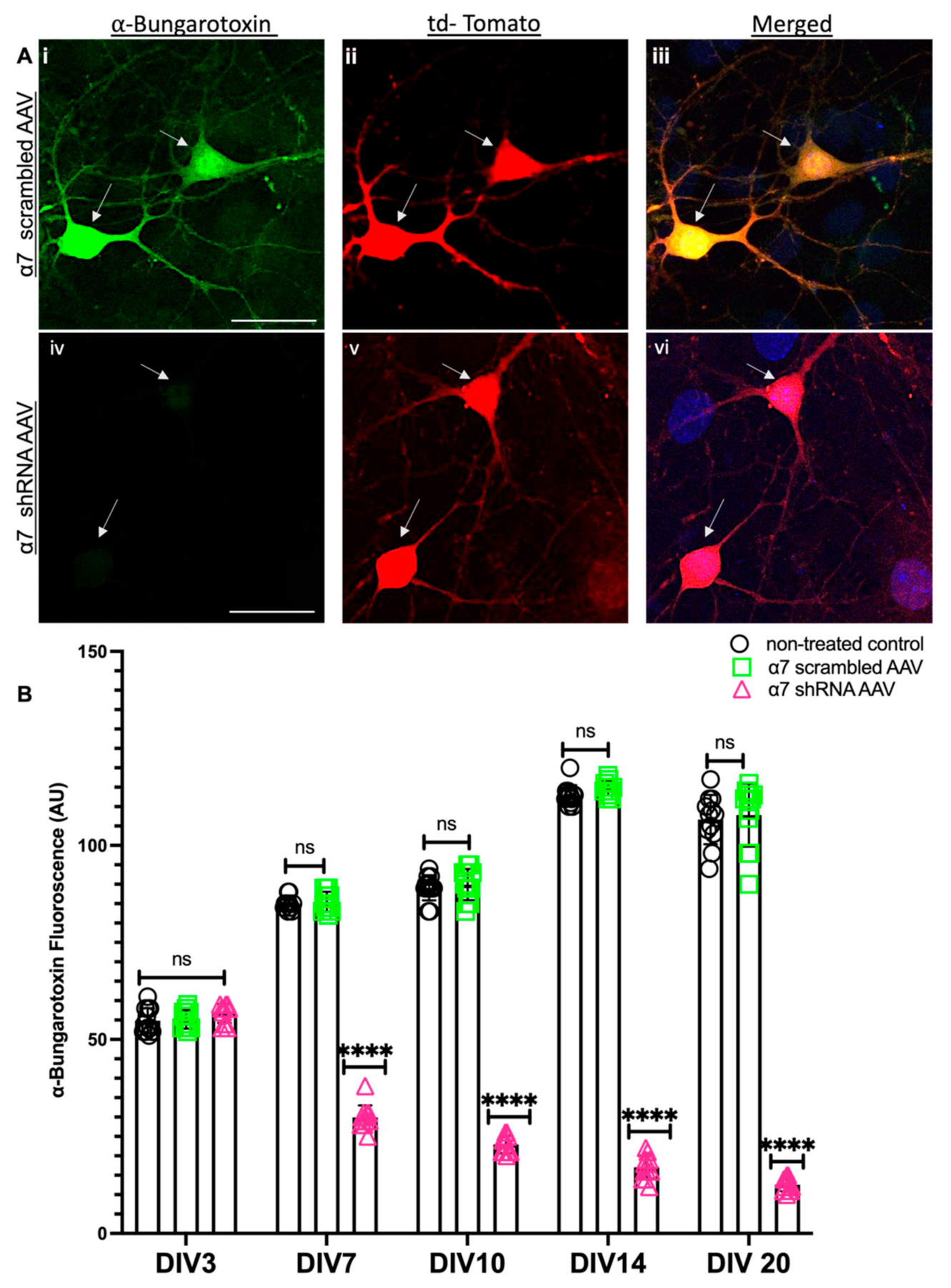
3.1. Selective Hippocampal, Neuron Specific KD of α7 nAChRs Differentially Regulates the Expression of the MEN1 Gene during Synaptogenesis and Synaptic Maturation
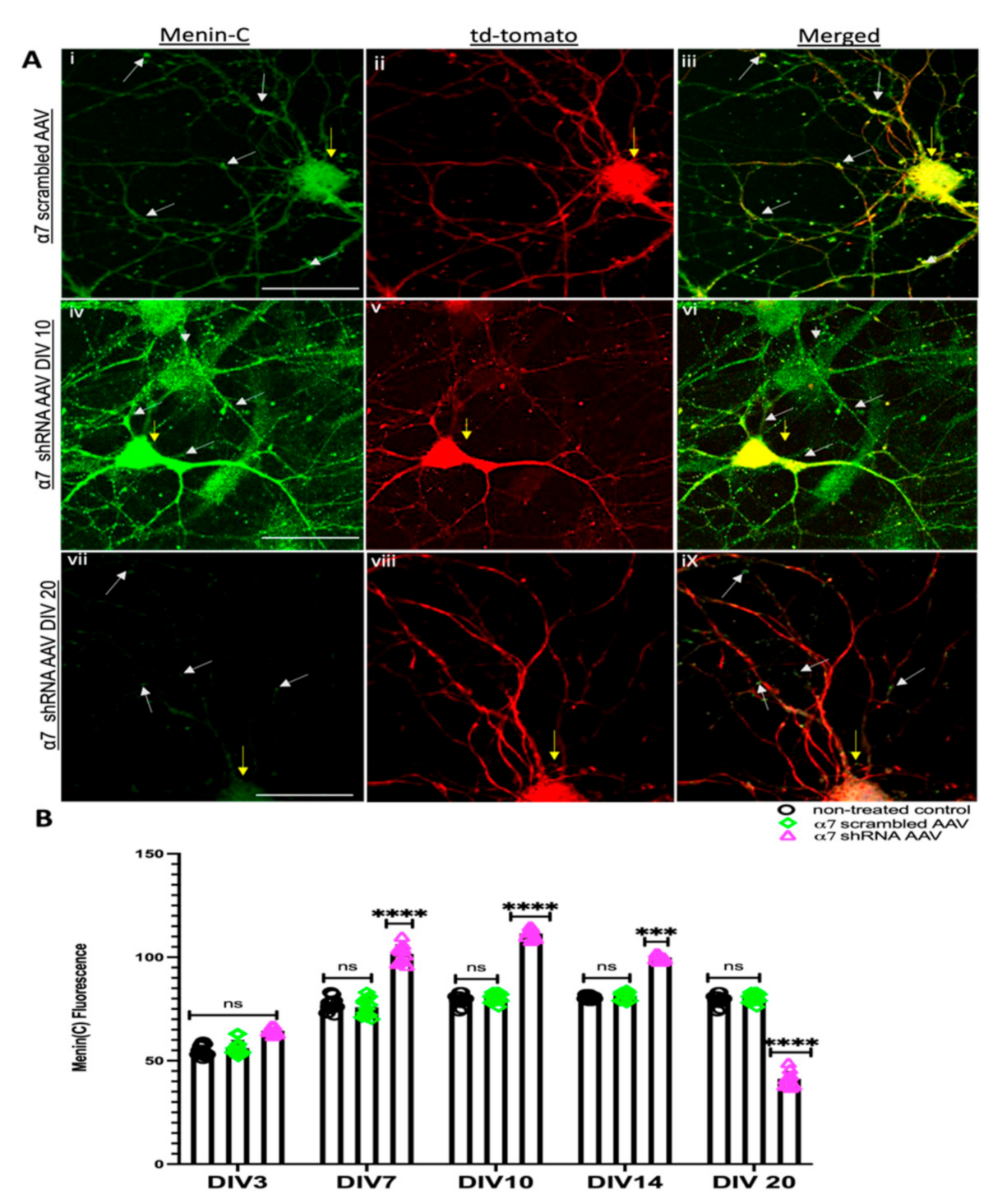
3.2. α7 nAChRs KD in Hippocampal Neurons Perturbs Menin Expression
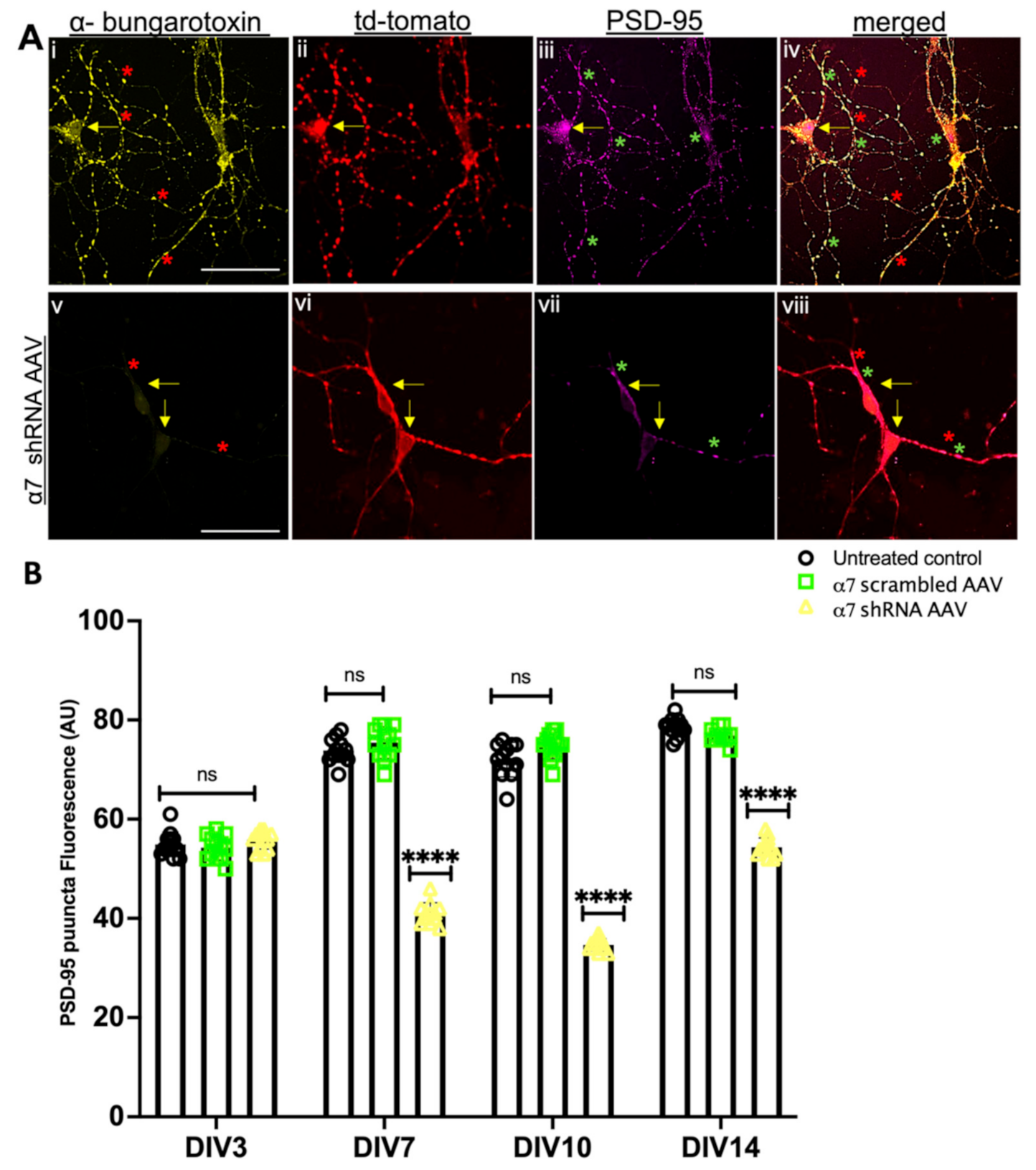
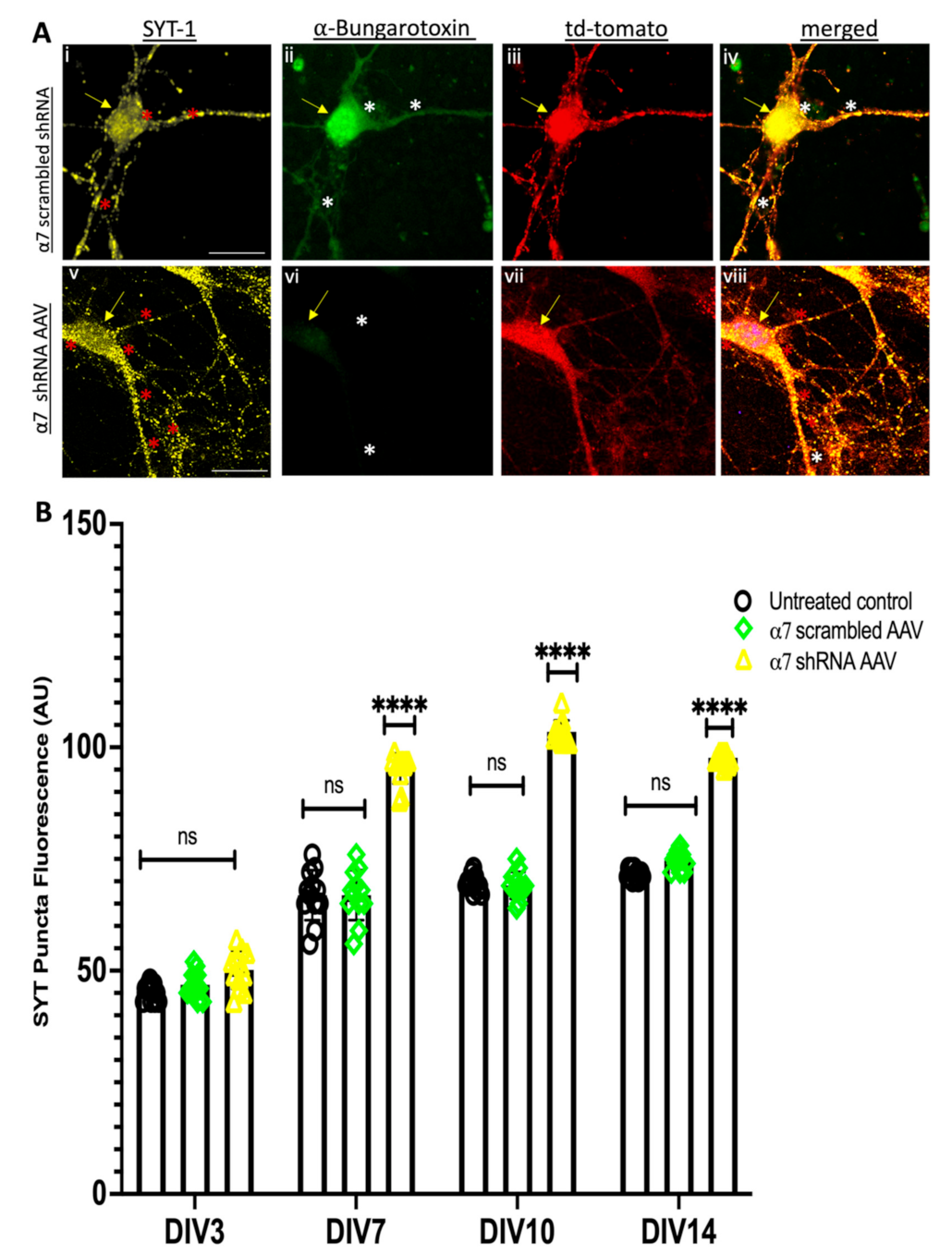
3.3. α7 nAChRs KD in Hippocampal Neuronal Cultures Altered Synaptic Proteins Assembly at the Pre and Postsynaptic Sites
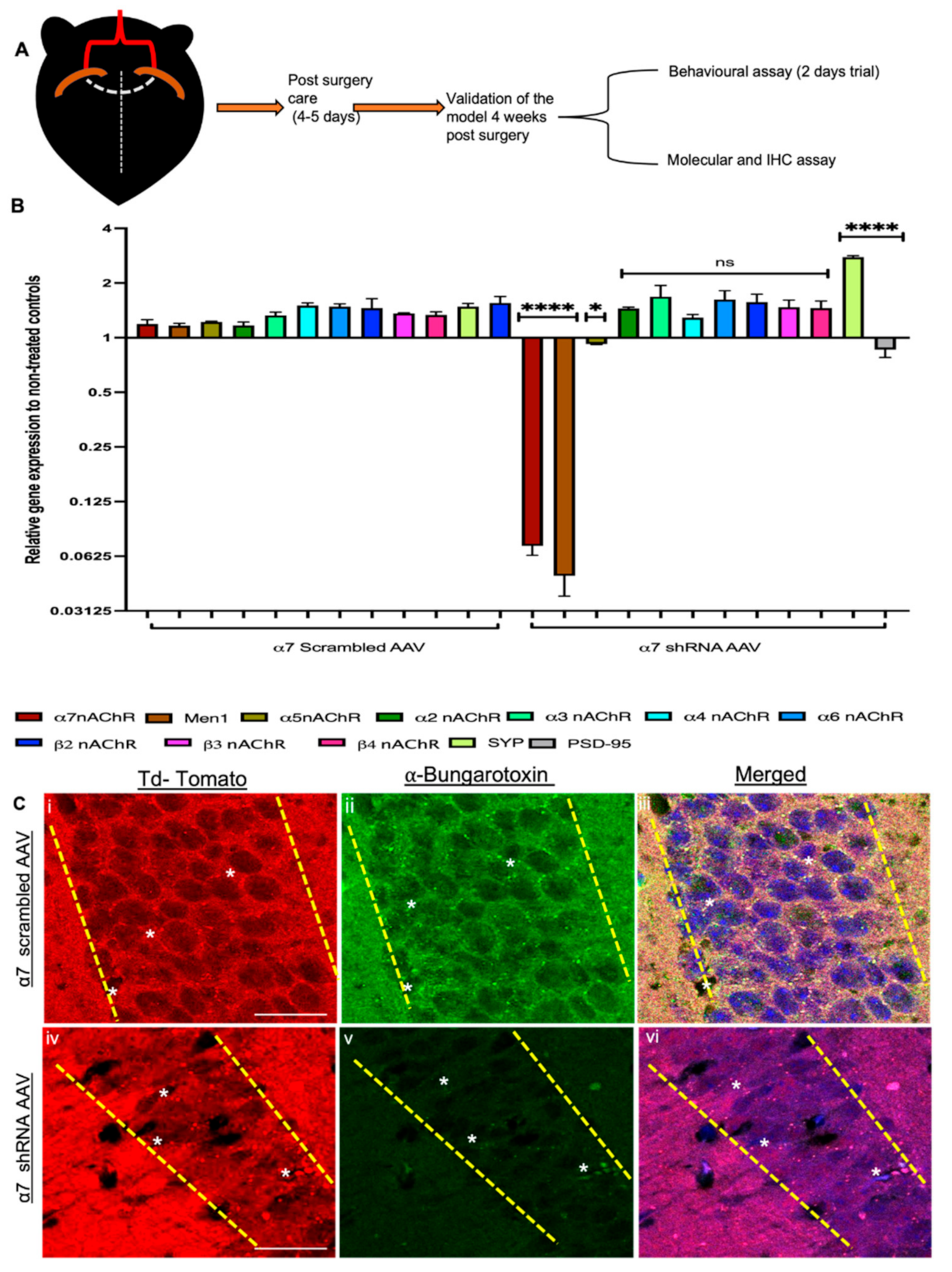
3.4. α7 nAChRs KD Altered MEN1 Gene Expression and Synaptic Assembly in α7 nAChRs shRNA AAV (CA1 Specific) KD Mouse
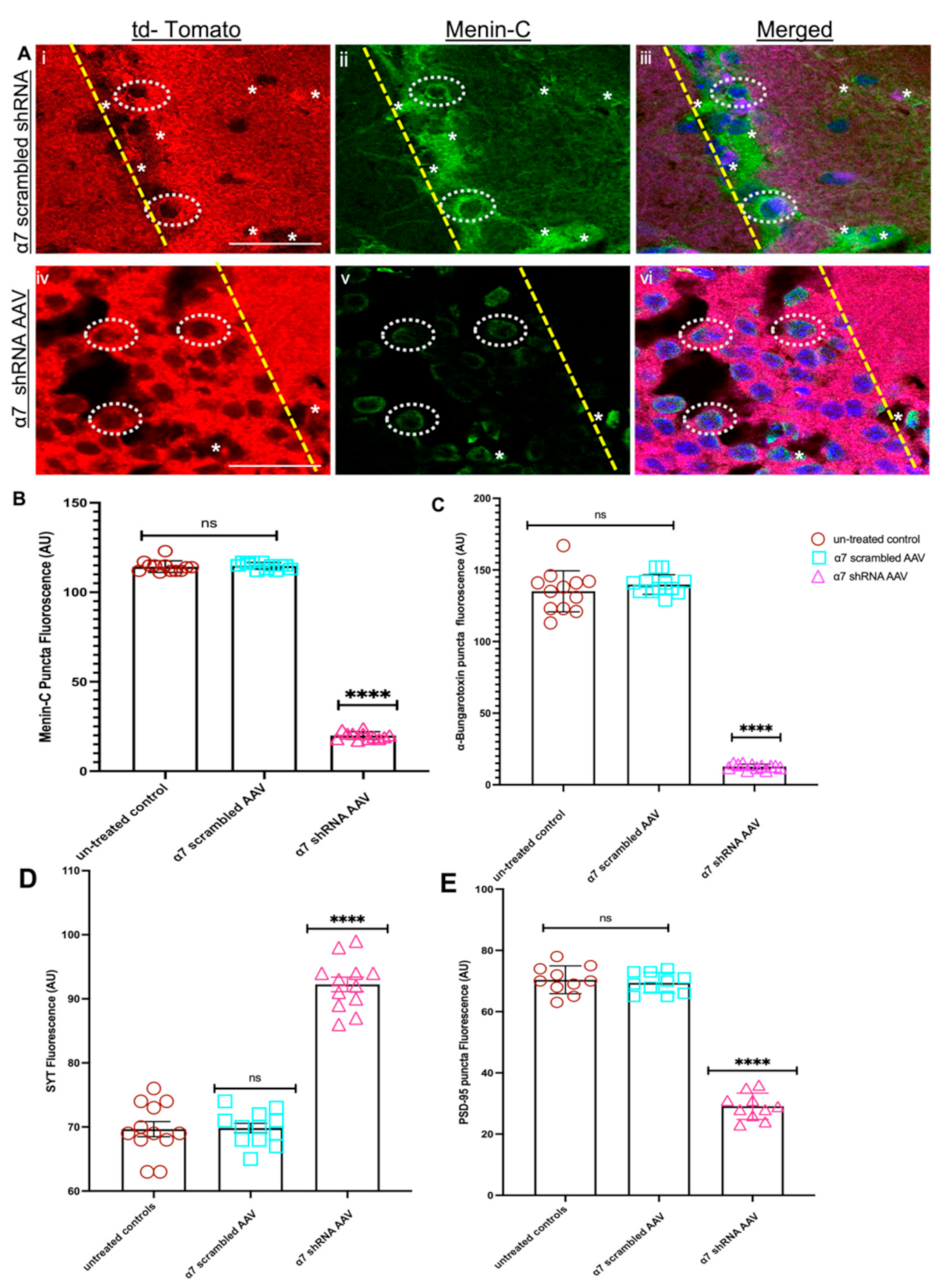
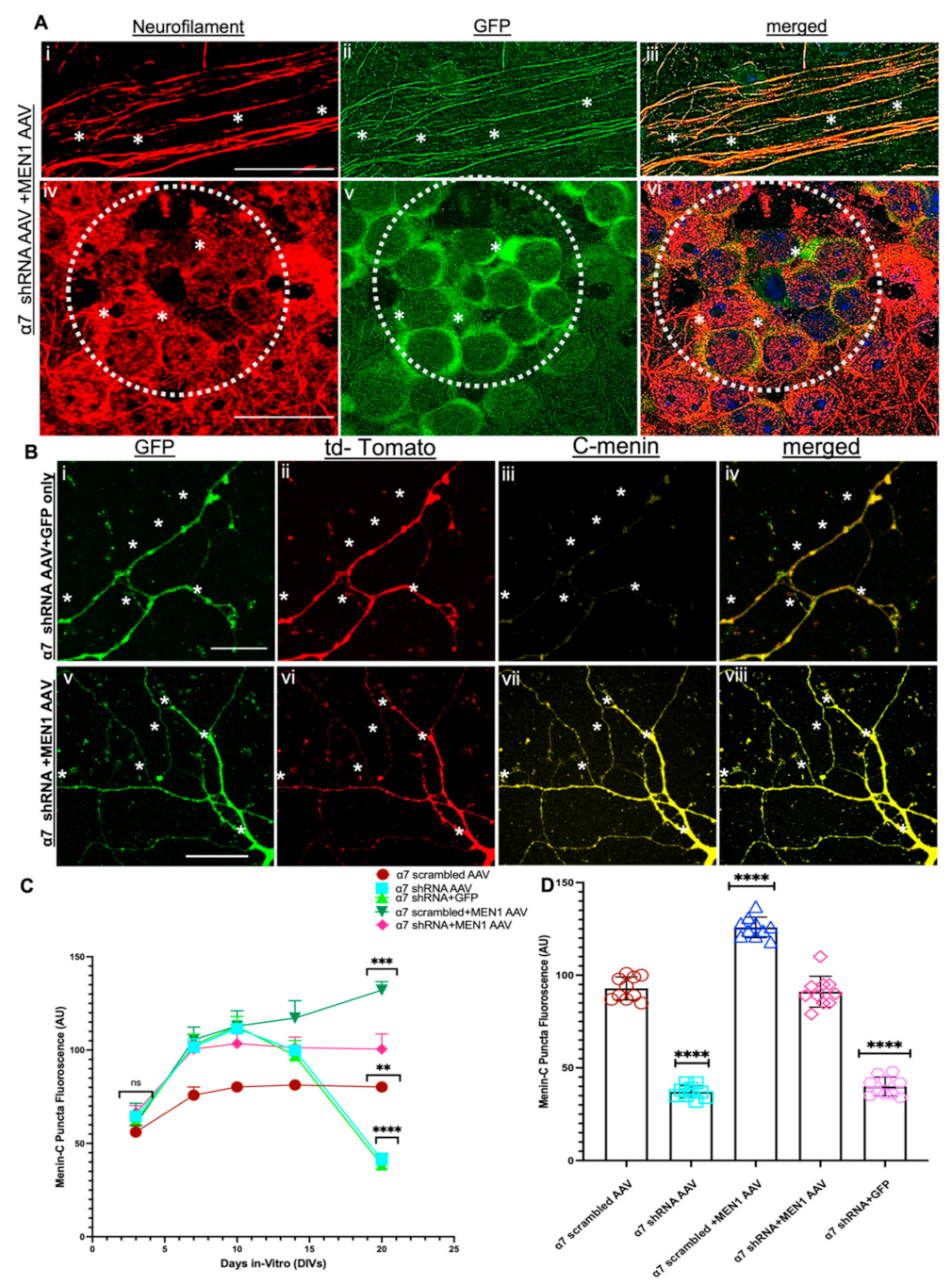
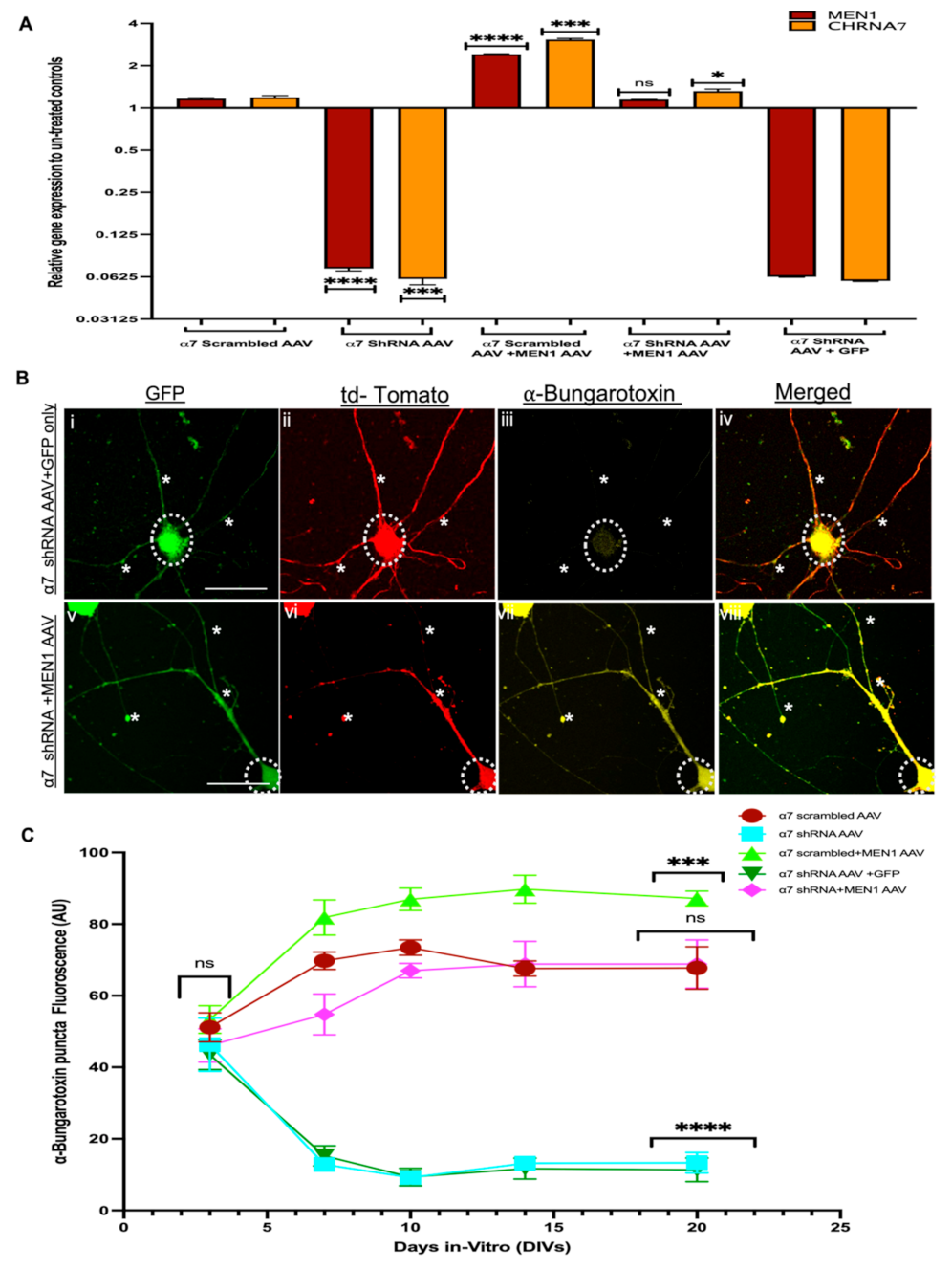
3.5. Restoring Menin in a7 nAChRs KD Hippocampal Neurons Rescued the Expression of a7 nAChRs and Clustering at Synaptic Sites
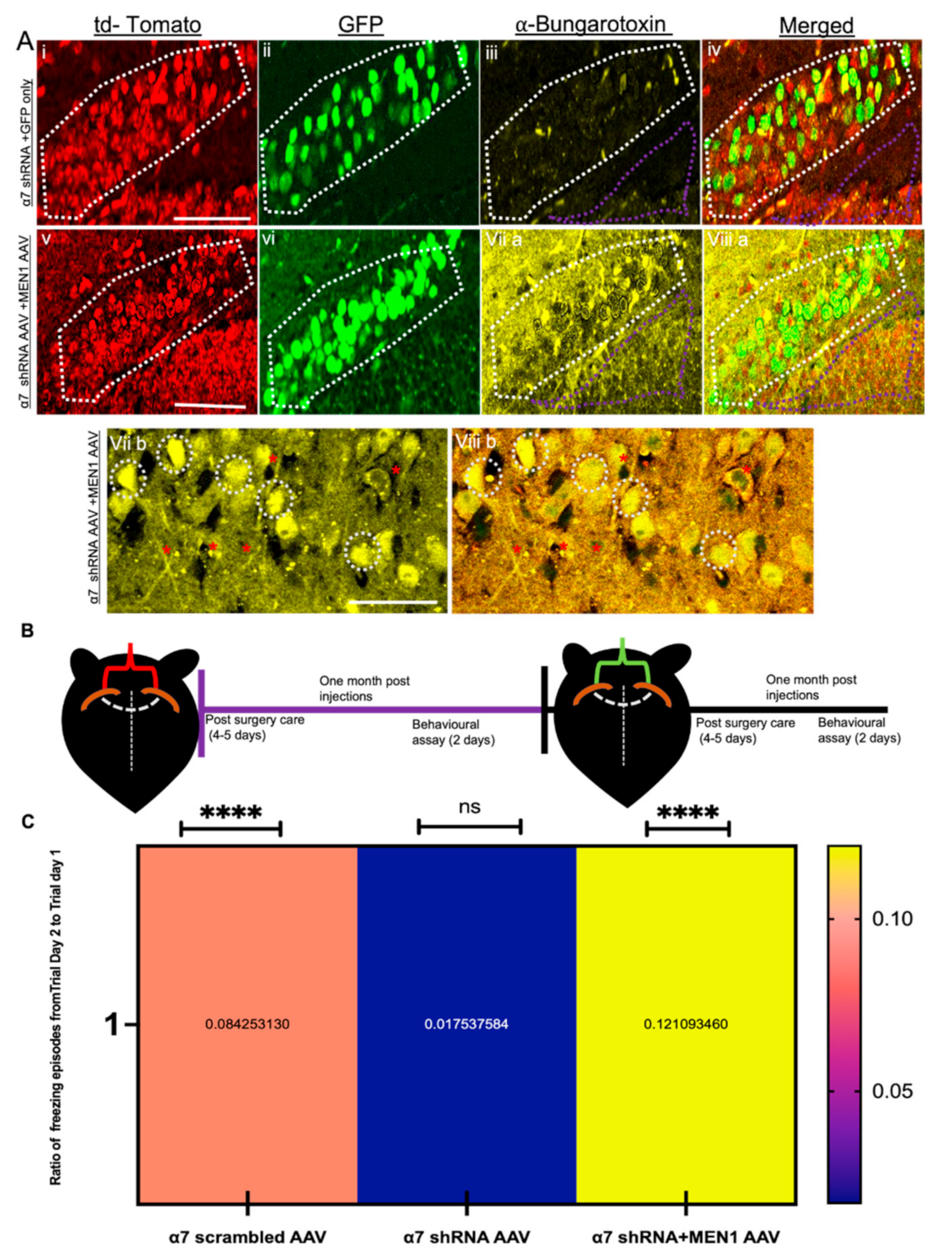
3.6. Overexpression of Exogenous Menin in the α7 nAChRs KD Mice Rescues Hippocampus Dependent Learning and Memory

4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Corradi, J.; Bouzat, C. Understanding the Bases of Function and Modulation of α7 Nicotinic Receptors: Implications for Drug Discovery. Mol. Pharmacol. 2016, 90, 288–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hackett, S.F.; Seidel, C.; Abraham, S.; Chadha, R.; Fortmann, S.D.; Campochiaro, P.A.; Cooke, J.P. The Nicotinic Cholinergic Pathway Contributes to Retinal Neovascularization in a Mouse Model of Retinopathy of Prematurity. Investig. Ophthalmol. Vis. Sci. 2017, 58, 1296–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Q.; Yakel, J.L. The effect of α7 nicotinic receptor activation on glutamatergic transmission in the hippocampus. Biochem. Pharmacol. 2015, 97, 439–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, S.; Yamada, J.; Ueno, S.; Kubota, H.; Furukawa, T.; Yamamoto, S.; Fukuda, A. Insertion of 7 Nicotinic Receptors at Neocortical Layer V GABAergic Synapses Is Induced by a Benzodiazepine, Midazolam. Cereb. Cortex 2006, 17, 653–660. [Google Scholar] [CrossRef]
- Bürli, T.; Baer, K.; Ewers, H.; Sidler, C.; Fuhrer, C.; Fritschy, J.M. Single particle tracking of alpha7 nicotinic AChR in hippocampal neurons reveals regulated confinement at glutamatergic and GABAergic perisynaptic sites. PLoS ONE 2010, 5, e11507. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Z.; Wang, J.; Wang, Y.; Henderson, Z.; Wang, X.; Zhang, X.; Song, J.; Lu, C. The modulation of nicotinic acetylcholine receptors on the neuronal network oscillations in rat hippocampal CA3 area. Sci. Rep. 2015, 5, 9493. [Google Scholar] [CrossRef] [Green Version]
- Hone, A.J.; McIntosh, J.M. Nicotinic acetylcholine receptors in neuropathic and inflammatory pain. FEBS Lett. 2018, 592, 1045–1062. [Google Scholar] [CrossRef] [Green Version]
- Mineur, Y.S.; Mose, T.N.; Blakeman, S.; Picciotto, M.R. Hippocampal α7 nicotinic ACh receptors contribute to modulation of depression-like behaviour in C57BL/6J mice. Br. J. Pharmacol. 2018, 175, 1903–1914. [Google Scholar] [CrossRef] [Green Version]
- Maurer, S.V.; Williams, C.L. The cholinergic system modulates memory and hippocampal plasticity via its interactions with non-neuronal cells. Front. Immunol. 2017, 8, 1489. [Google Scholar] [CrossRef] [Green Version]
