
Do Plasmodesmata Play a Prominent Role in Regulation of Auxin-Dependent Genes at Early Stages of Embryogenesis?



{kind=link}
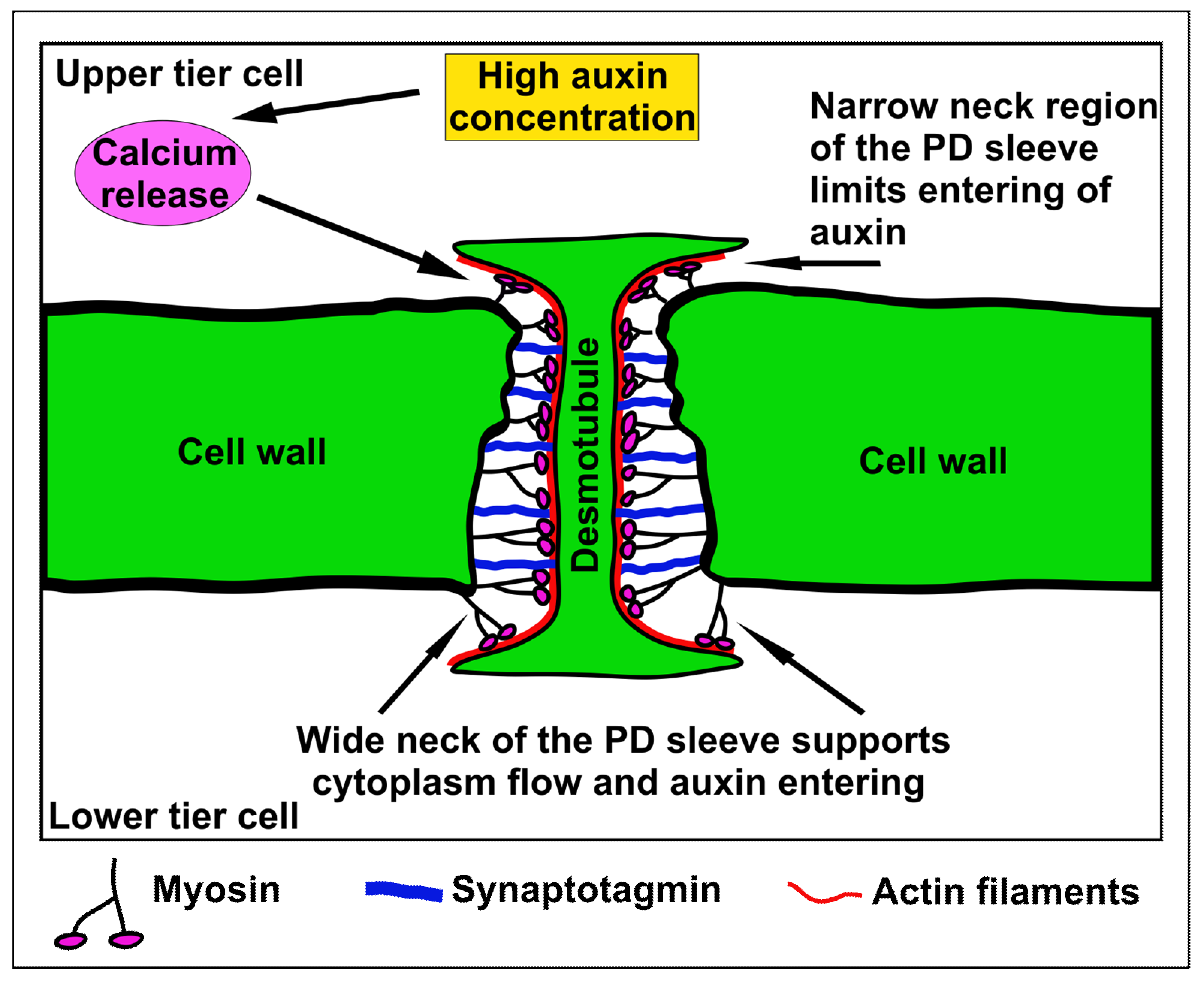{kind=link}
Abstract
:1. Introduction
2. Multi-Level Regulation of HD-ZIP Ⅲ and AP2-Domain Transcription Factors
3. The Role of Plasmodesmata during Developmental Processes
4. Do Callose-Dependent Changes of PD Permeability Regulate Transient Auxin Gradients?
5. Auxin May Regulate the Function of PD in a Callose-Independent Manner
6. Unidirectional Transport through PD—Science or Fiction
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jenik, P.D.; Barton, M.K. Surge and destroy: The role of auxin in plant embryogenesis. Development 2005, 132, 3577–3585. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.J.; Park, J.W.; Lee, H.W.; Kim, J. Genome-wide analysis of the auxin-responsive transcriptome downstream of iaa1 and its expression analysis reveal the diversity and complexity of auxin-regulated gene expression. J. Exp. Bot. 2009, 60, 3935–3957. [Google Scholar] [CrossRef] [Green Version]
- Paponov, I.A.; Paponov, M.; Teale, W.; Menges, M.; Chakrabortee, S.; Murray, J.A.; Palme, K. Comprehensive transcriptome analysis of auxin responses in Arabidopsis. Mol. Plant 2008, 1, 321–337. [Google Scholar] [CrossRef]
- Mohanta, T.K.; Bashir, T.; Hashem, A.; Abd_Allah, E.F.; Khan, A.L.; Al-Harrasi, A.S. Molecular players of auxin transport systems: Advances in genomic and molecular events. J. Plant. Interact. 2018, 13, 483–495. [Google Scholar] [CrossRef] [Green Version]
- Zazimalova, E.; Murphy, A.S.; Yang, H.; Hoyerova, K.; Hosek, P. Auxin transporters—why so many? Cold Spring Harb Perspect Biol. 2010, 2, a001552. [Google Scholar] [CrossRef] [Green Version]
- Cole, M.; Chandler, J.; Weijers, D.; Jacobs, B.; Comelli, P.; Werr, W. DORNROSCHEN is a direct target of the auxin response factor MONOPTEROS in the Arabidopsis embryo. Development 2009, 136, 1643–1651. [Google Scholar] [CrossRef] [Green Version]
- Wabnik, K.; Robert, H.S.; Smith, R.S.; Friml, J. Modeling framework for the establishment of the apical-basal embryonic axis in plants. Curr. Biol. 2013, 23, 2513–2518. [Google Scholar] [CrossRef]
- Friml, J.; Vieten, A.; Sauer, M.; Weijers, D.; Schwarz, H.; Hamann, T.; Offringa, R.; Jurgens, G. Efflux-dependent auxin gradients establish the apical-basal axis of Arabidopsis. Nature 2003, 426, 147–153. [Google Scholar] [CrossRef]
- Armengot, L.; Marques-Bueno, M.M.; Jaillais, Y. Regulation of polar auxin transport by protein and lipid kinases. J. Exp. Bot. 2016, 67, 4015–4037. [Google Scholar] [CrossRef] [Green Version]
- Willige, B.C.; Chory, J. A current perspective on the role of AGCVIII kinases in PIN-mediated apical hook development. Front. Plant. Sci. 2015, 6, 767. [Google Scholar] [CrossRef] [Green Version]
