
S100A4 in the Physiology and Pathology of the Central and Peripheral Nervous System




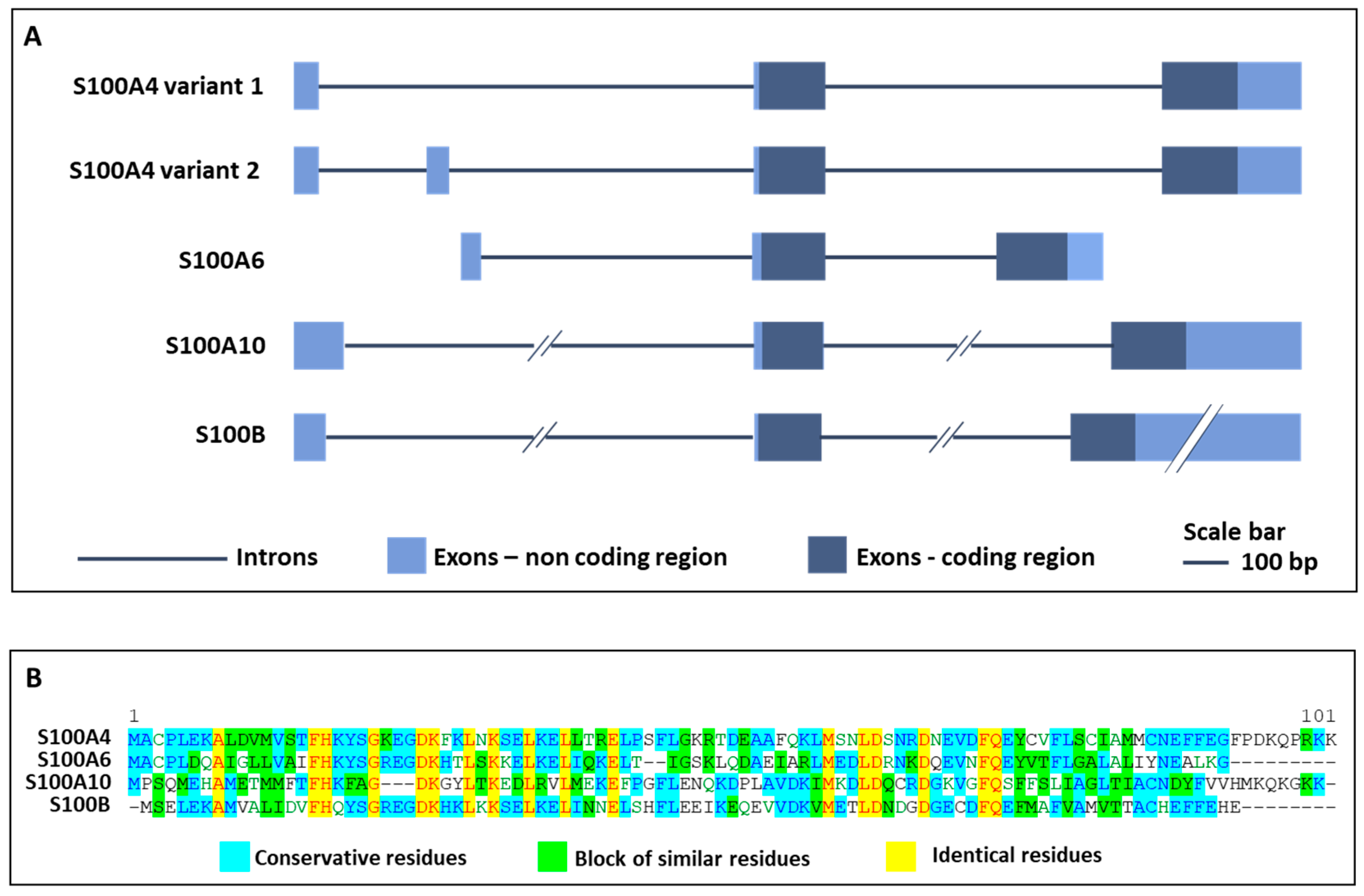{kind=link}
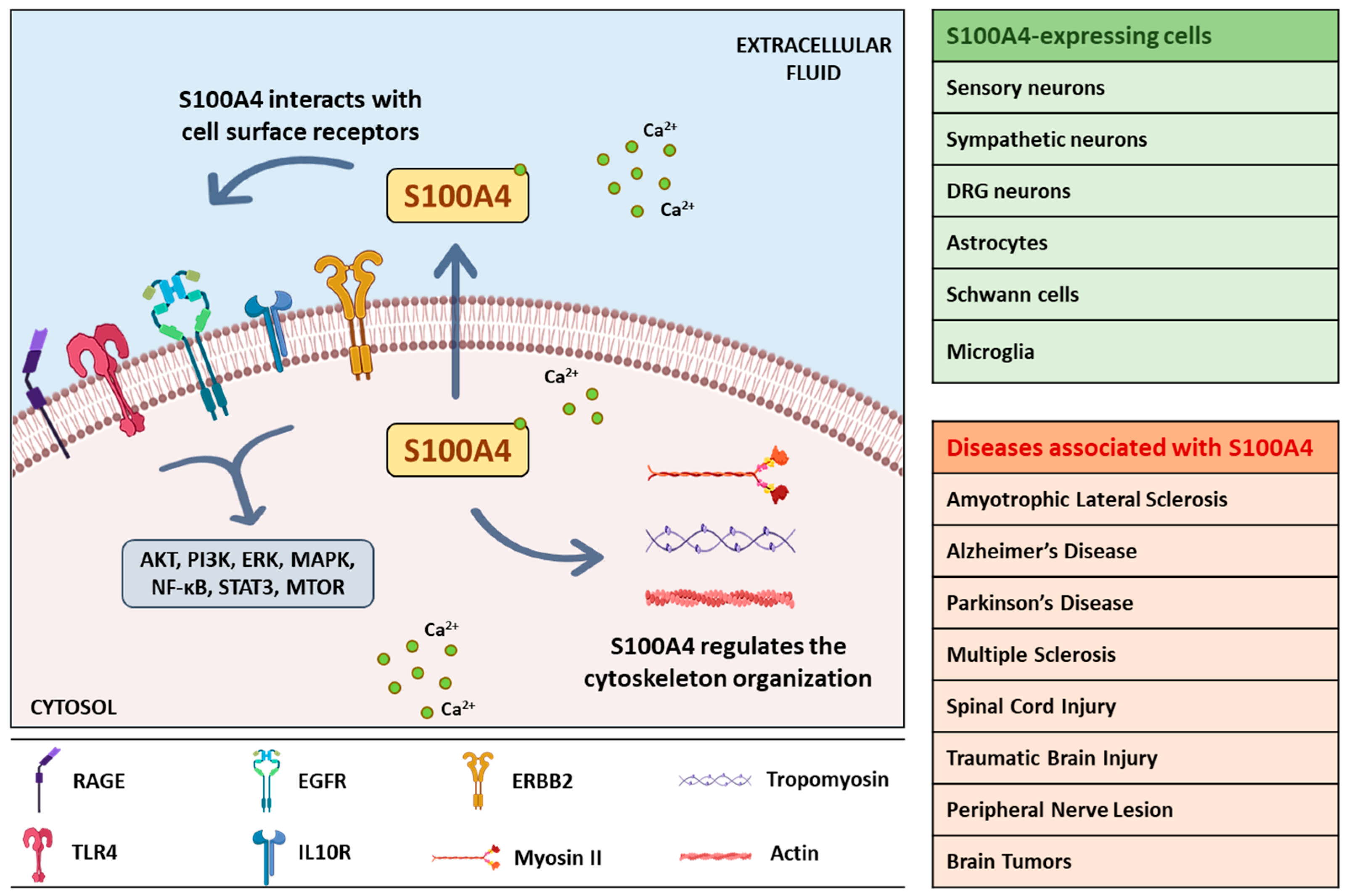{kind=link}
Abstract
:1. Introduction
2. S100A4 in the Nervous System Physiology
2.1. Neuronal Cells
2.2. Neuroglial Cells
3. S100A4 in the Pathology of the Nervous System
3.1. Brain Tumors
3.2. Neurodegenerative Disorders
3.3. Acute Injuries
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xia, H.; Gilbertsen, A.; Herrera, J.; Racila, E.; Smith, K.; Peterson, M.; Griffin, T.; Benyumov, A.; Yang, L.; Bitterman, P.B.; et al. Calcium-binding protein S100A4 confers mesenchymal progenitor cell fibrogenicity in idiopathic pulmonary fibrosis. J. Clin. Investig. 2017, 127, 2586–2597. [Google Scholar] [CrossRef] [Green Version]
- Donato, R.; Cannon, B.R.; Sorci, G.; Riuzzi, F.; Hsu, K.; Weber, D.J.; Geczy, C.L. Functions of S100 Proteins. Curr. Mol. Med. 2013, 13, 24–57. [Google Scholar] [CrossRef] [Green Version]
- Bresnick, A.R.; Weber, D.J.; Zimmer, D.B. S100 proteins in cancer. Nat. Rev. Cancer 2015, 15, 96–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miranda, K.J.; Loeser, R.F.; Yammani, R.R. Sumoylation and Nuclear Translocation of S100A4 Regulate IL-1β-mediated Production of Matrix Metalloproteinase-13. J. Biol. Chem. 2010, 285, 31517–31524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alanazi, B.; Munje, C.R.; Rastogi, N.; Williamson, A.J.K.; Taylor, S.; Hole, P.S.; Hodges, M.; Doyle, M.; Baker, S.; Gilkes, A.F.; et al. Integrated nuclear proteomics and transcriptomics identifies S100A4 as a therapeutic target in acute myeloid leukemia. Leukemia 2019, 34, 427–440. [Google Scholar] [CrossRef]
- Garrett, S.C.; Varney, K.M.; Weber, D.J.; Bresnick, A.R. S100A4, a Mediator of Metastasis. J. Biol. Chem. 2006, 281, 677–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boye, K.; Mælandsmo, G.M. S100A4 and Metastasis: A Small Actor Playing Many Roles. Am. J. Pathol. 2010, 176, 528–535. [Google Scholar] [CrossRef] [PubMed]
- Fei, F.; Qu, J.; Li, C.; Wang, X.; Li, Y.; Zhang, S. Role of metastasis-induced protein S100A4 in human non-tumor pathophysiologies. Cell Biosci. 2017, 7, 64. [Google Scholar] [CrossRef]
- Ambartsumian, N.; Klingelhöfer, J.; Grigorian, M. The Multifaceted S100A4 Protein in Cancer and Inflammation. Adv. Struct. Saf. Stud. 2019, 1929, 339–365. [Google Scholar] [CrossRef]
- Dulyaninova, N.G.; Ruiz, P.D.; Gamble, M.J.; Backer, J.M.; Bresnick, A.R. S100A4 regulates macrophage invasion by distinct myosin-dependent and myosin-independent mechanisms. Mol. Biol. Cell 2018, 29, 632–642. [Google Scholar] [CrossRef]