- Yakel, J.L. Nicotinic ACh receptors in the hippocampal circuit; functional expression and role in synaptic plasticity. J. Physiol. 2014, 592, 4147–4153. [Google Scholar] [CrossRef]
- Gu, Z.; Yakel, J.L. Timing-Dependent Septal Cholinergic Induction of Dynamic Hippocampal Synaptic Plasticity. Neuron 2011, 71, 155–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, J.-X.; Yakel, J.L. Nicotinic acetylcholine receptor-mediated calcium signaling in the nervous system. Acta Pharmacol. Sin. 2009, 30, 673–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Dai, A.; Zhou, Y.; Peng, G.; Hu, G.; Li, B.; Sham, J.; Ran, P. Nicotine Elevated Intracellular Ca2+ in Rat Airway Smooth Muscle Cells via Activating and Up-Regulating α7-Nicotinic Acetylcholine Receptor. Cell. Physiol. Biochem. 2014, 33, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Dineley, K.T.; Pandya, A.A.; Yakel, J.L. Nicotinic ACh receptors as therapeutic targets in CNS disorders. Trends Pharmacol. Sci. 2014, 36, 96–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godin, J.-R.; Roy, P.; Quadri, M.; Bagdas, D.; Toma, W.; Narendrula-Kotha, R.; Kishta, O.A.; Damaj, M.I.; Horenstein, N.A.; Papke, R.L.; et al. A silent agonist of α7 nicotinic acetylcholine receptors modulates inflammation ex vivo and attenuates EAE. Brain Behav. Immun. 2020, 87, 286–300. [Google Scholar] [CrossRef]
- Thomsen, M.S.; EI-Sayed, M.; Mikkelsen, J.D. Differential immediate and sustained memory enhancing effects of alpha7 nicotinic receptor agonists and allosteric modulators in rats. PLoS ONE 2011, 6, e27014. [Google Scholar] [CrossRef] [Green Version]
- Levin, D.E.; Hall, B.J.; Rezvani, A.H. Heterogeneity Across Brain Regions and Neurotransmitter Interactions with Nicotinic Effects on Memory Function. In The Neurobiology and Genetics of Nicotine and Tobacco; Springer: Cham, Switzerland, 2015; pp. 87–101. [Google Scholar]
- Martin, L.F.a.F.R. Schizophrenia and the alpha7 nicotinic acetylcholine receptor. Int. Rev. Neurobiol. 2007, 78, 225–246. [Google Scholar]
- Potasiewicz, A.; Nikiforuk, A.; Holuj, M.; Popik, P. Stimulation of nicotinic acetylcholine alpha7 receptors rescue schizophrenia-like cognitive impairments in rats. J. Psychopharmacol. 2017, 31, 260–271. [Google Scholar] [CrossRef]
- Vallés, A.S.; Borroni, M.V.; Barrantes, F.J. Targeting Brain α7 Nicotinic Acetylcholine Receptors in Alzheimer’s Disease: Rationale and Current Status. CNS Drugs 2014, 28, 975–987. [Google Scholar] [CrossRef]
- Kabbani, N.; Nicholas, R.A. Beyond the Channel: Metabotropic Signaling by Nicotinic Receptors. Trends Pharmacol. Sci. 2018, 39, 354–366. [Google Scholar] [CrossRef]
- Hogg, R.C.; Raggenbass, M.; Bertrand, D. Nicotinic acetylcholine receptors: From structure to brain function. Rev. Physiol. Biochem. Pharmacol. 2003, 147, 1–46. [Google Scholar] [PubMed]
