
The Methyltransferase Smyd1 Mediates LPS-Triggered Up-Regulation of IL-6 in Endothelial Cells

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cell Stimulation Experiments with LPS or IL-6
2.3. Treatment of Cells with PDTC or Bay 11-7082
2.4. Transfection with Human Expression Vectors
2.5. Transfection with Specific siRNAs
2.6. RNA Isolation, Reverse Transcription and Real Time qPCR
2.7. Immunoblotting
2.8. Immunofluorescence Analysis
2.9. ELISA Analysis
2.10. Luciferase Assay Reporter Gene Assay
2.11. Flow Cytometry
2.12. ChIP RT-qPCR Assays
2.13. Statistical Analysis
3. Results
3.1. LPS Stimulation of Endothelial Cells Leads to Higher Smyd1 Expression
3.2. Smyd1 Increases IL-6 Levels in Endothelial Cells
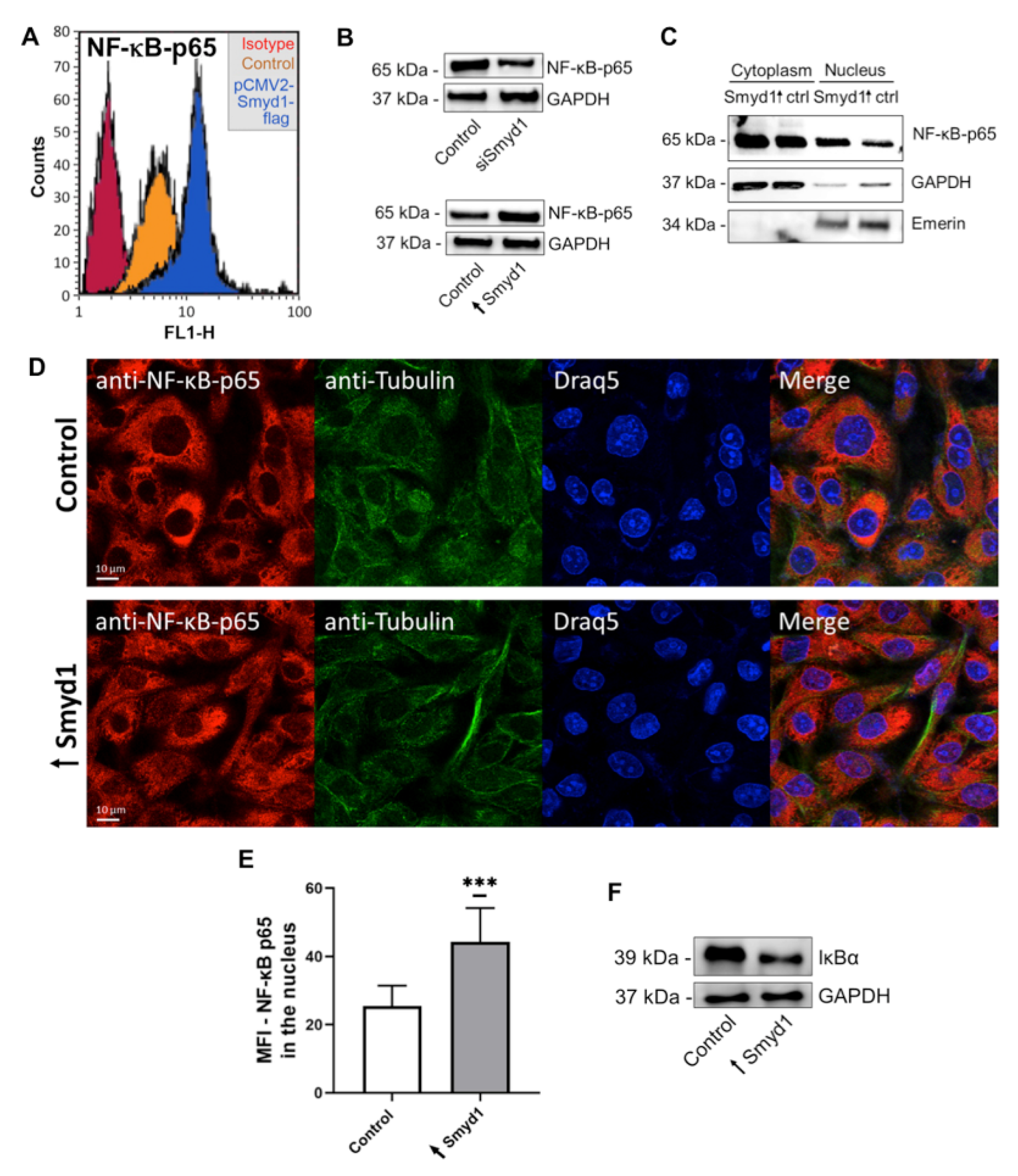
3.3. Smyd1 Induces IL-6 Expression via Activation of NF-κB
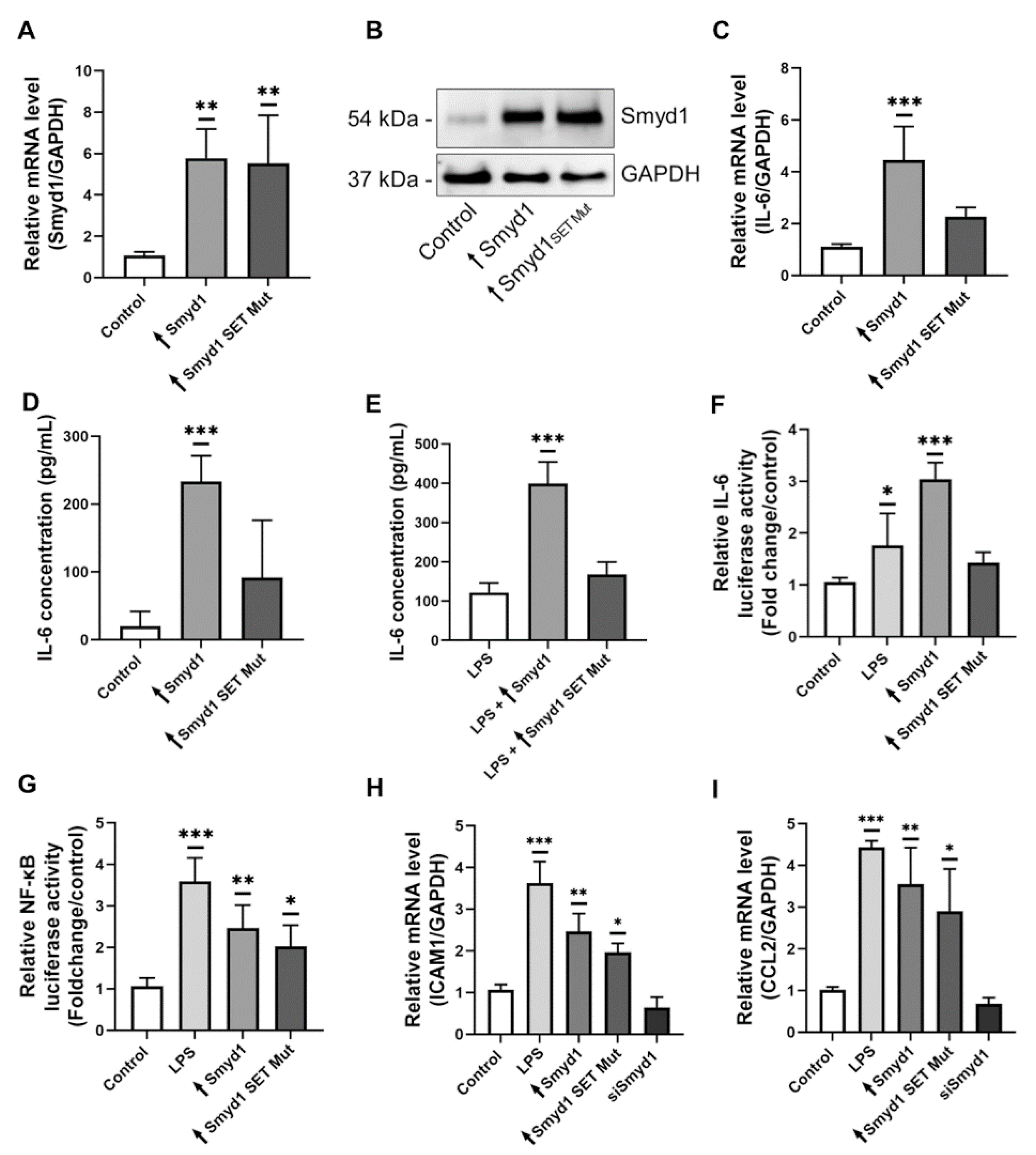
3.4. Smyd1 Induces IL-6 Expression also Independently of NF-κB
3.5. The Methyltransferase Activity of Smyd1 Is Involved in the Regulation of IL-6 Expression
3.6. Smyd1 Affects the H3K4me3 Methylation Pattern of the IL-6 Promoter
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qian, C.; Zhou, M.-M. SET domain protein lysine methyltransferases: Structure, specificity and catalysis. Cell Mol. Life Sci. 2006, 63, 2755–2763. [Google Scholar] [CrossRef]
- Dillon, S.C.; Zhang, X.; Trievel, R.C.; Cheng, X. The SET-domain protein superfamily: Protein lysine methyltransferases. Genome Biol. 2005, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Spellmon, N.; Holcomb, J.; Trescott, L.; Sirinupong, N.; Yang, Z. Structure and function of SET and MYND domain-containing proteins. Int. J. Mol. Sci. 2015, 16, 1406–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tracy, C.M.; Warren, J.S.; Szulik, M.; Wang, L.; Garcia, J.; Makaju, A.; Russell, K.; Miller, M.; Franklin, S. The Smyd family of methyltransferases: Role in cardiac and skeletal muscle physiology and pathology. Curr. Opin. Physiol. 2018, 1, 140–152. [Google Scholar] [CrossRef]
- Huang, L.; Xu, A.-M. SET and MYND domain containing protein 3 in cancer. Am. J. Transl. Res. 2017, 9, 1. [Google Scholar]
- Brown, M.A.; Sims, R.J.; Gottlieb, P.D.; Tucker, P.W. Identification and characterization of Smyd2: A split SET/MYND domain-containing histone H3 lysine 36-specific methyltransferase that interacts with the Sin3 histone deacetylase complex. Mol. Cancer 2006, 5, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bannister, A.J.; Kouzarides, T. Regulation of chromatin by histone modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef]