- Adamowski, M.; Friml, J. PIN-dependent auxin transport: Action, regulation, and evolution. Plant Cell 2015, 27, 20–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moller, B.; Weijers, D. Auxin control of embryo patterning. Cold Spring Harb Perspect Biol. 2009, 1, a001545. [Google Scholar] [CrossRef] [Green Version]
- Robert, H.S.; Grunewald, W.; Sauer, M.; Cannoot, B.; Soriano, M.; Swarup, R.; Weijers, D.; Bennett, M.; Boutilier, K.; Friml, J. Plant embryogenesis requires AUX/LAX-mediated auxin influx. Development 2015, 142, 702–711. [Google Scholar] [CrossRef] [Green Version]
- Robert, H.S.; Friml, J. Auxin and other signals on the move in plants. Nat. Chem. Biol. 2009, 5, 325–332. [Google Scholar] [CrossRef]
- Baba, A.I.; Valkai, I.; Labhane, N.M.; Koczka, L.; Andrasi, N.; Klement, E.; Darula, Z.; Medzihradszky, K.F.; Szabados, L.; Feher, A.; et al. CRK5 Protein Kinase Contributes to the Progression of Embryogenesis of Arabidopsis thaliana. Int. J. Mol. Sci. 2019, 20, 6120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slane, D.; Kong, J.; Berendzen, K.W.; Kilian, J.; Henschen, A.; Kolb, M.; Schmid, M.; Harter, K.; Mayer, U.; De Smet, I.; et al. Cell type-specific transcriptome analysis in the early Arabidopsis thaliana embryo. Development 2014, 141, 4831–4840. [Google Scholar] [CrossRef] [Green Version]
- Zhao, P.; Shi, D.-Q.; Yang, W.-C. Patterning the embryo in higher plants: Emerging pathways and challenges. Front. Biol. 2011, 6, 3–11. [Google Scholar] [CrossRef]
- Zhang, Z.; Laux, T. The asymmetric division of the Arabidopsis zygote: From cell polarity to an embryo axis. Sex. Plant Reprod 2011, 24, 161–169. [Google Scholar] [CrossRef]
- Emery, J.F.; Floyd, S.K.; Alvarez, J.; Eshed, Y.; Hawker, N.P.; Izhaki, A.; Baum, S.F.; Bowman, J.L. Radial patterning of Arabidopsis shoots by class III HD-ZIP and KANADI genes. Curr. Biol. 2003, 13, 1768–1774. [Google Scholar] [CrossRef] [Green Version]
- Smith, Z.R.; Long, J.A. Control of Arabidopsis apical-basal embryo polarity by antagonistic transcription factors. Nature 2010, 464, 423–426. [Google Scholar] [CrossRef] [Green Version]
- Aida, M.; Beis, D.; Heidstra, R.; Willemsen, V.; Blilou, I.; Galinha, C.; Nussaume, L.; Noh, Y.S.; Amasino, R.; Scheres, B. The PLETHORA genes mediate patterning of the Arabidopsis root stem cell niche. Cell 2004, 119, 109–120. [Google Scholar] [CrossRef] [Green Version]
- Blilou, I.; Xu, J.; Wildwater, M.; Willemsen, V.; Paponov, I.; Friml, J.; Heidstra, R.; Aida, M.; Palme, K.; Scheres, B. The PIN auxin efflux facilitator network controls growth and patterning in Arabidopsis roots. Nature 2005, 433, 39–44. [Google Scholar] [CrossRef]