- Maizón, H.B.; Barrentes, F.J. A deep learning-based approach to model anomalous diffusion of membrane proteins: The case of the nicotinic acetylcholine receptor. Brief. Bioinform. 2021. ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Stigloher, C.; Zhan, H.; Zhen, M.; Richmond, J.; Bessereau, J.L. The Presynaptic Dense Projection of the Caenorhabiditis elegans Cholinergic Neuromuscular Junction Localizes Synaptic Vesicles at the Active Zone through SYD-2/Liprin and UNC-10/RIM-Dependent Interactions. J. Neurosci. 2011, 31, 4388–4396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dau, A.; Komal, P.; Truong, M.; Morris, G.; Evans, G.; Nashmi, R. RIC-3 differentially modulates α4β2 and α7 nicotinic receptor assembly, expression, and nicotine-induced receptor upregulation. BMC Neurosci. 2013, 14, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-David, Y.; Mizrachi, T.; Kagan, S.; Krisher, T.; Cohen, E.; Brenner, T.; Treinin, M. RIC-3 expression and splicing regulate nAChR functional expression. Mol. Brain 2016, 9, 47. [Google Scholar] [CrossRef] [Green Version]
- Halevi, S.; Yassin, L.; Eshel, M.; Sala, F.; Sala, S.; Criado, M.; Treinin, M. Conservation within the RIC-3 gene family: Effectors of mammalian nicotinic acetylcholine receptor expression. J. Biol. Chem. 2003, 278, 34411–34417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flynn, N.; Syed, N.I. Src Family Kinases Play a Role in the Functional Clustering of Central Postsynaptic Nicotinic Acetylcholine Receptors. Front. Mar. Sci. 2020, 7, 8. [Google Scholar] [CrossRef] [Green Version]
- Gu, S.; Matta, J.A.; Lord, B.; Harrington, A.W.; Sutton, S.W.; Davini, W.B.; Bredt, D.S. Brain alpha 7 Nicotinic Acetylcholine Receptor Assembly Requires NACHO. Neuron 2016, 89, 948. [Google Scholar] [CrossRef] [Green Version]
- Flynn, N.; Getz, A.; Visser, F.; Janes, T.A.; Syed, N.I. Menin: A Tumor Suppressor That Mediates Postsynaptic Receptor Expression and Synaptogenesis between Central Neurons of Lymnaea stagnalis. PLoS ONE 2014, 9, e111103. [Google Scholar] [CrossRef]
- Getz, A.M.; Xu, F.; Visser, F.; Persson, R.; Syed, N.I. Tumor suppressor menin is required for subunit-specific nAChR α5 transcription and nAChR-dependent presynaptic facilitation in cultured mouse hippocampal neurons. Sci. Rep. 2017, 7, 1768. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekharappa, S.C.; Guru, S.C.; Manickam, P.; Olufemi, S.E.; Collins, F.S.; Emmert-Buck, M.R.; Debelenko, L.V.; Zhuang, Z.; Lubensky, I.A.; Liotta, L.A.; et al. Positional cloning of the gene for multiple endocrine neoplasia-type 1. Science 1997, 276, 404. [Google Scholar] [CrossRef] [PubMed]
- Lemmens, I.; Van de Ven, W.J.; Kas, K.; Zhang, C.X.; Giraud, S.; Wautot, V.; Buisson, N.; De Witte, K.; Salandre, J.; Lenoir, G.; et al. Identification of the Multiple Endocrine Neoplasia Type 1 (MEN1) Gene. Hum. Mol. Genet. 1997, 6, 1177–1183. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Gurung, B.; Wan, B.; Matkar, S.; Veniaminova, N.A.; Wan, K.; Merchant, J.L.; Hua, X.; Lei, M. The same pocket in menin binds both MLL and JUND but has opposite effects on transcription. Nature 2012, 482, 542–546. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.; Parente, F.; Piehl, F.; Farnebo, F.; Quincey, D.; Silins, G.; Bergman, L.; Carle, G.F.; Lemmens, I.; Grimmond, S.; et al. Characterization of the mouse Men1 gene and its expression during development. Oncogene 1998, 17, 2485–2493. [Google Scholar] [CrossRef] [Green Version]