- Li, Z.; Li, Y.; Liu, S.; Qin, Z. Extracellular S100A4 as a key player in fibrotic diseases. J. Cell. Mol. Med. 2020, 24, 5973–5983. [Google Scholar] [CrossRef] [Green Version]
- Destek, S.; Gul, V.O. S100A4 May Be a Good Prognostic Marker and a Therapeutic Target for Colon Cancer. J. Oncol. 2018, 2018, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Ganaie, A.A.; Mansini, A.P.; Hussain, T.; Rao, A.; Siddique, H.R.; Shabaneh, A.; Ferrari, M.G.; Murugan, P.; Klingelhöfer, J.; Wang, J.; et al. Anti-S100A4 Antibody Therapy Is Efficient in Treating Aggressive Prostate Cancer and Reversing Immunosuppression: Serum and Biopsy S100A4 as a Clinical Predictor. Mol. Cancer Ther. 2020, 19, 2598–2611. [Google Scholar] [CrossRef]
- Bai, H.; Qian, J.-L.; Han, B.-H. S100A4 Is an Independent Prognostic Factor for Patients with Lung Cancer: A Meta-Analysis. Genet. Test. Mol. Biomarkers 2014, 18, 371–374. [Google Scholar] [CrossRef]
- Cao, C.-M.; Yang, F.-X.; Wang, P.-L.; Yang, Q.-X.; Sun, X.-R. Clinicopathologic significance of S100A4 expression in osteosarcoma. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 833–839. [Google Scholar] [PubMed]
- Yang, Q.; Zhou, Y.; Li, Z.; Ding, Y.; Zhang, J.; Wu, Y. Overexpression of S100A4 protein may be associated with the development and progression of pancreatic cancer. J. Cancer Res. Ther. 2018, 14, 159. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.-H.; Dulyaninova, N.G.; House, R.P.; Almo, S.C.; Bresnick, A.R. S100A4 Regulates Macrophage Chemotaxis. Mol. Biol. Cell 2010, 21, 2598–2610. [Google Scholar] [CrossRef] [Green Version]
- Michetti, F.; D’Ambrosi, N.; Toesca, A.; Puglisi, M.A.; Serrano, A.; Marchese, E.; Corvino, V.; Geloso, M.C. The S100B story: From biomarker to active factor in neural injury. J. Neurochem. 2018, 148, 168–187. [Google Scholar] [CrossRef] [Green Version]
- Filipek, A.; Leśniak, W. S100A6 and Its Brain Ligands in Neurodegenerative Disorders. Int. J. Mol. Sci. 2020, 21, 3979. [Google Scholar] [CrossRef] [PubMed]
- García-Morales, V.; Rodríguez-Bey, G.; Gómez-Pérez, L.; Domínguez-Vías, G.; González-Forero, D.; Portillo, F.; Campos-Caro, A.; Gento-Caro, Á.; Issaoui, N.; Soler, R.M.; et al. Sp1-regulated expression of p11 contributes to motor neuron degeneration by membrane insertion of TASK1. Nat. Commun. 2019, 10, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Kozlova, E.N.; Lukanidin, E. Metastasis-Associated Mts1 (S100A4) Protein Is Selectively Expressed in White Matter Astrocytes and Is up-Regulated after Peripheral Nerve or Dorsal Root Injury. Glia 1999, 27, 249–258. [Google Scholar] [CrossRef]
