
NF-κB and TNF Affect the Astrocytic Differentiation from Neural Stem Cells

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethic Statement
2.2. Cell Culture
2.3. NF-κB Inhibition
2.4. MTT Cell Viability Assay
2.5. Tunel Assay
2.6. Western Blot
2.7. Microarrays Experiments and Quality Control
2.8. Real-Time PCR
2.9. Immunocytochemistry
2.10. Microarray, Limma Analysis
2.11. Statistical Analysis
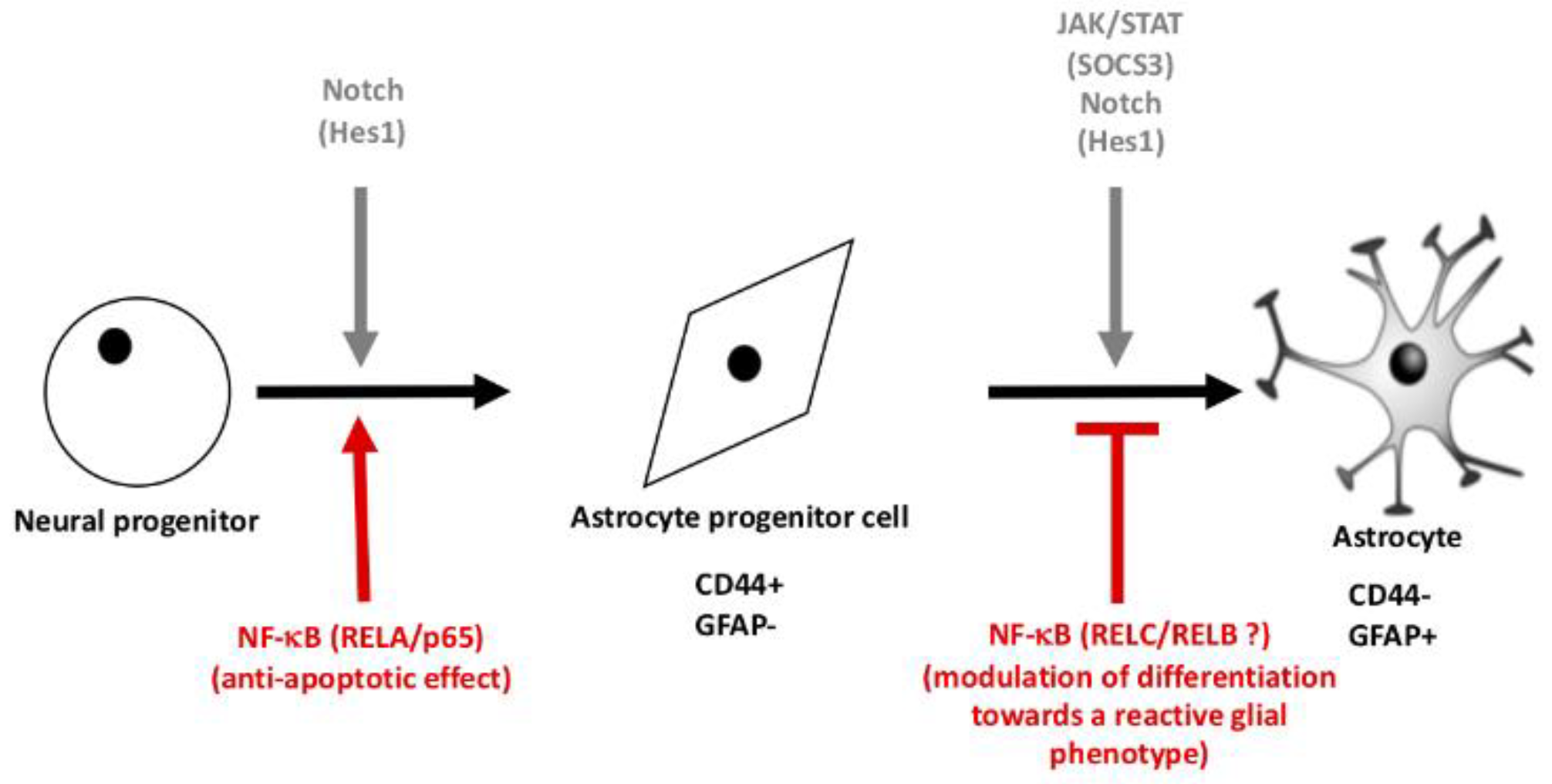
3. Results
3.1. NF-κB Is Essential for the Initiation of NPCs-Derived Astrocyte Differentiation
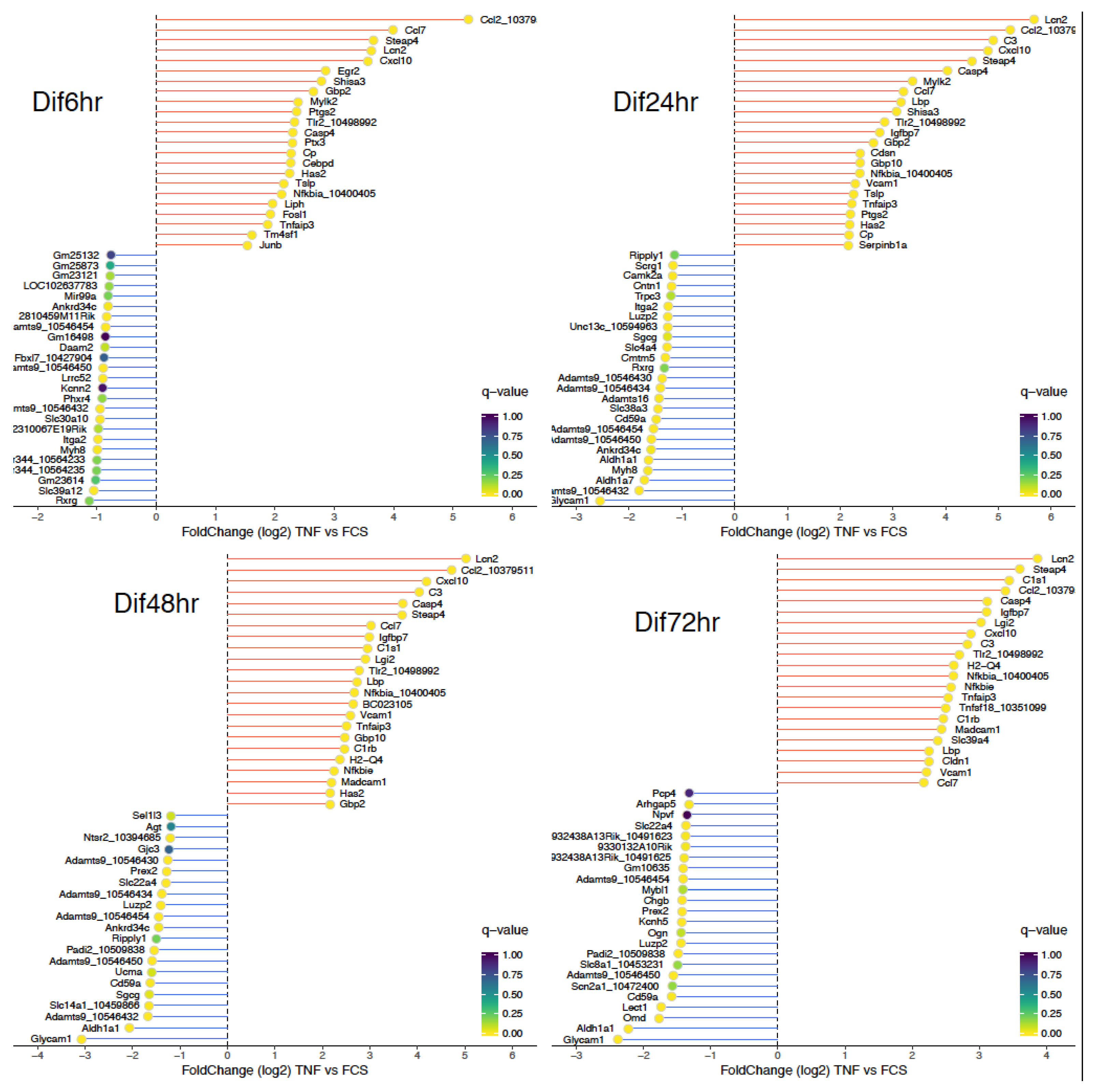
3.2. TNF-Treated NPCs Show an Inflammatory Phenotype Associated to NF-κB Activation
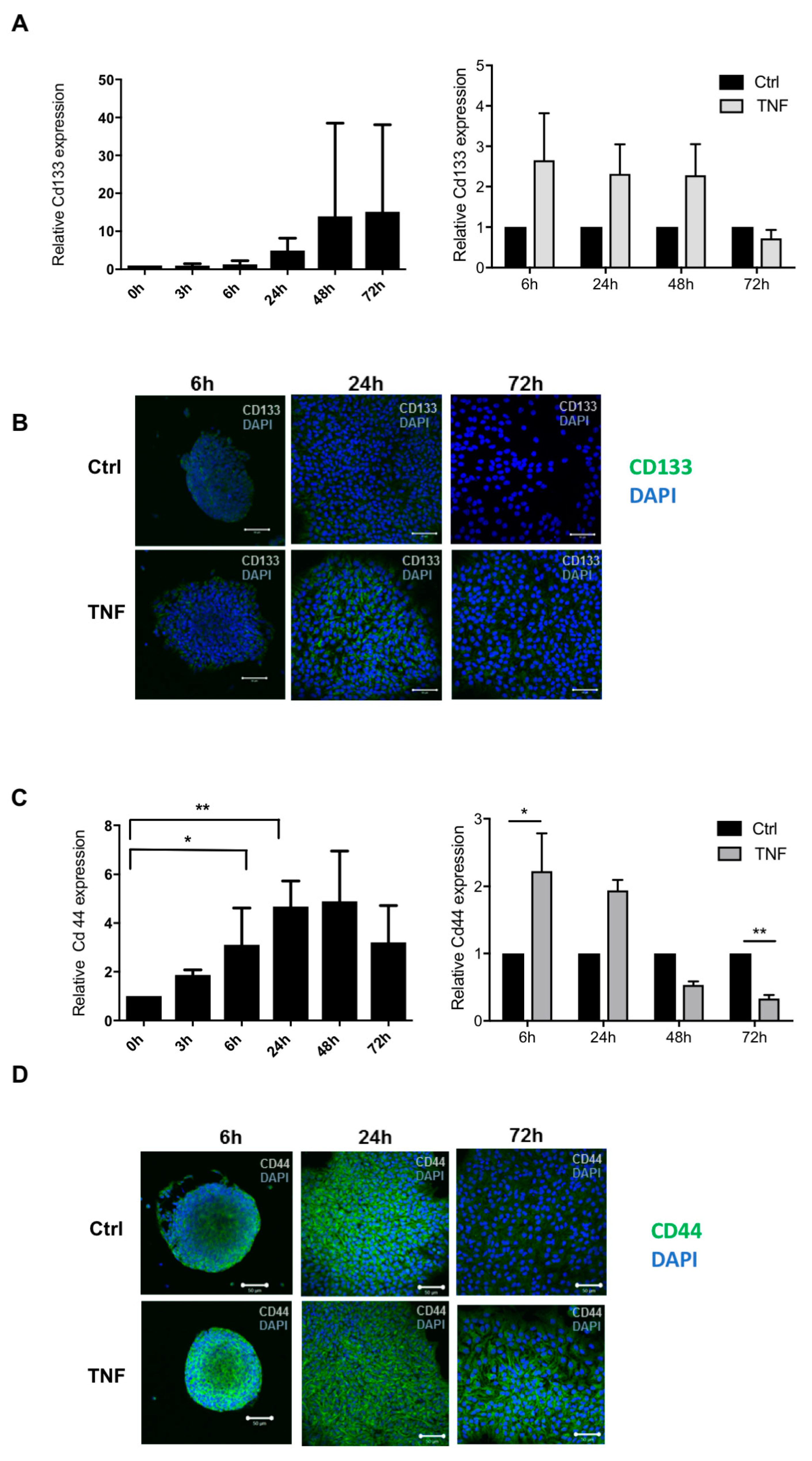
3.3. NF-κB Activation Following TNF Exposure Increases Immaturity Markers and Modulates Astrocytic Differentiation
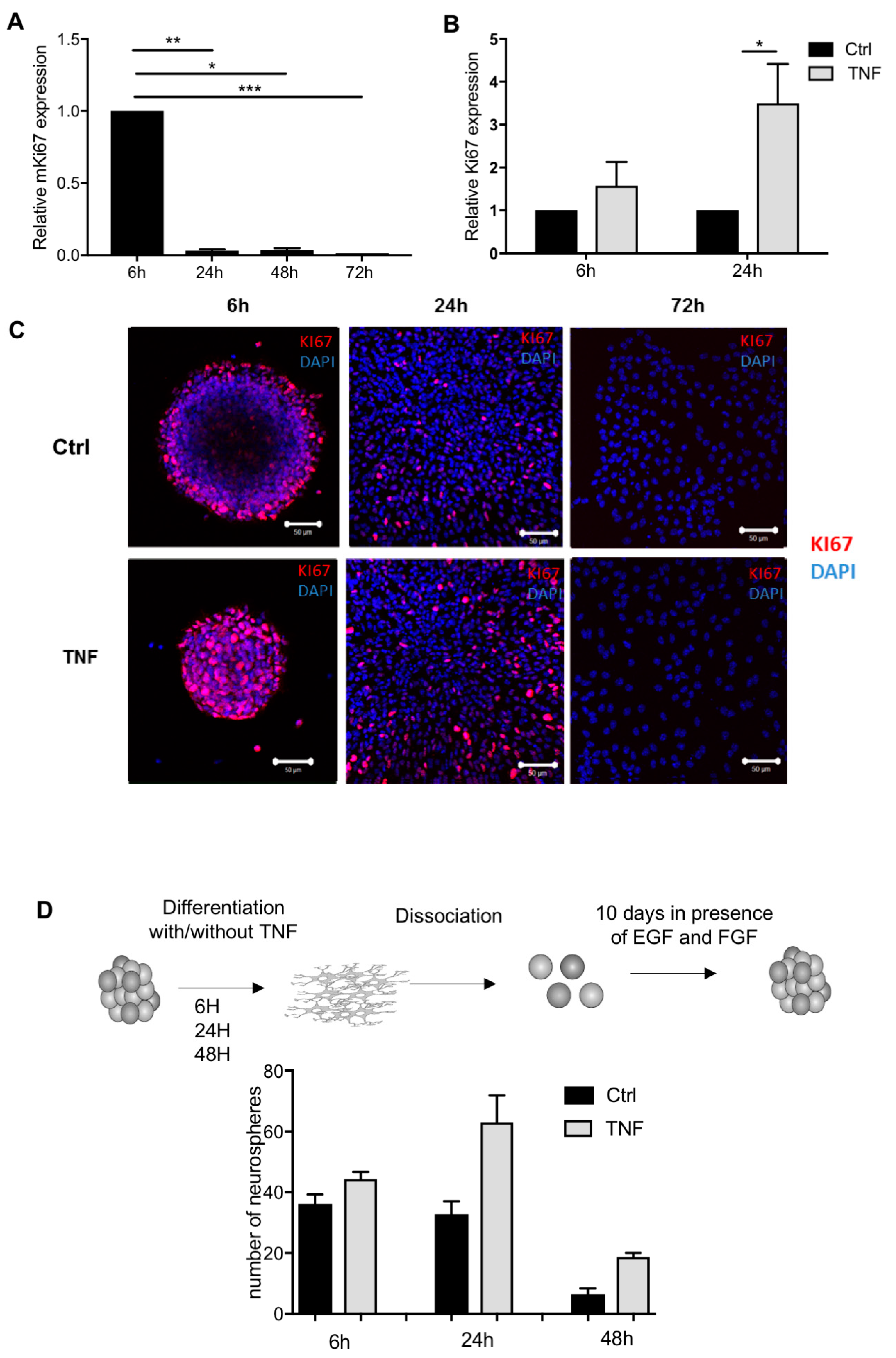
3.4. TNF-Treated NPCs Display Increased Proliferation and Maintain Multipotential Properties along Astrocyte Differentiation
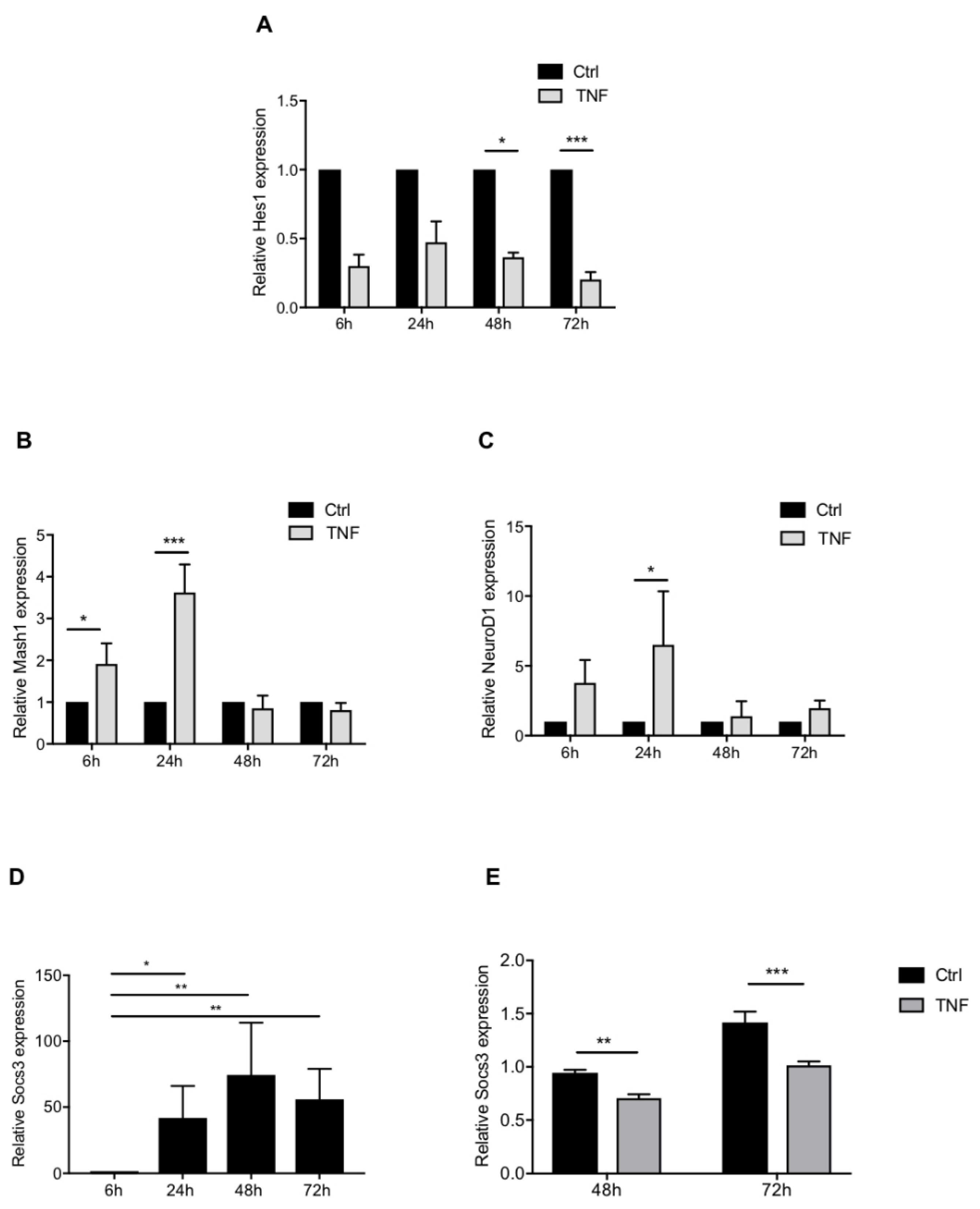
3.5. NOTCH and JAK-STAT Signaling Pathways Are Modulated in TNF-Treated NPCs-Derived Astrocytes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lang, B.; Liu, H.L.; Liu, R.; Feng, G.D.; Jiao, X.Y.; Ju, G. Astrocytes in injured adult rat spinal cord may acquire the potential of neural stem cells. Neuroscience 2004, 128, 775–783. [Google Scholar] [CrossRef]
- Buffo, A.; Rite, I.; Tripathi, P.; Lepier, A.; Colak, D.; Horn, A.P.; Mori, T.; Gotz, M. Origin and progeny of reactive gliosis: A source of multipotent cells in the injured brain. Proc. Natl. Acad. Sci. USA 2008, 105, 3581–3586. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Cheng, X.P.; Li, J.W.; Yao, Q.; Ju, G. De-differentiation response of cultured astrocytes to injury induced by scratch or conditioned culture medium of scratch-insulted astrocytes. Cell. Mol. Neurobiol. 2009, 29, 455–473. [Google Scholar] [CrossRef]