- Matkar, S.; Thiel, A.; Hua, X. Menin: A scaffold protein that controls gene expression and cell signaling. Trends Biochem. Sci. 2013, 38, 394–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Chen, G.; Xue, Q.; Yu, B. Early changes of β-catenins and menins in spinal cord dorsal horn after peripheral nerve injury. Cell. Mol. Neurobiol. 2010, 30, 885–890. [Google Scholar] [CrossRef] [PubMed]
- Van Kesteren, R.E.; Syed, N.I.; Munno, D.W.; Bouwman, J.; Feng, Z.P.; Geraerts, W.P.; Smit, A.B. Synapse Formation between Central Neurons Requires Postsynaptic Expression of the MEN1 Tumor Suppressor Gene. J. Neurosci. 2001, 21, RC161. [Google Scholar] [CrossRef]
- Xu, S.; Wu, H.; Wang, X.; Shen, X.; Guo, X.; Shen, R.; Wang, F. Tumor suppressor menin mediates peripheral nerve injury-induced neuropathic pain through potentiating synaptic plasticity. Neuroscience 2012, 223, 473–485. [Google Scholar] [CrossRef]
- Zhuang, K.; Huang, C.; Leng, L.; Zheng, H.; Gao, Y.; Chen, G.; Ji, Z.; Sun, H.; Hu, Y.; Wu, D.; et al. Neuron-Specific Menin Deletion Leads to Synaptic Dysfunction and Cognitive Impairment by Modulating p35 Expression. Cell Rep. 2018, 24, 701–712. [Google Scholar] [CrossRef] [Green Version]
- Leng, L.; Zhuang, K.; Liu, Z.; Huang, C.; Gao, Y.; Chen, G.; Lin, H.; Hu, Y.; Wu, D.; Shi, M.; et al. Menin Deficiency Leads to Depressive-like Behaviors in Mice by Modulating Astrocyte-Mediated Neuroinflammation. Neuron 2018, 100, 551–563.e557. [Google Scholar] [CrossRef] [Green Version]
- Batool, S.; Zaidi, J.; Akhter, B.; Ulfat, A.K.; Visser, F.; Syed, N.I. Spatiotemporal Patterns of Menin Localization in Developing Murine Brain: Co-Expression with the Elements of Cholinergic Synaptic Machinery. Cells 2021, 10, 1215. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-J.; Cunningham, M.G. Thin Sectioning of Slice Preparations for Immunohistochemistry. J. Vis. Exp. 2007, 3, e194. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.-C.; Chu, S.-W. High Spatio-Temporal-Resolution Detection of Chlorophyll Fluorescence Dynamics from a Single Chloroplast with Confocal Imaging Fluorometer. Plant. Methods 2016, 13, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelossof, R.; Fairchild, L.; Huang, C.H.; Widmer, C.; Sreedharan, V.T.; Sinha, N.; Lai, D.Y.; Guan, Y.; Premsrirut, P.K.; Tschaharganeh, D.F.; et al. Prediction of ultra-potent shRNAs with a sequential classification algorithm. Nat. Biotechnol. 2017, 35, 350–353. [Google Scholar] [CrossRef]
- Bielefeld, P.; Sierra, A.; Encinas, J.M.; Maletic-Savatic, M.; Anderson, A.; Fitzsimons, C.P. A Standardized Protocol for Stereotaxic Intrahippocampal Administration of Kainic Acid Combined with Electroencephalographic Seizure Monitoring in Mice. Front. Neurosci. 2017, 11, 160. [Google Scholar] [CrossRef]
- Tsutajima, J.; Kunitake, T.; Wakazono, Y.; Takamiya, K. Selective injection system into hippocampus CA1 via monitored theta oscillation. PLoS ONE 2013, 8, e83129. [Google Scholar]
- Yap, B.W.; Sim, C.H. Comparisons of various types of normality tests. J. Stat. Comput. Simul. 2011, 81, 2141–2155. [Google Scholar] [CrossRef]
- Batool, S.; Raza, H.; Zaidi, J.; Riaz, S.; Hasan, S.; Syed, N.I. Synapse formation: From cellular and molecular mechanisms to neurodevelopmental and neurodegenerative disorders. J. Neurophysiol. 2019, 121, 1381–1397. [Google Scholar] [CrossRef]