- Strahl, B.D.; Allis, C.D. The language of covalent histone modifications. Nature 2000, 403, 41–45. [Google Scholar] [CrossRef]
- Sims, R.J.; Weihe, E.K.; Zhu, L.; O’Malley, S.; Harriss, J.V.; Gottlieb, P.D. m-Bop, a repressor protein essential for cardiogenesis, interacts with skNAC, a heart-and muscle-specific transcription factor. J. Biol. Chem. 2002, 277, 26524–26529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, T.L.; Ma, Y.; Park, C.Y.; Harriss, J.; Pierce, S.A.; Dekker, J.D.; Valenzuela, N.; Srivastava, D.; Schwartz, R.J.; Stewart, M.D. Smyd1 facilitates heart development by antagonizing oxidative and ER stress responses. PLoS ONE 2015, 10, e0121765. [Google Scholar] [CrossRef] [Green Version]
- Franklin, S.; Kimball, T.; Rasmussen, T.L.; Rosa-Garrido, M.; Chen, H.; Tran, T.; Miller, M.R.; Gray, R.; Jiang, S.; Ren, S. The chromatin-binding protein Smyd1 restricts adult mammalian heart growth. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H1234–H1247. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Wu, J.; Sun, B.; Zhong, C.; Ding, J. Structural and biochemical studies of human lysine methyltransferase Smyd3 reveal the important functional roles of its post-SET and TPR domains and the regulation of its activity by DNA binding. Nucleic Acids Res. 2011, 39, 4438–4449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stender, J.D.; Pascual, G.; Liu, W.; Kaikkonen, M.U.; Do, K.; Spann, N.J.; Boutros, M.; Perrimon, N.; Rosenfeld, M.G.; Glass, C.K. Control of proinflammatory gene programs by regulated trimethylation and demethylation of histone H4K20. Mol. Cell 2012, 48, 28–38. [Google Scholar] [CrossRef] [Green Version]
- Thompson, E.C.; Travers, A.A. A Drosophila Smyd4 homologue is a muscle-specific transcriptional modulator involved in development. PLoS ONE 2008, 3, e3008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirinupong, N.; Brunzelle, J.; Ye, J.; Pirzada, A.; Nico, L.; Yang, Z. Crystal structure of cardiac-specific histone methyltransferase SmyD1 reveals unusual active site architecture. J. Biol. Chem. 2010, 285, 40635–40644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, X.; Qian, Y.; Wang, Q.; Yuan, W.; Mo, X.; Li, Y.; Jiang, Z.; Xu, W.; Deng, Y.; Wan, Y. SMYD1, an SRF-interacting partner, is involved in angiogenesis. PLoS ONE 2016, 11, e0146468. [Google Scholar]