- Michelucci, A.; Bithell, A.; Burney, M.J.; Johnston, C.E.; Wong, K.Y.; Teng, S.W.; Desai, J.; Gumbleton, N.; Anderson, G.; Stanton, L.W.; et al. The Neurogenic Potential of Astrocytes Is Regulated by Inflammatory Signals. Mol. Neurobiol. 2016, 53, 3724–3739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharif, A.; Legendre, P.; Prevot, V.; Allet, C.; Romao, L.; Studler, J.M.; Chneiweiss, H.; Junier, M.P. Transforming growth factor alpha promotes sequential conversion of mature astrocytes into neural progenitors and stem cells. Oncogene 2007, 26, 2695–2706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabel, S.; Koncina, E.; Dorban, G.; Heurtaux, T.; Birck, C.; Glaab, E.; Michelucci, A.; Heuschling, P.; Grandbarbe, L. Inflammation Promotes a Conversion of Astrocytes into Neural Progenitor Cells via NF-kappaB Activation. Mol. Neurobiol. 2016, 53, 5041–5055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovacs, A.D.; Chakraborty-Sett, S.; Ramirez, S.H.; Sniderhan, L.F.; Williamson, A.L.; Maggirwar, S.B. Mechanism of NF-kappaB inactivation induced by survival signal withdrawal in cerebellar granule neurons. Eur. J. Neurosci. 2004, 20, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Grilli, M.; Memo, M. Nuclear factor-kappaB/Rel proteins: A point of convergence of signalling pathways relevant in neuronal function and dysfunction. Biochem. Pharmacol. 1999, 57, 1–7. [Google Scholar] [CrossRef]
- Kucharczak, J.; Simmons, M.J.; Fan, Y.; Gelinas, C. To be, or not to be: NF-kappaB is the answer--role of Rel/NF-kappaB in the regulation of apoptosis. Oncogene 2003, 22, 8961–8982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Methot, L.; Hermann, R.; Tang, Y.; Lo, R.; Al-Jehani, H.; Jhas, S.; Svoboda, D.; Slack, R.S.; Barker, P.A.; Stifani, S. Interaction and antagonistic roles of NF-kappaB and Hes6 in the regulation of cortical neurogenesis. Mol. Cell. Biol. 2013, 33, 2797–2808. [Google Scholar] [CrossRef] [Green Version]