- Crosson, S.M.; Dib, P.; Smith, J.K.; Zolotukhin, S. Helper-free Production of Laboratory Grade AAV and Purification by Iodixanol Density Gradient Centrifugation. Mol. Ther. Methods Clin. Dev. 2018, 10, 1–7. [Google Scholar] [CrossRef]
- Cullen, D.K.; Gilroy, M.E.; Irons, H.R.; LaPlaca, M.C. Synapse-to-neuron ratio is inversely related to neuronal density in mature neuronal cultures. Brain Res. 2010, 1359, 44–55. [Google Scholar] [CrossRef] [Green Version]
- Köhler, S.; Black, S.E.; Sinden, M.; Szekely, C.; Kidron, D.; Parker, J.L.; Foster, J.K.; Moscovitch, M.; Winocour, G.; Szalai, J.P.; et al. Memory impairments associated with hippocampal versus parahippocampal-gyrus atrophy: An MR volumetry study in Alzheimer’s disease. Neuropsychologia 1998, 36, 901–914. [Google Scholar] [CrossRef]
- Freund, R.K.; Graw, S.; Choo, K.S.; Stevens, K.E.; Leonard, S.; Dell’Acqua, M.L. Genetic knockout of the α7 nicotinic acetylcholine receptor gene alters hippocampal long-term potentiation in a background strain-dependent manner. Neurosci. Lett. 2016, 627, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez, C.M.; Cortez, I.; Gu, Z.; Colón-Sáez, J.O.; Lamb, P.W.; Wakamiya, M.; Yakel, J.L.; Dineley, K.T. Research tool: Validation of floxed α7 nicotinic acetylcholine receptor conditional knockout mice using in vitro and in vivo approaches. J. Physiol. 2014, 592, 3201–3214. [Google Scholar] [CrossRef] [PubMed]
- Morley, B.; Mervis, R. Dendritic spine alterations in the hippocampus and parietal cortex of alpha7 nicotinic acetylcholine receptor knockout mice. Neuroscience 2012, 233, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Parada, E.; Egea, J.; Buendia, I.; Negredo, P.; Cunha, A.C.; Cardoso, S.; Soares, M.P.; López, M.G. The microglial α7-acetylcholine nicotinic receptor is a key element in promoting neuroprotection by inducing heme oxygenase-1 via nuclear factor erythroid-2-related factor 2. Antioxidants Redox Signal. 2013, 19, 1135–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Challis, R.C.; Ravindra Kumar, S.; Chan, K.Y.; Challis, C.; Beadle, K.; Jang, M.J.; Kim, H.M.; Rajendran, P.S.; Tompkins, J.D.; Shivkumar, K.; et al. Systemic AAV vectors for widespread and targeted gene delivery in rodents. Nat. Protoc. 2019, 14, 379–414. [Google Scholar] [CrossRef]
- Anand, K.S.; Dhikav, V. Hippocampus in health and disease: An overview. Ann. Indian Acad. Neurol. 2012, 15, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Jarrard, L.E. On the role of the hippocampus in learning and memory in the rat. Behav. Neural Biol. 1993, 60, 9–26. [Google Scholar] [CrossRef]
- Shoji, H.; Takao, K.; Hattori, S.; Miyakawa, T. Contextual and Cued Fear Conditioning Test Using a Video Analyzing System in Mice. J. Vis. Exp. 2014, 85, e50871. [Google Scholar] [CrossRef] [Green Version]
- Choi, M.; Ahn, S.; Yang, E.J.; Kim, H.; Chong, Y.H.; Kim, H.-S. Hippocampus-based contextual memory alters the morphological characteristics of astrocytes in the dentate gyrus. Mol. Brain 2016, 9, 72. [Google Scholar] [CrossRef] [Green Version]