- Sandelin, M.; Zabihi, S.; Liu, L.; Wicher, G.; Kozlova, E.N. Metastasis-associated S100A4 (Mts1) protein is expressed in subpopulations of sensory and autonomic neurons and in Schwann cells of the adult rat. J. Comp. Neurol. 2004, 473, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Moldovan, M.; Pinchenko, V.; Dmytriyeva, O.; Pankratova, S.; Fugleholm, K.; Klingelhofer, J.; Bock, E.; Berezin, V.; Krarup, C.; Kiryushko, D. Peptide Mimetic of the S100A4 Protein Modulates Peripheral Nerve Regeneration and Attenuates the Progression of Neuropathy in Myelin Protein P0 Null Mice. Mol. Med. 2013, 19, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Trolle, C.; Ivert, P.; Hoeber, J.; Rocamonde-Lago, I.; Vasylovska, S.; Lukanidin, E.; Kozlova, E.N. Boundary cap neural crest stem cell transplants contribute Mts1/S100A4-expressing cells in the glial scar. Regen. Med. 2017, 12, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Schmid, D.; Zeis, T.; Sobrio, M.; Schaeren-Wiemers, N. MAL Overexpression Leads to Disturbed Expression of Genes That Influence Cytoskeletal Organization and Differentiation of Schwann Cells. ASN Neuro 2014, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.Y.; Xia, C.-L.; Dong, D.-C.; Heizmann, C.W.; Yew, D.T. Differential expression of S100 proteins in the developing human hippocampus and temporal cortex. Microsc. Res. Tech. 2003, 60, 600–613. [Google Scholar] [CrossRef]
- Aberg, F.; Kozlova, E.N. Metastasis-Associated Mts1 (S100A4) Protein in the Developing and Adult Central Nervous System. J. Comp. Neurol. 2000, 424, 269–282. [Google Scholar]
- Hoyaux, D.; Decaestecker, C.; Heizmann, C.W.; Vogl, T.; Schäfer, B.W.; Salmon, I.; Kiss, R.; Pochet, R. S100 proteins in Corpora Amylacea from normal human brain11Published on the World Wide Web on 5 May 2000. Brain Res. 2000, 867, 280–288. [Google Scholar] [CrossRef]
- Novitskaya, V.; Grigorian, M.; Kriajevska, M.; Tarabykina, S.; Bronstein, I.; Berezin, V.; Bock, E.; Lukanidin, E. Oligomeric Forms of the Metastasis-related Mts1 (S100A4) Protein Stimulate Neuronal Differentiation in Cultures of Rat Hippocampal Neurons. J. Biol. Chem. 2000, 275, 41278–41286. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, M.V.; Køhler, L.B.; Grigorian, M.; Novitskaya, V.; Bock, E.; Lukanidin, E.; Berezin, V. The Mts1/S100A4 protein is a neuroprotectant. J. Neurosci. Res. 2004, 77, 777–786. [Google Scholar] [CrossRef]
- Kiryushko, D.; Novitskaya, V.; Soroka, V.; Klingelhofer, J.; Lukanidin, E.; Berezin, V.; Bock, E. Molecular Mechanisms of Ca2+ Signaling in Neurons Induced by the S100A4 Protein. Mol. Cell. Biol. 2006, 26, 3625–3638. [Google Scholar] [CrossRef] [Green Version]