- O’Loughlin, E.; Pakan, J.M.P.; Yilmazer-Hanke, D.; McDermott, K.W. Acute in utero exposure to lipopolysaccharide induces inflammation in the pre- and postnatal brain and alters the glial cytoarchitecture in the developing amygdala. J. Neuroinflammation 2017, 14, 212. [Google Scholar] [CrossRef]
- Wen, S.; Li, H.; Liu, J. Dynamic signaling for neural stem cell fate determination. Cell Adhes. Migr. 2009, 3, 107–117. [Google Scholar] [CrossRef] [Green Version]
- Fazio, C.; Ricciardiello, L. Inflammation and Notch signaling: A crosstalk with opposite effects on tumorigenesis. Cell Death Dis. 2016, 7, e2515. [Google Scholar] [CrossRef]
- Grivennikov, S.I.; Karin, M. Dangerous liaisons: STAT3 and NF-kappaB collaboration and crosstalk in cancer. Cytokine Growth Factor Rev. 2010, 21, 11–19. [Google Scholar] [CrossRef] [Green Version]
- van der Kraan, P.M.; Davidson, E.N. Cross-talk between bone morphogenetic proteins and inflammatory pathways. Arthritis Res. Ther. 2015, 17, 326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grandbarbe, L.; Bouissac, J.; Rand, M.; Hrabe de Angelis, M.; Artavanis-Tsakonas, S.; Mohier, E. Delta-Notch signaling controls the generation of neurons/glia from neural stem cells in a stepwise process. Development 2003, 130, 1391–1402. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Smyth, G.K. Linear models and empirical bayes methods for assessing differential expression in microarray experiments. Stat. Appl. Genet. Mol. Biol. 2004, 3. [Google Scholar] [CrossRef] [PubMed]
- Storey, J.D.; Tibshirani, R. Statistical significance for genomewide studies. Proc. Natl. Acad. Sci. USA 2003, 100, 9440–9445. [Google Scholar] [CrossRef] [Green Version]
- Pierce, J.W.; Schoenleber, R.; Jesmok, G.; Best, J.; Moore, S.A.; Collins, T.; Gerritsen, M.E. Novel inhibitors of cytokine-induced IkappaBalpha phosphorylation and endothelial cell adhesion molecule expression show anti-inflammatory effects in vivo. J. Biol. Chem. 1997, 272, 21096–21103. [Google Scholar] [CrossRef] [Green Version]