- Becker, S.; Steinemann, G.; Karle, W.; Roos, K.; Liem, C.H.; Muralikumar, S.; Volkamer, A.; Munz, B.; Zakrzewicz, A.; Berkholz, J. Stability of Smyd1 in endothelial cells is controlled by PML-dependent SUMOylation upon cytokine stimulation. Biochem. J. 2021, 478, 217–234. [Google Scholar] [CrossRef]
- Medzhitov, R. Toll-like receptors and innate immunity. Nature Rev. Immunol. 2001, 1, 135–145. [Google Scholar] [CrossRef]
- Lush, C.W.; Cepinskas, G.; Kvietys, P.R. LPS tolerance in human endothelial cells: Reduced PMN adhesion, E-selectin expression, and NF-κB mobilization. Am. J. Physiol. Heart Circ. Physiol. 2000, 278, H853–H861. [Google Scholar] [CrossRef]
- Wong, D.; Dorovini-Zis, K. Upregulation of intercellular adhesion molecule-1 (ICAM-1) expression in primary cultures of human brain microvessel endothelial cells by cytokines and lipopolysaccharide. J. Neuroimmunol. 1992, 39, 11–21. [Google Scholar] [CrossRef]
- Bozza, F.A.; Salluh, J.I.; Japiassu, A.M.; Soares, M.; Assis, E.F.; Gomes, R.N.; Bozza, M.T.; Castro-Faria-Neto, H.C.; Bozza, P.T. Cytokine profiles as markers of disease severity in sepsis: A multiplex analysis. Crit. Care 2007, 11, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jirik, F.; Podor, T.; Hirano, T.; Kishimoto, T.; Loskutoff, D.; Carson, D.; Lotz, M. Bacterial lipopolysaccharide and inflammatory mediators augment IL-6 secretion by human endothelial cells. J. Immunol. 1989, 142, 144–147. [Google Scholar]
- Bannerman, D.D.; Sathyamoorthy, M.; Goldblum, S.E. Bacterial lipopolysaccharide disrupts endothelial monolayer integrity and survival signaling events through caspase cleavage of adherens junction proteins. J. Biol. Chem. 1998, 273, 35371–35380. [Google Scholar] [CrossRef] [Green Version]
- Hotchkiss, R.S.; Tinsley, K.W.; Swanson, P.E.; Karl, I.E. Endothelial cell apoptosis in sepsis. Crit. Care Med. 2002, 30, S225–S228. [Google Scholar] [CrossRef] [PubMed]
- Pawlinski, R.; Mackman, N. Tissue factor, coagulation proteases, and protease-activated receptors in endotoxemia and sepsis. Crit. Care Med. 2004, 32, S293–S297. [Google Scholar] [CrossRef]
- Pinsky, M.R. Dysregulation of the immune response in severe sepsis. Am. J. Med Sci. 2004, 328, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Volk, T.; Kox, W. Endothelium function in sepsis. Inflamm. Res. 2000, 49, 185–198. [Google Scholar] [CrossRef]