- Fang, Z.; Forslund, N.; Takenaga, K.; Lukanidin, E.; Kozlova, E. Sensory neurite outgrowth on white matter astrocytes is influenced by intracellular and extracellular S100A4 protein. J. Neurosci. Res. 2006, 83, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.; Tang, L. Schwann cells genetically modified to express S100A4 increases GAP43 expression in spiral ganglion neurons in vitro. Bioengineered 2016, 8, 404–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrano, A.; Apolloni, S.; Rossi, S.; Lattante, S.; Sabatelli, M.; Peric, M.; Andjus, P.; Michetti, F.; Carrì, M.T.; Cozzolino, M.; et al. The S100A4 Transcriptional Inhibitor Niclosamide Reduces Pro-Inflammatory and Migratory Phenotypes of Microglia: Implications for Amyotrophic Lateral Sclerosis. Cells 2019, 8, 1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozlova, E.N.; Lukanidin, E. Mts1 Protein Expression in the Central Nervous System after Injury. Glia 2002, 37, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Gross, S.R.; Sin, C.G.T.; Barraclough, R.; Rudland, P.S. Joining S100 proteins and migration: For better or for worse, in sickness and in health. Cell. Mol. Life Sci. 2013, 71, 1551–1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takenaga, K.; Kozlova, E.N. Role of intracellular S100A4 for migration of rat astrocytes. Glia 2005, 53, 313–321. [Google Scholar] [CrossRef]
- Fang, Z.; Duthoit, N.; Wicher, G.; Kallskog, O.; Ambartsumian, N.; Lukanidin, E.; Takenaga, K.; Kozlova, E. Intracellular calcium-binding protein S100A4 influences injury-induced migration of white matter astrocytes. Acta Neuropathol. 2006, 111, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Wang, Y.; Zhang, J.; Tu, J.; Wang, X.-J.; Su, X.-D.; Wang, L.; Zhang, Y. Tunneling-nanotube direction determination in neurons and astrocytes. Cell Death Dis. 2012, 3, e438. [Google Scholar] [CrossRef]
- Srinivasan, K.; Friedman, B.A.; Etxeberria, A.; Huntley, M.A.; van der Brug, M.P.; Foreman, O.; Paw, J.S.; Modrusan, Z.; Beach, T.G.; Serrano, G.E.; et al. Alzheimer’s Patient Microglia Exhibit Enhanced Aging and Unique Transcriptional Activation. Cell Rep. 2020, 31, 107843. [Google Scholar] [CrossRef]
- Li, Y.; Bao, J.; Bian, Y.; Erben, U.; Wang, P.; Song, K.; Liu, S.; Li, Z.; Gao, Z.; Qin, Z. S100A4+ Macrophages Are Necessary for Pulmonary Fibrosis by Activating Lung Fibroblasts. Front. Immunol. 2018, 9, 1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orre, L.M.; Panizza, E.O.; Kaminskyy, V.; Vernet, E.; Gräslund, T.; Zhivotovsky, B.; Lehtiö, J. S100A4 interacts with p53 in the nucleus and promotes p53 degradation. Oncogene 2013, 32, 5531–5540. [Google Scholar] [CrossRef] [PubMed]