- Adam, S.A.; Schnell, O.; Poschl, J.; Eigenbrod, S.; Kretzschmar, H.A.; Tonn, J.C.; Schuller, U. ALDH1A1 is a marker of astrocytic differentiation during brain development and correlates with better survival in glioblastoma patients. Brain Pathol. 2012, 22, 788–797. [Google Scholar] [CrossRef] [PubMed]
- Pavlou, M.A.S.; Grandbarbe, L.; Buckley, N.J.; Niclou, S.P.; Michelucci, A. Transcriptional and epigenetic mechanisms underlying astrocyte identity. Prog. Neurobiol. 2019, 174, 36–52. [Google Scholar] [CrossRef] [PubMed]
- Park, K.M.; Bowers, W.J. Tumor necrosis factor-alpha mediated signaling in neuronal homeostasis and dysfunction. Cell. Signal. 2010, 22, 977–983. [Google Scholar] [CrossRef] [Green Version]
- Tak, P.P.; Gerlag, D.M.; Aupperle, K.R.; van de Geest, D.A.; Overbeek, M.; Bennett, B.L.; Boyle, D.L.; Manning, A.M.; Firestein, G.S. Inhibitor of nuclear factor kappaB kinase beta is a key regulator of synovial inflammation. Arthritis. Rheum. 2001, 44, 1897–1907. [Google Scholar] [CrossRef]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Hu, W. NFkappaB signaling regulates embryonic and adult neurogenesis. Front. Biol. 2012, 7. [Google Scholar] [CrossRef]
- O’Neill, L.A.; Kaltschmidt, C. NF-kappa B: A crucial transcription factor for glial and neuronal cell function. Trends Neurosci. 1997, 20, 252–258. [Google Scholar] [CrossRef]
- Widera, D.; Mikenberg, I.; Elvers, M.; Kaltschmidt, C.; Kaltschmidt, B. Tumor necrosis factor alpha triggers proliferation of adult neural stem cells via IKK/NF-kappaB signaling. BMC Neurosci. 2006, 7, 64. [Google Scholar] [CrossRef] [Green Version]
- Widera, D.; Kaus, A.; Kaltschmidt, C.; Kaltschmidt, B. Neural stem cells, inflammation and NF-kappaB: Basic principle of maintenance and repair or origin of brain tumours? J. Cell. Mol. Med. 2008, 12, 459–470. [Google Scholar] [CrossRef] [Green Version]
- Imielski, Y.; Schwamborn, J.C.; Luningschror, P.; Heimann, P.; Holzberg, M.; Werner, H.; Leske, O.; Puschel, A.W.; Memet, S.; Heumann, R.; et al. Regrowing the adult brain: NF-kappaB controls functional circuit formation and tissue homeostasis in the dentate gyrus. PLoS ONE 2012, 7, e30838. [Google Scholar] [CrossRef]