- Miyashima, S.; Honda, M.; Hashimoto, K.; Tatematsu, K.; Hashimoto, T.; Sato-Nara, K.; Okada, K.; Nakajima, K. A comprehensive expression analysis of the Arabidopsis MICRORNA165/6 gene family during embryogenesis reveals a conserved role in meristem specification and a non-cell-autonomous function. Plant Cell Physiol. 2013, 54, 375–384. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Tucker, E.; Hermann, M.; Laux, T. A Molecular Framework for the Embryonic Initiation of Shoot Meristem Stem Cells. Dev. Cell 2017, 40, 264–277 e264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, A.; Yamamoto, K.T. Overexpression of the non-canonical Aux/IAA genes causes auxin-related aberrant phenotypes in Arabidopsis. Physiol. Plant 2008, 133, 397–405. [Google Scholar] [CrossRef]
- Muller, C.J.; Valdes, A.E.; Wang, G.; Ramachandran, P.; Beste, L.; Uddenberg, D.; Carlsbecker, A. PHABULOSA Mediates an Auxin Signaling Loop to Regulate Vascular Patterning in Arabidopsis. Plant Physiol. 2016, 170, 956–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, B.; Yu, Q.; Liu, J.; Wen, X.; Yan, Z.; Hu, K.; Li, H.; Kong, X.; Li, C.; Tian, H.; et al. Non-canonical AUX/IAA protein IAA33 competes with canonical AUX/IAA repressor IAA5 to negatively regulate auxin signaling. EMBO J. 2020, 39, e101515. [Google Scholar] [CrossRef]
- Tan, S.; Luschnig, C.; Friml, J. Pho-view of Auxin: Reversible Protein Phosphorylation in Auxin Biosynthesis, Transport and Signaling. Mol. Plant 2021, 14, 151–165. [Google Scholar] [CrossRef] [PubMed]
- Rademacher, E.H.; Lokerse, A.S.; Schlereth, A.; Llavata-Peris, C.I.; Bayer, M.; Kientz, M.; Freire Rios, A.; Borst, J.W.; Lukowitz, W.; Jurgens, G.; et al. Different auxin response machineries control distinct cell fates in the early plant embryo. Dev. Cell 2012, 22, 211–222. [Google Scholar] [CrossRef] [Green Version]
- Llavata-Peris, C.I. Proteomic and mechanistic analysis of Auxin Response Factors in the Arabidopsis embryo. PhD Thesis, Wageningen University, Wageningen, The Netherlands, 2013. [Google Scholar]
- Gu, X.L.; Wang, H.; Huang, H.; Cui, X.F. SPT6L encoding a putative WG/GW-repeat protein regulates apical-basal polarity of embryo in Arabidopsis. Mol. Plant 2012, 5, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Roberts, A.G.; Oparka, K.J. Plasmodesmata and the control of symplastic transport. Plant Cell Environ. 2003, 26, 103–124. [Google Scholar] [CrossRef] [Green Version]
- White, R.G.; Barton, D.A. The cytoskeleton in plasmodesmata: A role in intercellular transport? J. Exp. Bot. 2011, 62, 5249–5266. [Google Scholar] [CrossRef] [Green Version]
- Kitagawa, M.; Jackson, D. Plasmodesmata-Mediated Cell-to-Cell Communication in the Shoot Apical Meristem: How Stem Cells Talk. Plants 2017, 6, 12. [Google Scholar] [CrossRef]
- Sun, Y.; Huang, D.; Chen, X. Dynamic regulation of plasmodesmatal permeability and its application to horticultural research. Hortic. Res. 2019, 6, 47. [Google Scholar] [CrossRef] [Green Version]
- Sager, R.E.; Lee, J.Y. Plasmodesmata at a glance. J. Cell Sci. 2018, 131. [Google Scholar] [CrossRef] [Green Version]
- Faulkner, C. Plasmodesmata and the symplast. Curr. Biol. 2018, 28, R1374–R1378. [Google Scholar] [CrossRef] [Green Version]