- Maroski, J.; Vorderwülbecke, B.J.; Fiedorowicz, K.; Da Silva-Azevedo, L.; Siegel, G.; Marki, A.; Pries, A.R.; Zakrzewicz, A. Shear stress increases endothelial hyaluronan synthase 2 and hyaluronan synthesis especially in regard to an atheroprotective flow profile. Exp. Physiol. 2011, 96, 977–986. [Google Scholar] [CrossRef]
- Berkholz, J.; Kuzyniak, W.; Hoepfner, M.; Munz, B. Overexpression of the skNAC gene in human rhabdomyosarcoma cells enhances their differentiation potential and inhibits tumor cell growth and spreading. Clin. Exp. Metastasis. 2014, 31, 869–879. [Google Scholar] [CrossRef] [PubMed]
- Alspach, E.; Flanagan, K.C.; Luo, X.; Ruhland, M.K.; Huang, H.; Pazolli, E.; Donlin, M.J.; Marsh, T.; Piwnica-Worms, D.; Monahan, J. p38MAPK plays a crucial role in stromal-mediated tumorigenesis. Cancer. Discov. 2014, 4, 716–729. [Google Scholar] [CrossRef] [Green Version]
- Dandrea, M.; Donadelli, M.; Costanzo, C.; Scarpa, A.; Palmieri, M. MeCP2/H3meK9 are involved in IL-6 gene silencing in pancreatic adenocarcinoma cell lines. Nucleic Acids Res. 2009, 37, 6681–6690. [Google Scholar] [CrossRef] [Green Version]
- Poplutz, M.K.; Wessels, I.; Rink, L.; Uciechowski, P. Regulation of the Interleukin-6 gene expression during monocytic differentiation of HL-60 cells by chromatin remodeling and methylation. Immunobiology 2014, 219, 619–626. [Google Scholar] [CrossRef]
- Yang, F.; Zhou, S.; Wang, C.; Huang, Y.; Li, H.; Wang, Y.; Zhu, Z.; Tang, J.; Yan, M. Epigenetic modifications of interleukin-6 in synovial fibroblasts from osteoarthritis patients. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, T.L.; Tucker, H.O. Loss of SMYD1 Results in Perinatal Lethality via Selective Defects within Myotonic Muscle Descendants. Diseases 2019, 7, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottlieb, P.D.; Pierce, S.A.; Sims, R.J.; Yamagishi, H.; Weihe, E.K.; Harriss, J.V.; Maika, S.D.; Kuziel, W.A.; King, H.L.; Olson, E.N. Bop encodes a muscle-restricted protein containing MYND and SET domains and is essential for cardiac differentiation and morphogenesis. Nat. Genet. 2002, 31, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Rotllant, J.; Li, H.; DeDeyne, P.; Du, S.J. SmyD1, a histone methyltransferase, is required for myofibril organization and muscle contraction in zebrafish embryos. Proc. Nat. Acad. Sci. USA 2006, 103, 2713–2718. [Google Scholar] [CrossRef] [Green Version]