- Pan, C.-X.; Lara, P.; Evans, C.P.; Parikh, M.; Dall’Era, M.; Liu, C.; Robles, D.; Gao, A. Niclosamide in combination with abiraterone and prednisone in men with castration-resistant prostate cancer (CRPC): Initial results from a phase Ib/II trial. J. Clin. Oncol. 2018, 36, 192. [Google Scholar] [CrossRef]
- Burock, S.; Daum, S.; Keilholz, U.; Neumann, K.; Walther, W.; Stein, U. Phase II trial to investigate the safety and efficacy of orally applied niclosamide in patients with metachronous or sychronous metastases of a colorectal cancer progressing after therapy: The NIKOLO trial. BMC Cancer 2018, 18, 1–7. [Google Scholar] [CrossRef]
- Fong, W.; To, K.K.W. Drug repurposing to overcome resistance to various therapies for colorectal cancer. Cell. Mol. Life Sci. 2019, 76, 3383–3406. [Google Scholar] [CrossRef]
- Chow, K.-H.; Park, H.J.; George, J.; Yamamoto, K.; Gallup, A.D.; Graber, J.H.; Chen, Y.; Jiang, W.; Steindler, D.A.; Neilson, E.G.; et al. S100A4 Is a Biomarker and Regulator of Glioma Stem Cells That Is Critical for Mesenchymal Transition in Glioblastoma. Cancer Res. 2017, 77, 5360–5373. [Google Scholar] [CrossRef] [Green Version]
- Lindsey, J.C.E.; Lusher, M.A.; Anderton, J.; Gilbertson, R.J.; Ellison, D.W.; Clifford, S.C. Epigenetic deregulation of multiple S100 gene family members by differential hypomethylation and hypermethylation events in medulloblastoma. Br. J. Cancer 2007, 97, 267–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernan, R.; Fasheh, R.; Calabrese, C.; Frank, A.J.; MacLean, K.H.; Allard, D.; Barraclough, R.; Gilbertson, R.J. ERBB2 up-regulates S100A4 and several other prometastatic genes in medulloblastoma. Cancer Res. 2003, 63, 140–148. [Google Scholar]
- Rand, V.; Prebble, E.; Ridley, L.; Howard, M.; Wei, W.; Brundler, M.-A.; Fee, B.E.; Riggins, G.J.; Coyle, B.; Grundy, R.G.; et al. Investigation of chromosome 1q reveals differential expression of members of the S100 family in clinical subgroups of intracranial paediatric ependymoma. Br. J. Cancer 2008, 99, 1136–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.; Zhao, C.; Zhao, Z.; Hu, L.; Ye, F.; Wang, H.; Fang, Z.; Wu, Y.; Chen, X. Specific glioblastoma multiforme prognostic-subtype distinctions based on DNA methylation patterns. Cancer Gene Ther. 2019, 27, 1–13. [Google Scholar] [CrossRef]
- Tang, Y.; Qing, C.; Wang, J.; Zeng, Z. DNA Methylation-based Diagnostic and Prognostic Biomarkers for Glioblastoma. Cell Transplant. 2020, 29, 963689720933241. [Google Scholar] [CrossRef] [PubMed]