- Lovett-Barron, M.; Kaifosh, P.; Kheirbek, M.A.; Danielson, N.; Zaremba, J.D.; Reardon, T.R.; Turi, G.F.; Hen, R.; Zemelman, B.V.; Losonczy, A. Dendritic Inhibition in the Hippocampus Supports Fear Learning. Science 2014, 343, 857–863. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, J.C.; Su, K.; Goldberg, A.R.; Luna, V.M.; Biane, J.S.; Ordek, G.; Zhou, P.; Ong, S.K.; Wright, M.A.; Zweifel, L.; et al. Anxiety Cells in a Hippocampal-Hypothalamic Circuit. Neuron 2018, 97, 670–683.e676. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Ramirez, S.; Pang, P.T.; Puryear, C.B.; Govindarajan, A.; Deisseroth, K.; Tonegawa, S. Optogenetic stimulation of a hippocampal engram activates fear memory recall. Nature 2012, 484, 381–385. [Google Scholar] [CrossRef] [Green Version]
- Vorhees, C.V.; Williams, M.T. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, S.J.; Stackman, R.W., Jr. Assessing rodent hippocampal involvement in the novel object recognition task. A review. Behav. Brain Res. 2015, 285, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Van Asselen, M.; Kessels, R.P.; Neggers, S.F.; Kappelle, L.J.; Frijns, C.J.; Postma, A. Brain areas involved in spatial working memory. Neuropsychologia 2006, 44, 1185–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, J.; Maren, S. Differential roles for hippocampal areas CA1 and CA3 in the contextual encoding and retrieval of extinguished fear. Learn. Mem. 2008, 15, 244–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramaniyan, M.; Manivannan, S.; Chelur, V.; Tsetsenis, T.; Jiang, E.; Dani, J.A. Fear conditioning potentiates the hippocampal CA1 commissural pathway in vivo and increases awake phase sleep. Hippocampus 2021, 31, 1154–1175. [Google Scholar] [CrossRef]
- Takamiya, S.; Shiotani, K.; Ohnuki, T.; Osako, Y.; Tanisumi, Y.; Yuki, S.; Manabe, H.; Hirokawa, J.; Sakurai, Y. Hippocampal CA1 Neurons Represent Positive Feedback During the Learning Process of an Associative Memory Task. Front. Syst. Neurosci. 2021, 15, 718619. [Google Scholar] [CrossRef]
- Jimenez, J.C.; Berry, J.E.; Lim, S.C.; Ong, S.K.; Kheirbek, M.A.; Hen, R. Contextual fear memory retrieval by correlated ensembles of ventral CA1 neurons. Nat. Commun. 2020, 11, 3492. [Google Scholar] [CrossRef]
- Bali, Z.K.; Nagy, L.V.; Hernádi, I. Alpha7 nicotinic acetylcholine receptors play a predominant role in the cholinergic potentiation of N-Methyl-D-aspartate evoked firing responses of hippocampal CA1 pyramidal cells. Front. Cell. Neurosci. 2017, 11, 271. [Google Scholar] [CrossRef] [PubMed]
- Cortez, I.; Ishimwe, E.; Hernadex, C.; Dineley, K.T. The Role of Astrocytic α7 Nicotinic Acetylcholine Receptors in Neuroinflammation-Mediated Cognitive Impairment. Alzheimer’s Dement. 2019, 15, P542–P543. [Google Scholar] [CrossRef]
- Keith, D.; El-Husseini, A. Excitation control: Balancing PSD-95 function at the synapse. Front. Mol. Neurosci. 2008, 1, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Husseini, A.E.; Schnell, E.; Chetkovich, D.M.; Nicoll, R.A.; Bredt, D.S. Involvement in Maturation of Excitatory Synapses. Science 2000, 290, 1364–1368. [Google Scholar] [CrossRef] [PubMed]
- Neff, R.A., 3rd; Gomez-Varela, D.; Fernandes, C.C.; Berg, D.K. Postsynaptic scaffolds for nicotinic receptors on neurons. Acta Pharmacol. Sin. 2009, 30, 694–701. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Varela, D.; Schmidt, M.; Schoellerman, J.; Peters, E.C.; Berg, D.K. PMCA2 via PSD-95 Controls Calcium Signaling by 7-Containing Nicotinic Acetylcholine Receptors on Aspiny Interneurons. J. Neurosci. 2012, 32, 6894–6905. [Google Scholar] [CrossRef]