- Berkholz, J.; Orgeur, M.; Stricker, S.; Munz, B. skNAC and Smyd1 in transcriptional control. Exp. Cell Res. 2015, 336, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Schwartz, R.J.; Liu, J.; Sun, F.; Li, Q.; Ma, Y. Smyd1 Orchestrates Early Heart Development Through Positive and Negative Gene Regulation. Front. Cell Dev. Biol. 2021, 9, 500. [Google Scholar] [CrossRef] [PubMed]
- Warren, J.S.; Tracy, C.M.; Miller, M.R.; Makaju, A.; Szulik, M.W.; Oka, S.-i.; Yuzyuk, T.N.; Cox, J.E.; Kumar, A.; Lozier, B.K. Histone methyltransferase Smyd1 regulates mitochondrial energetics in the heart. Proc. Nat. Acad. Sci. USA 2018, 115, E7871–E7880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, N.R.; Klose, R.J. Understanding the relationship between DNA methylation and histone lysine methylation. Biochim. Biophys. Acta 2014, 1839, 1362–1372. [Google Scholar] [CrossRef] [Green Version]
- Thomson, J.P.; Skene, P.J.; Selfridge, J.; Clouaire, T.; Guy, J.; Webb, S.; Kerr, A.R.; Deaton, A.; Andrews, R.; James, K.D. CpG islands influence chromatin structure via the CpG-binding protein Cfp1. Nature 2010, 464, 1082–1086. [Google Scholar] [CrossRef] [Green Version]
- Porter, N.J.; Christianson, D.W. Structure, mechanism, and inhibition of the zinc-dependent histone deacetylases. Curr. Opin. Struct. Biol. 2019, 59, 9. [Google Scholar] [CrossRef]
- Brasier, A.R. The nuclear factor-κB–interleukin-6 signalling pathway mediating vascular inflammation. Cardiovasc. Res. 2010, 86, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Pålsson-McDermott, E.M.; O’Neill, L.A. Signal transduction by the lipopolysaccharide receptor, Toll-like receptor-4. Immunology 2004, 113, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Dauphinee, S.M.; Karsan, A. Lipopolysaccharide signaling in endothelial cells. Lab. Investig. 2006, 86, 9–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Hagler, J.; Palombella, V.J.; Melandri, F.; Scherer, D.; Ballard, D.; Maniatis, T. Signal-induced site-specific phosphorylation targets I kappa B alpha to the ubiquitin-proteasome pathway. Genes. Dev. 1995, 9, 1586–1597. [Google Scholar] [CrossRef] [Green Version]
- Brown, K.; Park, S.; Kanno, T.; Franzoso, G.; Siebenlist, U. Mutual regulation of the transcriptional activator NF-kappa B and its inhibitor, I kappa B-alpha. Proc. Nat. Acad. Sci. USA 1993, 90, 2532–2536. [Google Scholar] [CrossRef] [Green Version]
- Balamurugan, K.; Mendoza-Villanueva, D.; Sharan, S.; Summers, G.H.; Dobrolecki, L.E.; Lewis, M.T.; Sterneck, E. C/EBPδ links IL-6 and HIF-1 signaling to promote breast cancer stem cell-associated phenotypes. Oncogene 2019, 38, 3765–3780. [Google Scholar] [CrossRef] [PubMed]