- Sager, R.; Lee, J.Y. Plasmodesmata in integrated cell signalling: Insights from development and environmental signals and stresses. J. Exp. Bot. 2014, 65, 6337–6358. [Google Scholar] [CrossRef] [Green Version]
- Schulz, R.; Jensen, W.A. Capsella embryogenesis: The egg, zygote, and young embryo. Am. J. Bot. 1968, 55, 807–819. [Google Scholar] [CrossRef]
- Schlereth, A.; Moller, B.; Liu, W.; Kientz, M.; Flipse, J.; Rademacher, E.H.; Schmid, M.; Jurgens, G.; Weijers, D. MONOPTEROS controls embryonic root initiation by regulating a mobile transcription factor. Nature 2010, 464, 913–916. [Google Scholar] [CrossRef]
- Llavata-Peris, C.; Lokerse, A.; Moller, B.; De Rybel, B.; Weijers, D. Imaging of phenotypes, gene expression, and protein localization during embryonic root formation in Arabidopsis. Methods Mol. Biol 2013, 959, 137–148. [Google Scholar] [CrossRef]
- Kim, I.; Zambryski, P.C. Cell-to-cell communication via plasmodesmata during Arabidopsis embryogenesis. Curr. Opin. Plant Biol. 2005, 8, 593–599. [Google Scholar] [CrossRef]
- Kim, I.; Kobayashi, K.; Cho, E.; Zambryski, P.C. Subdomains for transport via plasmodesmata corresponding to the apical–basal axis are established during Arabidopsis embryogenesis. PNAS 2005, 102, 11945–11950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stadler, R.; Lauterbach, C.; Sauer, N. Cell-to-cell movement of green fluorescent protein reveals post-phloem transport in the outer integument and identifies symplastic domains in Arabidopsis seeds and embryos. Plant Physiol. 2005, 139, 701–712. [Google Scholar] [CrossRef] [Green Version]
- Saatian, B.; Austin, R.S.; Tian, G.; Chen, C.; Nguyen, V.; Kohalmi, S.E.; Geelen, D.; Cui, Y. Analysis of a novel mutant allele of GSL8 reveals its key roles in cytokinesis and symplastic trafficking in Arabidopsis. BMC Plant Biol. 2018, 18, 295. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.Y.; Liu, L.; Lee, E.; Han, X.; Rim, Y.; Chu, H.; Kim, S.W.; Sack, F.; Kim, J.Y. The Arabidopsis callose synthase gene GSL8 is required for cytokinesis and cell patterning. Plant Physiol. 2009, 150, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Gillmor, C.S.; Lukowitz, W.; Brininstool, G.; Sedbrook, J.C.; Hamann, T.; Poindexter, P.; Somerville, C. Glycosylphosphatidylinositol-anchored proteins are required for cell wall synthesis and morphogenesis in Arabidopsis. Plant Cell 2005, 17, 1128–1140. [Google Scholar] [CrossRef]
- Mellor, N.L.; Voss, U.; Janes, G.; Bennett, M.J.; Wells, D.M.; Band, L.R. Auxin fluxes through plasmodesmata modify root-tip auxin distribution. Development 2020, 147. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.; Liu, X.; De Storme, N.; Jensen, K.H.; Xu, Q.; Yang, J.; Liu, X.; Chen, S.; Martens, H.J.; Schulz, A.; et al. Directionality of Plasmodesmata-Mediated Transport in Arabidopsis Leaves Supports Auxin Channeling. Curr. Biol. 2020, 30, 1970–1977 e1974. [Google Scholar] [CrossRef]