- Takenaga, K.; Nygren, J.; Zelenina, M.; Ohira, M.; Iuchi, T.; Lukanidin, E.; Sjöquist, M.; Kozlova, E.N. Modified expression of Mts1/S100A4 protein in C6 glioma cells or surrounding astrocytes affects migration of tumor cells in vitro and in vivo. Neurobiol. Dis. 2007, 25, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Belot, N.; Pochet, R.; Heizmann, C.W.; Kiss, R.; Decaestecker, C. Extracellular S100A4 stimulates the migration rate of astrocytic tumor cells by modifying the organization of their actin cytoskeleton. Biochim. et Biophys. Acta (BBA)—Proteins Proteom. 2002, 1600, 74–83. [Google Scholar] [CrossRef]
- Silva, J.M.; Wippel, H.H.; Santos, M.D.M.; Verissimo, D.C.A.; Santos, R.M.; Nogueira, F.C.S.; Passos, G.A.R.; Sprengel, S.L.; Borba, L.A.B.; Carvalho, P.C.; et al. Proteomics pinpoints alterations in grade I meningiomas of male versus female patients. Sci. Rep. 2020, 10, 10335. [Google Scholar] [CrossRef]
- Pankratova, S.; Klingelhofer, J.; Dmytriyeva, O.; Owczarek, S.; Renziehausen, A.; Syed, N.; Porter, A.E.; Dexter, D.T.; Kiryushko, D. The S100A4 Protein Signals through the ErbB4 Receptor to Promote Neuronal Survival. Theranostics 2018, 8, 3977–3990. [Google Scholar] [CrossRef] [PubMed]
- Mei, L.; Nave, K.-A. Neuregulin-ERBB signaling in the nervous system and neuropsychiatric diseases. Neuron 2014, 83, 27–49. [Google Scholar] [CrossRef] [Green Version]
- Earhart, G.M.; Clark, B.R.; Tabbal, S.D.; Perlmutter, J.S. Gait and balance in essential tremor: Variable effects of bilateral thalamic stimulation. Mov. Disord. 2009, 24, 386–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sydow, O.; Thobois, S.; Alesch, F.; Speelman, J.D. Multicentre European study of thalamic stimulation in essential tremor: A six year follow up. J. Neurol. Neurosurg. Psychiatry 2003, 74, 1387–1391. [Google Scholar] [CrossRef]
- Kádár, E.; Lim, L.W.; Carreras, G.; Genis, D.; Temel, Y.; Huguet, G. High-frequency stimulation of the ventrolateral thalamus regulates gene expression in hippocampus, motor cortex and caudate–putamen. Brain Res. 2011, 1391, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Münch, A.E.; Chung, W.-S.; Peterson, T.C.; et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef]
- D’Ambrosi, N.; Apolloni, S. Fibrotic Scar in Neurodegenerative Diseases. Front. Immunol. 2020, 11, 1394. [Google Scholar] [CrossRef]