- Hungness, E.S.; Luo, G.j.; Pritts, T.A.; Sun, X.; Robb, B.W.; Hershko, D.; Hasselgren, P.O. Transcription factors C/EBP-β and-δ regulate IL-6 production in IL-1β-stimulated human enterocytes. J. Cell Physiol. 2002, 192, 64–70. [Google Scholar] [CrossRef]
- Persson, E.; Voznesensky, O.S.; Huang, Y.-F.; Lerner, U.H. Increased expression of interleukin-6 by vasoactive intestinal peptide is associated with regulation of CREB, AP-1 and C/EBP, but not NF-κB, in mouse calvarial osteoblasts. Bone 2005, 37, 513–529. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, J.M.; Glenn, J.D.; Calabresi, P.A.; Whartenby, K.A. KLF4 modulates expression of IL-6 in dendritic cells via both promoter activation and epigenetic modification. J. Biol. Chem. 2013, 288, 23868–23874. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.; Moon, J.W.; Choi, J.-H.; Lee, Y.W.; Park, S.-H.; Kim, G.J. Epigenetic alterations of IL-6/STAT3 signaling by placental stem cells promote hepatic regeneration in a rat model with CCl4-induced liver injury. Int. J. Stem. Cells 2015, 8, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.; Kim, J.; Yun, J.-M.; Ohn, T.; Gong, Q. MeCP2 regulates gene expression through recognition of H3K27me3. Nat. Commun. 2020, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Yu, Y.; Huang, H.; Fan, H.; Hu, L.; Yin, C.; Li, K.; Fulton, D.J.; Chen, F. Epigenetic regulation of interleukin 6 by histone acetylation in macrophages and its role in paraquat-induced pulmonary fibrosis. Front. Immunol. 2017, 7, 696. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Liu, G.; Xiong, S.; Liu, H.; Chen, X.; Zheng, B. The histone methyltransferase Smyd2 is a negative regulator of macrophage activation by suppressing interleukin 6 (IL-6) and tumor necrosis factor α (TNF-α) production. J. Biol. Chem. 2015, 290, 5414–5423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alkhatib, S.G.; Landry, J.W. The nucleosome remodeling factor. FEBS Lett. 2011, 585, 3197–3207. [Google Scholar] [CrossRef] [Green Version]
- Wysocka, J.; Swigut, T.; Xiao, H.; Milne, T.A.; Kwon, S.Y.; Landry, J.; Kauer, M.; Tackett, A.J.; Chait, B.T.; Badenhorst, P. A PHD finger of NURF couples histone H3 lysine 4 trimethylation with chromatin remodelling. Nature 2006, 442, 86–90. [Google Scholar] [CrossRef]