- Han, X.; Hyun, T.K.; Zhang, M.; Kumar, R.; Koh, E.J.; Kang, B.H.; Lucas, W.J.; Kim, J.Y. Auxin-callose-mediated plasmodesmal gating is essential for tropic auxin gradient formation and signaling. Dev. Cell 2014, 28, 132–146. [Google Scholar] [CrossRef] [Green Version]
- Jackson, D. Plasmodesmata spread their influence. F1000Prime Rep. 2015, 7, 25. [Google Scholar] [CrossRef] [Green Version]
- Sager, R.; Wang, X.; Hill, K.; Yoo, B.C.; Caplan, J.; Nedo, A.; Tran, T.; Bennett, M.J.; Lee, J.Y. Auxin-dependent control of a plasmodesmal regulator creates a negative feedback loop modulating lateral root emergence. Nat. Commun. 2020, 11, 364. [Google Scholar] [CrossRef]
- Xu, B.; Cheval, C.; Laohavisit, A.; Hocking, B.; Chiasson, D.; Olsson, T.S.G.; Shirasu, K.; Faulkner, C.; Gilliham, M. A calmodulin-like protein regulates plasmodesmal closure during bacterial immune responses. New Phytol. 2017, 215, 77–84. [Google Scholar] [CrossRef] [Green Version]
- De Storme, N.; Geelen, D. Callose homeostasis at plasmodesmata: Molecular regulators and developmental relevance. Front. Plant Sci. 2014, 5, 138. [Google Scholar] [CrossRef] [Green Version]
- Zavaliev, R.; Ueki, S.; Epel, B.L.; Citovsky, V. Biology of callose (beta-1,3-glucan) turnover at plasmodesmata. Protoplasma 2011, 248, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, V.; Vashisht, D.; Cletus, J.; Sakthivel, N. Plant beta-1,3-glucanases: Their biological functions and transgenic expression against phytopathogenic fungi. Biotechnol. Lett. 2012, 34, 1983–1990. [Google Scholar] [CrossRef] [PubMed]
- Benitez-Alfonso, Y.; Faulkner, C.; Pendle, A.; Miyashima, S.; Helariutta, Y.; Maule, A. Symplastic intercellular connectivity regulates lateral root patterning. Dev. Cell 2013, 26, 136–147. [Google Scholar] [CrossRef] [Green Version]
- Kotake, T.; Nakagawa, N.; Takeda, K.; Sakurai, N. Auxin-Induced Elongation Growth and Expressions of Cell Wall-Bound Exoand Endo-b-Glucanases in Barley Coleoptiles. Plant Cell Physiol. 2000, 41, 1272–1278. [Google Scholar] [CrossRef] [Green Version]
- Tucker, E.B.; Boss, W.F. Mastoparan-lnduced lntracellular Ca2+ Fluxes May Regulate CeII-to-CeII communication in PIants. Plant Physiol. 1996, 11, 459–467. [Google Scholar] [CrossRef] [Green Version]
- Holdaway-Clarke, T.L.; Walker, N.A.; Heple, P.K.; Overall, R.L. Physiological elevations in cytoplasmic free calcium by cold or ion injection result in transient closure of higher plant plasmodesmata. Planta 2000, 210, 329–335. [Google Scholar] [CrossRef]
- Overall, R.L.; Blackman, L.M. A model of the macromolecular structure of plasmodesmata. Trends Plant Sci. 1996, 9, 307–311. [Google Scholar] [CrossRef]
- Baluska, F.; Cvrckova, F.; Kendrick-Jones, J.; Volkmann, D. Sink Plasmodesmata as Gateways for Phloem Unloading. Myosin VIII and Calreticulin as Molecular Determinants of Sink Strength. Plant Physiol. 2001, 126, 39–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilsner, J.; Nicolas, W.; Rosado, A.; Bayer, E.M. Staying Tight: Plasmodesmal Membrane Contact Sites and the Control of Cell-to-Cell Connectivity in Plants. Annu. Rev. Plant Biol. 2016, 67, 337–364. [Google Scholar] [CrossRef]