- Sofroniew, M.V. Molecular dissection of reactive astrogliosis and glial scar formation. Trends Neurosci. 2009, 32, 638–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filipi, T.; Hermanova, Z.; Tureckova, J.; Vanatko, O.; Anderova, A.M. Glial Cells—The Strategic Targets in Amyotrophic Lateral Sclerosis Treatment. J. Clin. Med. 2020, 9, 261. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Sun, Y.; Ling, S.-C.; Ferraiuolo, L.; McAlonis-Downes, M.; Zou, Y.; Drenner, K.; Wang, Y.; Ditsworth, D.; Tokunaga, S.; et al. Translational profiling identifies a cascade of damage initiated in motor neurons and spreading to glia in mutant SOD1-mediated ALS. Proc. Natl. Acad. Sci. USA 2015, 112, E6993–E7002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turvey, M.E.; Koudelka, T.; Comerford, I.; Greer, J.M.; Carroll, W.; Bernard, C.C.A.; Hoffmann, P.; McColl, S.R. Quantitative Proteome Profiling of CNS-Infiltrating Autoreactive CD4+ Cells Reveals Selective Changes during Experimental Autoimmune Encephalomyelitis. J. Proteome Res. 2014, 13, 3655–3670. [Google Scholar] [CrossRef] [PubMed]
- Fissolo, N.; Matute-Blanch, C.; Osman, M.; Costa, C.; Pinteac, R.; Miró, B.; Sanchez, A.; Brito, V.; Dujmovic, I.; Voortman, M.; et al. CSF SERPINA3 Levels Are Elevated in Patients With Progressive MS. Neurol. Neuroimmunol. Neuroinflamm. 2021, 8, 2021. [Google Scholar] [CrossRef]
- Noori, A.; Mezlini, A.M.; Hyman, B.T.; Serrano-Pozo, A.; Das, S. Differential gene expression data from the human central nervous system across Alzheimer’s disease, Lewy body diseases, and the amyotrophic lateral sclerosis and frontotemporal dementia spectrum. Data Brief 2021, 35, 106863. [Google Scholar] [CrossRef]
- Noori, A.; Mezlini, A.M.; Hyman, B.T.; Serrano-Pozo, A.; Das, S. Systematic review and meta-analysis of human transcriptomics reveals neuroinflammation, deficient energy metabolism, and proteostasis failure across neurodegeneration. Neurobiol. Dis. 2021, 149, 105225. [Google Scholar] [CrossRef] [PubMed]
- Jassam, Y.N.; Izzy, S.; Whalen, M.; McGavern, D.B.; El Khoury, J. Neuroimmunology of Traumatic Brain Injury: Time for a Paradigm Shift. Neuron 2017, 95, 1246–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dmytriyeva, O.; Pankratova, S.; Owczarek, S.; Sonn, K.; Soroka, V.; Ridley, C.M.; Marsolais, A.; Lopez-Hoyos, M.; Ambartsumian, N.; Lukanidin, E.; et al. The metastasis-promoting S100A4 protein confers neuroprotection in brain injury. Nat. Commun. 2012, 3, 1197. [Google Scholar] [CrossRef] [Green Version]