- Sultana, R.; Banks, W.A.; Butterfield, D.A. Decreased levels of PSD95 and two associated proteins and increased levels of BCl2 and caspase 3 in hippocampus from subjects with amnestic mild cognitive impairment: Insights into their potential roles for loss of synapses and memory, accumulation of Aβ, and neurodegeneration in a prodromal stage of Alzheimer’s disease. J. Neurosci. Res. 2010, 88, 469–477. [Google Scholar]
- Shao, C.Y.; Mirra, S.S.; Sait, H.B.; Sacktor, T.C.; Sigurdsson, E.M. Postsynaptic degeneration as revealed by PSD-95 reduction occurs after advanced Aβ and tau pathology in transgenic mouse models of Alzheimer’s disease. Acta Neuropathol. 2011, 122, 285–292. [Google Scholar] [CrossRef] [Green Version]
- Glavan, G.; Schliebs, R.; Zivin, M. Synaptotagmins in Neurodegeneration. Anat. Rec. 2009, 292, 1849–1862. [Google Scholar] [CrossRef]
- Gautam, V.; D’Avanzo, C.; Berezovska, O.; Tanzi, R.E.; Kovacs, D.M. Synaptotagmins interact with APP and promote Aβ generation. Mol. Neurodegener. 2015, 10, 31. [Google Scholar] [CrossRef] [Green Version]
- Moss Bendtsen, K.; Jensen, M.H.; Krishna, S.; Semsey, S. The role of mRNA and protein stability in the function of coupled positive and negative feedback systems in eukaryotic cells. Sci. Rep. 2015, 5, 13910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vernino, S.; Amador, M.; Luetje, C.W.; Patrick, J.; Dani, J.A. Calcium modulation and high calcium permeability of neuronal nicotinic acetylcholine receptors. Neuron 1992, 8, 127–134. [Google Scholar] [CrossRef]
- Colón-Sáez, J.O.; Yakel, J.L. A mutation in the extracellular domain of the α7 nAChR reduces calcium permeability. Pflügers Arch. 2014, 466, 1571–1579. [Google Scholar] [CrossRef] [Green Version]
- Getz, A.M.; Visser, F.; Bell, E.M.; Xu, F.; Flynn, N.M.; Zaidi, W.; Syed, N.I. Two proteolytic fragments of menin coordinate the nuclear transcription and postsynaptic clustering of neurotransmitter receptors during synaptogenesis between Lymnaea neurons. Sci. Rep. 2016, 6, 31779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matta, J.A.; Gu, S.; Davini, W.B.; Lord, B.; Siuda, E.R.; Harrington, A.W.; Bredt, D.S. NACHO Mediates Nicotinic Acetylcholine Receptor Function throughout the Brain. Cell Rep. 2017, 19, 688–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turrigiano, G.G. The Self-Tuning Neuron: Synaptic Scaling of Excitatory Synapses. Cell 2008, 135, 422–435. [Google Scholar] [CrossRef] [Green Version]
- Moss, B.J.; Park, L.; Dahlberg, C.L.; Juo, P. The CaM Kinase CMK-1 Mediates a Negative Feedback Mechanism Coupling the C. elegans Glutamate Receptor GLR-1 with Its Own Transcription. PLoS Genet. 2016, 12, e1006180. [Google Scholar] [CrossRef]
- Chowdhury, D.; Hell, J.W. Homeostatic synaptic scaling: Molecular regulators of synaptic AMPA-type glutamate receptors. F1000Research 2018, 7, 234. [Google Scholar] [CrossRef] [Green Version]
- Jin, D.-Z.; Guo, M.-L.; Xue, B.; Fibuch, E.E.; Choe, E.S.; Mao, L.-M.; Wang, J.Q. Phosphorylation and feedback regulation of metabotropic glutamate receptor 1 by calcium/calmodulin-dependent protein kinase II. J. Neurosci. 2013, 33, 3402–3412. [Google Scholar] [CrossRef] [Green Version]