- Tominaga, M.; Kojima, H.; Yokota, E.; Nakamori, R.; Anson, M.; Shimmen, T.; Oiwa, K. Calcium-induced Mechanical Change in the Neck Domain Alters the Activity of Plant Myosin XI. J. Biol. Chem. 2012, 287, 30711–30718. [Google Scholar] [CrossRef] [Green Version]
- Vanneste, S.; Friml, J. Calcium: The Missing Link in Auxin Action. Plants 2013, 2, 650–675. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Hernandez, V.; Benitez, M.; Boudaoud, A. Interplay between turgor pressure and plasmodesmata during plant development. J. Exp. Bot. 2020, 71, 768–777. [Google Scholar] [CrossRef] [PubMed]
- Brunkard, J.O.; Xu, M.; Scarpin, M.R.; Chatterjee, S.; Shemyakina, E.A.; Goodman, H.M.; Zambryski, P. TOR dynamically regulates plant cell-cell transport. Proc. Natl. Acad. Sci. USA 2020, 117, 5049–5058. [Google Scholar] [CrossRef] [PubMed]
- Schepetilnikov, M.; Ryabova, L.A. Auxin Signaling in Regulation of Plant Translation Reinitiation. Front. Plant Sci 2017, 8, 1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oparka, K.J.; Prior, D.A.M. Direct evidence for pressure-generated closure of plasmodesmata. Plant J. 1992, 2, 741–750. [Google Scholar] [CrossRef]
- Park, K.; Knoblauch, J.; Oparka, K.; Jensen, K.H. Controlling intercellular flow through mechanosensitive plasmodesmata nanopores. Nat. Commun. 2019, 10, 3564. [Google Scholar] [CrossRef] [PubMed]
- Dreyer, I.; Uozumi, N. Potassium channels in plant cells. FEBS J. 2011, 278, 4293–4303. [Google Scholar] [CrossRef]
- Jahn, L.; Mucha, S.; Bergmann, S.; Horn, C.; Staswick, P.; Steffens, B.; Siemens, J.; Ludwig-Muller, J. The Clubroot Pathogen (Plasmodiophora brassicae) Influences Auxin Signaling to Regulate Auxin Homeostasis in Arabidopsis. Plants 2013, 2, 726–749. [Google Scholar] [CrossRef] [Green Version]
- Philippar, K.; Ivashikina, N.; Ache, P.; Christian, M.; Luthen, H.; Palme, K.; Hedrich, R. Auxin activates KAT1 and KAT2, two K+-channel genes expressed in seedlings of Arabidopsis thaliana. Plant J. 2004, 37, 815–827. [Google Scholar] [CrossRef]
- Philippar, K.; Buchsenschutz, K.; Edwards, D.; Loffler, J.; Luthen, H.; Kranz, E.; Edwards, K.J.; Hedrich, R. The auxin-induced K(+) channel gene Zmk1 in maize functions in coleoptile growth and is required for embryo development. Plant Mol. Biol. 2006, 61, 757–768. [Google Scholar] [CrossRef]
- Bauly, J.M.; Sealy, I.M.; Macdonald, H.; Brearley, J.; Droge, S.; Hillmer, S.; Robinson, D.G.; Venis, M.A.; Blatt, M.R.; Lazarus, C.M.; et al. Overexpression of Auxin-Binding Protein Enhances the Sensitivity of Guard Cells to Auxin. Plant Physiol. 2000, 124, 1229–1238. [Google Scholar] [CrossRef] [Green Version]