- Kaltschmidt, B.; Kaltschmidt, C. NF-KappaB in Long-Term Memory and Structural Plasticity in the Adult Mammalian Brain. Front. Mol. Neurosci. 2015, 8, 69. [Google Scholar] [CrossRef] [Green Version]
- Bortolotto, V.; Cuccurazzu, B.; Canonico, P.L.; Grilli, M. NF-kappaB mediated regulation of adult hippocampal neurogenesis: Relevance to mood disorders and antidepressant activity. Biomed. Res. Int. 2014, 2014, 612798. [Google Scholar] [CrossRef] [PubMed]
- Bonini, S.A.; Mastinu, A.; Maccarinelli, G.; Mitola, S.; Premoli, M.; La Rosa, L.R.; Ferrari-Toninelli, G.; Grilli, M.; Memo, M. Cortical Structure Alterations and Social Behavior Impairment in p50-Deficient Mice. Cereb. Cortex. 2016, 26, 2832–2849. [Google Scholar] [CrossRef]
- Ruiz-Perera, L.M.; Greiner, J.F.W.; Kaltschmidt, C.; Kaltschmidt, B. A Matter of Choice: Inhibition of c-Rel Shifts Neuronal to Oligodendroglial Fate in Human Stem Cells. Cells 2020, 9, 1037. [Google Scholar] [CrossRef] [PubMed]
- Crocker, S.J.; Frausto, R.F.; Whitton, J.L.; Milner, R. A novel method to establish microglia-free astrocyte cultures: Comparison of matrix metalloproteinase expression profiles in pure cultures of astrocytes and microglia. Glia 2008, 56, 1187–1198. [Google Scholar] [CrossRef] [Green Version]
- Brunet, J.F.; Grollimund, L.; Chatton, J.Y.; Lengacher, S.; Magistretti, P.J.; Villemure, J.G.; Pellerin, L. Early acquisition of typical metabolic features upon differentiation of mouse neural stem cells into astrocytes. Glia 2004, 46, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.M.; Kim, M.H.; Kim, B.H.; Jung, S.H.; Kim, Y.S.; Park, H.J.; Hong, J.T.; Min, K.R.; Kim, Y. Inhibitory action of novel aromatic diamine compound on lipopolysaccharide-induced nuclear translocation of NF-kappaB without affecting IkappaB degradation. FEBS Lett. 2004, 571, 50–54. [Google Scholar] [CrossRef] [Green Version]