- Santos-Rosa, H.; Schneider, R.; Bernstein, B.E.; Karabetsou, N.; Morillon, A.; Weise, C.; Schreiber, S.L.; Mellor, J.; Kouzarides, T. Methylation of histone H3 K4 mediates association of the Isw1p ATPase with chromatin. Mol. Cell 2003, 12, 1325–1332. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Template | Forward Primer | Reverse Primer | Product Size | Annealing Temperature |
|---|---|---|---|---|
| GAPDH | 5′-ATG ACC TTG CCC ACA GCC TT-3′ | 5′-AAC TGC TTA GCA CCC CTG GC-3′ | 200 bp | 60 °C |
| IL-6 | 5′-TGC CAG CCT GCT GAC GAA G-3′ | 5′-AGC TGC GCA GAA TGA GAT GAG-3′ | 90 bp | 56 °C |
| Smyd1 | 5′-CTG GAG AAG CAG GAG CCA GTG TT-3′ | 5′-GCA TAG GCT TTG CAG ATC ATC CC-3′ | 257 bp | 60 °C |
| ICAM1 | 5′-GGC CGG CCA GCT TAT ACA C-3′ | 5′-TAG ACA CTT GAG CTC GGG CA-3′ | 166 bp | 58 °C |
| VCAM1 | 5′- TCA GAT TGG AGA CTC AGT CAT GT-3′ | 5′-ACT CCT CAC CTT CCC GCTC-3′ | 109 bp | 62 °C |
| CCL2 | 5′-GAG AGG CTG AGA CTA ACC CAG A-3′ | 5′-ATC ACA GCT TCT TTG GGA CAC T -3′ | 259 bp | 62 °C |
| Template | Forward Primer | Reverse Primer | Product Size | Position (for/rev) |
|---|---|---|---|---|
| IL-6 region 1 | 5′-TTT TCA CAC CAA AGA ATC CC-3′ | 5′-CTT ATT TAC CAA ACA TGG TGT-3′ | 231 bp | 3532/3763 |
| IL-6 region 2 | 5′-CAG GTG AAG AAA GTG GCA GA-3 | 5′-GAC CAG ATT AAC AGG CTA GAA-3′ | 292 bp | 4363/4655 |
| IL-6 region 3 | 5′-TCC TTA GCC CTG GAA CTG CC-3′ | 5′-AGG CAA CAC CAG GAG CAG CCC C-3′ | 197 bp | 5150/5347 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shamloul, A.; Steinemann, G.; Roos, K.; Liem, C.H.; Bernd, J.; Braun, T.; Zakrzewicz, A.; Berkholz, J. The Methyltransferase Smyd1 Mediates LPS-Triggered Up-Regulation of IL-6 in Endothelial Cells. Cells 2021, 10, 3515. https://doi.org/10.3390/cells10123515
Shamloul A, Steinemann G, Roos K, Liem CH, Bernd J, Braun T, Zakrzewicz A, Berkholz J. The Methyltransferase Smyd1 Mediates LPS-Triggered Up-Regulation of IL-6 in Endothelial Cells. Cells. 2021; 10(12):3515. https://doi.org/10.3390/cells10123515
Chicago/Turabian StyleShamloul, Ahmed, Gustav Steinemann, Kerrin Roos, Celine Huajia Liem, Jonathan Bernd, Thorsten Braun, Andreas Zakrzewicz, and Janine Berkholz. 2021. "The Methyltransferase Smyd1 Mediates LPS-Triggered Up-Regulation of IL-6 in Endothelial Cells" Cells 10, no. 12: 3515. https://doi.org/10.3390/cells10123515
APA StyleShamloul, A., Steinemann, G., Roos, K., Liem, C. H., Bernd, J., Braun, T., Zakrzewicz, A., & Berkholz, J. (2021). The Methyltransferase Smyd1 Mediates LPS-Triggered Up-Regulation of IL-6 in Endothelial Cells. Cells, 10(12), 3515. https://doi.org/10.3390/cells10123515