- Thiel, G.; Blatt, M.R.; Fricker, M.D.; White, I.R.; Millner, P. Modulation of K+ channels in Vicia stomatal guard cells by peptide homologs to the auxin-binding protein C terminus. Proc. Natl. Acad. Sci. USA 1993, 90, 11493–11497. [Google Scholar] [CrossRef] [Green Version]
- Christensen, N.M.; Faulkner, C.; Oparka, K. Evidence for unidirectional flow through plasmodesmata. Plant Physiol. 2009, 150, 96–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; He, Y.; Zhao, J.; Zhang, L.; Sun, M.X. Polar protein transport between apical and basal cells during tobacco early embryogenesis. Plant Cell Rep. 2013, 32, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Mettbach, U.; Strnad, M.; Mancuso, S.; Baluska, F. Immunogold-EM analysis reveal brefeldin a-sensitive clusters of auxin in Arabidopsis root apex cells. Commun. Integr. Biol. 2017, 10, e1327105. [Google Scholar] [CrossRef]
- Schlicht, M.; Strnad, M.; Scanlon, M.J.; Mancuso, S.; Hochholdinger, F.; Palme, K.; Volkmann, D.; Menzel, D.; Baluska, F. Auxin immunolocalization implicates vesicular neurotransmitter-like mode of polar auxin transport in root apices. Plant Signal. Behav. 2006, 1, 122–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baluska, F.; Strnad, M.; Mancuso, S. Substantial Evidence for Auxin Secretory Vesicles. Plant Physiol. 2018, 176, 2586–2587. [Google Scholar] [CrossRef] [Green Version]
- Robinson, D.G.; Hawes, C.; Hillmer, S.; Jurgens, G.; Schwechheimer, C.; Stierhof, Y.D.; Viotti, C. Auxin and Vesicle Traffic. Plant Physiol. 2018, 176, 1884–1888. [Google Scholar] [CrossRef] [PubMed]
- Hille, S.; Akhmanova, M.; Glanc, M.; Johnson, A.; Friml, J. Relative Contribution of PIN-Containing Secretory Vesicles and Plasma Membrane PINs to the Directed Auxin Transport: Theoretical Estimation. Int. J. Mol. Sci. 2018, 19, 3566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, C.Y.; Weijers, D. A toolkit for studying cellular reorganization during early embryogenesis in Arabidopsis thaliana. Plant J. 2018, 93, 963–976. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Winnicki, K.; Polit, J.T.; Żabka, A.; Maszewski, J. Do Plasmodesmata Play a Prominent Role in Regulation of Auxin-Dependent Genes at Early Stages of Embryogenesis? Cells 2021, 10, 733. https://doi.org/10.3390/cells10040733
Winnicki K, Polit JT, Żabka A, Maszewski J. Do Plasmodesmata Play a Prominent Role in Regulation of Auxin-Dependent Genes at Early Stages of Embryogenesis? Cells. 2021; 10(4):733. https://doi.org/10.3390/cells10040733
Chicago/Turabian StyleWinnicki, Konrad, Justyna Teresa Polit, Aneta Żabka, and Janusz Maszewski. 2021. "Do Plasmodesmata Play a Prominent Role in Regulation of Auxin-Dependent Genes at Early Stages of Embryogenesis?" Cells 10, no. 4: 733. https://doi.org/10.3390/cells10040733
APA StyleWinnicki, K., Polit, J. T., Żabka, A., & Maszewski, J. (2021). Do Plasmodesmata Play a Prominent Role in Regulation of Auxin-Dependent Genes at Early Stages of Embryogenesis? Cells, 10(4), 733. https://doi.org/10.3390/cells10040733