- Lipponen, A.; Paananen, J.; Puhakka, N.; Pitkänen, A. Analysis of Post-Traumatic Brain Injury Gene Expression Signature Reveals Tubulins, Nfe2l2, Nfkb, Cd44 and S100a4 as Treatment Targets. Sci. Rep. 2016, 6, 31570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Yang, N.; Luo, J.; Cheng, G.; Zhang, X.; He, T.; Xing, Y. Overexpression of S100A4 protects retinal ganglion cells against retinal ischemia-reperfusion injury in mice. Exp. Eye Res. 2020, 201, 108281. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, E.J.; Burnside, E.R. Moving beyond the glial scar for spinal cord repair. Nat. Commun. 2019, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Fawcett, J.W.; Asher, R. The glial scar and central nervous system repair. Brain Res. Bull. 1999, 49, 377–391. [Google Scholar] [CrossRef]
- Zhang, K.-H.; Han, S.; Lu, P.-H.; Xu, X.-M. Upregulation of S100A4 after spinal cord transection in adult rats. Acta Pharmacol. Sin. 2004, 25, 1007–1012. [Google Scholar]
- Wróblewski, R.; Roomans, G.M.; Kozlova, E.N. Effects of Dorsal Root Transection on Morphology and Chemical Composition of Degenerating Nerve Fibers and Reactive Astrocytes in the Dorsal Funiculus. Exp. Neurol. 2000, 164, 236–245. [Google Scholar] [CrossRef]
- Chen, W.; Mook, R.A., Jr.; Premont, R.T.; Wang, J. Niclosamide: Beyond an antihelminthic drug. Cell. Signal. 2018, 41, 89–96. [Google Scholar] [CrossRef]
- Boyapally, R.; Pulivendala, G.; Bale, S.; Godugu, C. Niclosamide alleviates pulmonary fibrosis in vitro and in vivo by attenuation of epithelial-to-mesenchymal transition, matrix proteins & Wnt/β-catenin signaling: A drug repurposing study. Life Sci. 2019, 220, 8–20. [Google Scholar] [CrossRef]
- Cabrita, I.; Benedetto, R.; Schreiber, R.; Kunzelmann, K. Niclosamide repurposed for the treatment of inflammatory airway disease. JCI Insight 2019, 4, 4. [Google Scholar] [CrossRef]
- Cerles, O.; Benoit, E.; Chéreau, C.; Chouzenoux, S.; Morin, F.; Guillaumot, M.-A.; Coriat, R.; Kavian, N.; Loussier, T.; Santulli, P.; et al. Niclosamide Inhibits Oxaliplatin Neurotoxicity while Improving Colorectal Cancer Therapeutic Response. Mol. Cancer Ther. 2016, 16, 300–311. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Ambrosi, N.; Milani, M.; Apolloni, S. S100A4 in the Physiology and Pathology of the Central and Peripheral Nervous System. Cells 2021, 10, 798. https://doi.org/10.3390/cells10040798
D’Ambrosi N, Milani M, Apolloni S. S100A4 in the Physiology and Pathology of the Central and Peripheral Nervous System. Cells. 2021; 10(4):798. https://doi.org/10.3390/cells10040798
Chicago/Turabian StyleD’Ambrosi, Nadia, Martina Milani, and Savina Apolloni. 2021. "S100A4 in the Physiology and Pathology of the Central and Peripheral Nervous System" Cells 10, no. 4: 798. https://doi.org/10.3390/cells10040798
APA StyleD’Ambrosi, N., Milani, M., & Apolloni, S. (2021). S100A4 in the Physiology and Pathology of the Central and Peripheral Nervous System. Cells, 10(4), 798. https://doi.org/10.3390/cells10040798