- Kaltschmidt, B.; Kaltschmidt, C.; Hofmann, T.G.; Hehner, S.P.; Droge, W.; Schmitz, M.L. The pro- or anti-apoptotic function of NF-kappaB is determined by the nature of the apoptotic stimulus. Eur. J. Biochem. 2000, 267, 3828–3835. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Lee, H.; Bellas, R.E.; Schauer, S.L.; Arsura, M.; Katz, D.; FitzGerald, M.J.; Rothstein, T.L.; Sherr, D.H.; Sonenshein, G.E. Inhibition of NF-kappaB/Rel induces apoptosis of murine B cells. EMBO J. 1996, 15, 4682–4690. [Google Scholar] [CrossRef]
- Pizzi, M.; Sarnico, I.; Lanzillotta, A.; Battistin, L.; Spano, P. Post-ischemic brain damage: NF-kappaB dimer heterogeneity as a molecular determinant of neuron vulnerability. FEBS J. 2009, 276, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Sarnico, I.; Lanzillotta, A.; Benarese, M.; Alghisi, M.; Baiguera, C.; Battistin, L.; Spano, P.; Pizzi, M. NF-kappaB dimers in the regulation of neuronal survival. Int. Rev. Neurobiol. 2009, 85, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Bellavia, D.; Campese, A.F.; Alesse, E.; Vacca, A.; Felli, M.P.; Balestri, A.; Stoppacciaro, A.; Tiveron, C.; Tatangelo, L.; Giovarelli, M.; et al. Constitutive activation of NF-kappaB and T-cell leukemia/lymphoma in Notch3 transgenic mice. EMBO J. 2000, 19, 3337–3348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naruse, M.; Shibasaki, K.; Yokoyama, S.; Kurachi, M.; Ishizaki, Y. Dynamic changes of CD44 expression from progenitors to subpopulations of astrocytes and neurons in developing cerebellum. PLoS ONE 2013, 8, e53109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, A.; Kessler, J.D.; Read, T.A.; Kaiser, C.; Corbeil, D.; Huttner, W.B.; Johnson, J.E.; Wechsler-Reya, R.J. Isolation of neural stem cells from the postnatal cerebellum. Nat. Neurosci. 2005, 8, 723–729. [Google Scholar] [CrossRef] [Green Version]