- Hahm, E.T.; Nagaraja, R.Y.; Waro, G.; Tsunoda, S. Cholinergic Homeostatic Synaptic Plasticity Drives the Progression of Aβ-Induced Changes in Neural Activity. Cell Rep. 2018, 24, 342–354. [Google Scholar] [CrossRef] [Green Version]
- Eadaim, A.; Hahm, E.-T.; Justice, D.E.; Tsunoda, S. Cholinergic Synaptic Homeostasis Is Tuned by an NFAT-Mediated α7 nAChR-K v 4/Shal Coupled Regulatory System. Cell Rep. 2020, 32, 108119. [Google Scholar] [CrossRef]
- Berdugo-Vega, G.; Arias-Gil, G.; López-Fernández, A.; Artegiani, B.; Wasielewska, J.M.; Lee, C.-C.; Lippert, M.T.; Kempermann, G.; Takagaki, K.; Calegari, F. Increasing neurogenesis refines hippocampal activity rejuvenating navigational learning strategies and contextual memory throughout life. Nat. Commun. 2020, 11, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kempermann, G.; Song, H.; Gage, F.H. Neurogenesis in the adult hippocampus. Cold Spring Harb. Perspect. Biol. 2015, 7, a018812. [Google Scholar] [CrossRef] [Green Version]
- Parri, H.R.; Hernandez, C.M.; Dineley, K.T. Research update: Alpha7 nicotinic acetylcholine receptor mechanisms in Alzheimer’s disease. Biochem. Pharmacol. 2011, 82, 931–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Öhrfelt, A.; Brinkmalm, A.; Dumurgier, J.; Brinkmalm, G.; Hansson, O.; Zetterberg, H.; Bouaziz-Amar, E.; Hugon, J.; Paquet, C.; Blennow, K. The pre-synaptic vesicle protein synaptotagmin is a novel biomarker for Alzheimer’s disease. Alzheimer’s Res. Ther. 2016, 8, 41. [Google Scholar] [CrossRef] [Green Version]
- Caton, M.; Ochoa, E.; Barrantes, F.J. The role of nicotinic cholinergic neurotransmission in delusional thinking. NPJ Schizophr. 2020, 6, 16. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Genbank Accession Number | 97 Mer shRNA Sequence (siRNA Underlined) | Splash Score | Antisense. Guide. Sequence |
|---|---|---|---|---|
| CHRNA7 | NM_007390 | TGCTGTTGACAGTGAGCGAGACCAGGATCATTCTTCTGAATAGTGAAGCCACAGATGTATTCAGAAGAATGATCCTGGTCCTGCCTACTGCCTCGGA | 1.49 | TTCAGAAGAATGATCCTGGTCC |
| α7 scrambled | NM_007390 | TGCTGTTGACAGTGAGCGATCAACTATATGCGAGGTGACTTAGTGAAGCCACAGATGTAAGTCACCTCGCATATAGTTGAGTGCCTACTGCCTCGGA | 1.49 | AGTCACCTCGCATATAGTTGAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Batool, S.; Akhter, B.; Zaidi, J.; Visser, F.; Petrie, G.; Hill, M.; Syed, N.I. Neuronal Menin Overexpression Rescues Learning and Memory Phenotype in CA1-Specific α7 nAChRs KD Mice. Cells 2021, 10, 3286. https://doi.org/10.3390/cells10123286
Batool S, Akhter B, Zaidi J, Visser F, Petrie G, Hill M, Syed NI. Neuronal Menin Overexpression Rescues Learning and Memory Phenotype in CA1-Specific α7 nAChRs KD Mice. Cells. 2021; 10(12):3286. https://doi.org/10.3390/cells10123286
Chicago/Turabian StyleBatool, Shadab, Basma Akhter, Jawwad Zaidi, Frank Visser, Gavin Petrie, Matthew Hill, and Naweed I. Syed. 2021. "Neuronal Menin Overexpression Rescues Learning and Memory Phenotype in CA1-Specific α7 nAChRs KD Mice" Cells 10, no. 12: 3286. https://doi.org/10.3390/cells10123286
APA StyleBatool, S., Akhter, B., Zaidi, J., Visser, F., Petrie, G., Hill, M., & Syed, N. I. (2021). Neuronal Menin Overexpression Rescues Learning and Memory Phenotype in CA1-Specific α7 nAChRs KD Mice. Cells, 10(12), 3286. https://doi.org/10.3390/cells10123286