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: Biology and pathology. Acta Neuropathol. 2010, 119, 7–35. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Liu, Y.; Levine, E.M.; Rao, M.S. Hes1 but not Hes5 regulates an astrocyte versus oligodendrocyte fate choice in glial restricted precursors. Dev. Dyn. 2003, 226, 675–689. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, C.; Blum, R.; Gascon, S.; Masserdotti, G.; Tripathi, P.; Sanchez, R.; Tiedt, S.; Schroeder, T.; Gotz, M.; Berninger, B. Directing astroglia from the cerebral cortex into subtype specific functional neurons. PLoS Biol. 2010, 8, e1000373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osipo, C.; Golde, T.E.; Osborne, B.A.; Miele, L.A. Off the beaten pathway: The complex cross talk between Notch and NF-kappaB. Lab. Investig. 2008, 88, 11–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maniati, E.; Bossard, M.; Cook, N.; Candido, J.B.; Emami-Shahri, N.; Nedospasov, S.A.; Balkwill, F.R.; Tuveson, D.A.; Hagemann, T. Crosstalk between the canonical NF-kappaB and Notch signaling pathways inhibits Ppargamma expression and promotes pancreatic cancer progression in mice. J. Clin. Investig. 2011, 121, 4685–4699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urayama, S.; Semi, K.; Sanosaka, T.; Hori, Y.; Namihira, M.; Kohyama, J.; Takizawa, T.; Nakashima, K. Chromatin accessibility at a STAT3 target site is altered prior to astrocyte differentiation. Cell. Struct. Funct. 2013, 38, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Mao, R.; Yang, J. NF-kappaB and STAT3 signaling pathways collaboratively link inflammation to cancer. Protein Cell. 2013, 4, 176–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molofsky, A.V.; Glasgow, S.M.; Chaboub, L.S.; Tsai, H.H.; Murnen, A.T.; Kelley, K.W.; Fancy, S.P.; Yuen, T.J.; Madireddy, L.; Baranzini, S.; et al. Expression profiling of Aldh1l1-precursors in the developing spinal cord reveals glial lineage-specific genes and direct Sox9-Nfe2l1 interactions. Glia 2013, 61, 1518–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonaguidi, M.A.; McGuire, T.; Hu, M.; Kan, L.; Samanta, J.; Kessler, J.A. LIF and BMP signaling generate separate and discrete types of GFAP-expressing cells. Development 2005, 132, 5503–5514. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse |
|---|---|---|
| Casp3 | GGCATTGAGACAGACAGT | GTAGAGTAAGCATACAGGAAGT |
| Cd133 | ACCAGCGGCAGAAGCAGAATG | TGAGCAGACAAATCACCAGGAGAG |
| Cd44 | TGGCACTGGCTCTGATTC | GTCTCTGATGGTTCCTTGTTC |
| Gfap | GGTTGAATCGCTGGAGGAG | CTGTGAGGTCTGGCTTGG |
| Hes1 | GCCAATTTGCCTTTCTCATCC | GGTGACACTGCGTTAGGAC |
| Hes5 | CGGTGGTGGAGAAGATGC | CTTGGAGTTGGGCTGGTG |
| Id1 | TGCTGCCCTGATTATGA | GAAAGTCACCTTCCTGTAAA |
| Id3 | ATGAACGGCTGCTACTC | CTCCACCTTGCTCACTT |
| IκBα | GCCAGTGTAGCAGTCTTGAC | GCCAGGTAGCCGTGAGTAG |
| Mash1 | AAGATGAGCAAGGTGGAG | AGTCGTTGGAGTAGTTGG |
| MKi67 | TTCCTTCAGCAAGCCTGAG | GTATTAGGAGGCAAGTT |
| NeuroD1 | GAACTACATCTGGGCTCTG | GAAAGTCCGAGGGTTGAG |
| Socs3 | ACCCTCCACAT-CTTTGTC | TCATACTGATCCAGGAACTCC |
| β-Actin | AGGGAAATCGTGCGTGACATCAAAGAG | GGAGGAAGAGGATGCGGAGTGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Birck, C.; Ginolhac, A.; Pavlou, M.A.S.; Michelucci, A.; Heuschling, P.; Grandbarbe, L. NF-κB and TNF Affect the Astrocytic Differentiation from Neural Stem Cells. Cells 2021, 10, 840. https://doi.org/10.3390/cells10040840
Birck C, Ginolhac A, Pavlou MAS, Michelucci A, Heuschling P, Grandbarbe L. NF-κB and TNF Affect the Astrocytic Differentiation from Neural Stem Cells. Cells. 2021; 10(4):840. https://doi.org/10.3390/cells10040840
Chicago/Turabian StyleBirck, Cindy, Aurélien Ginolhac, Maria Angeliki S. Pavlou, Alessandro Michelucci, Paul Heuschling, and Luc Grandbarbe. 2021. "NF-κB and TNF Affect the Astrocytic Differentiation from Neural Stem Cells" Cells 10, no. 4: 840. https://doi.org/10.3390/cells10040840
APA StyleBirck, C., Ginolhac, A., Pavlou, M. A. S., Michelucci, A., Heuschling, P., & Grandbarbe, L. (2021). NF-κB and TNF Affect the Astrocytic Differentiation from Neural Stem Cells. Cells, 10(4), 840. https://doi.org/10.3390/cells10040840