
Periostin Short Fragment with Exon 17 via Aberrant Alternative Splicing Is Required for Breast Cancer Growth and Metastasis


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Cell Lines and Reagents
2.3. Antibodies against POSTN
2.4. Dot Blotting
2.5. Immunohistological Assay
2.6. Western Blotting
2.7. Immunoprecipitation
2.8. Liquid Chromatography–Tandem Mass Spectrometry (LC–MS/MS) and Data Analysis
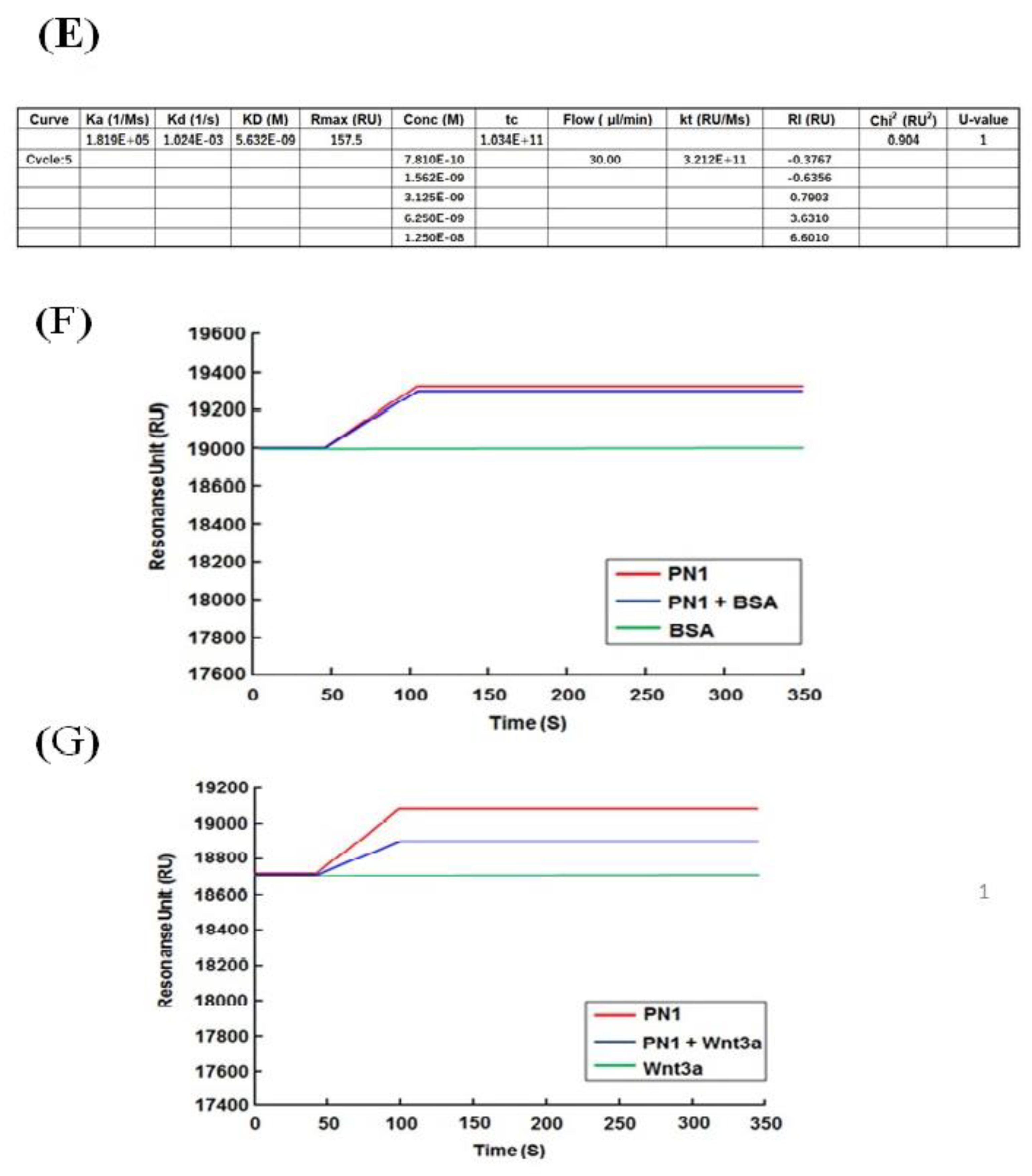
2.9. Surface Plasmon Resonance Analysis
2.10. Total RNA Extraction via Quantitative Real-Time PCR
2.11. Statistical Analysis
3. Results
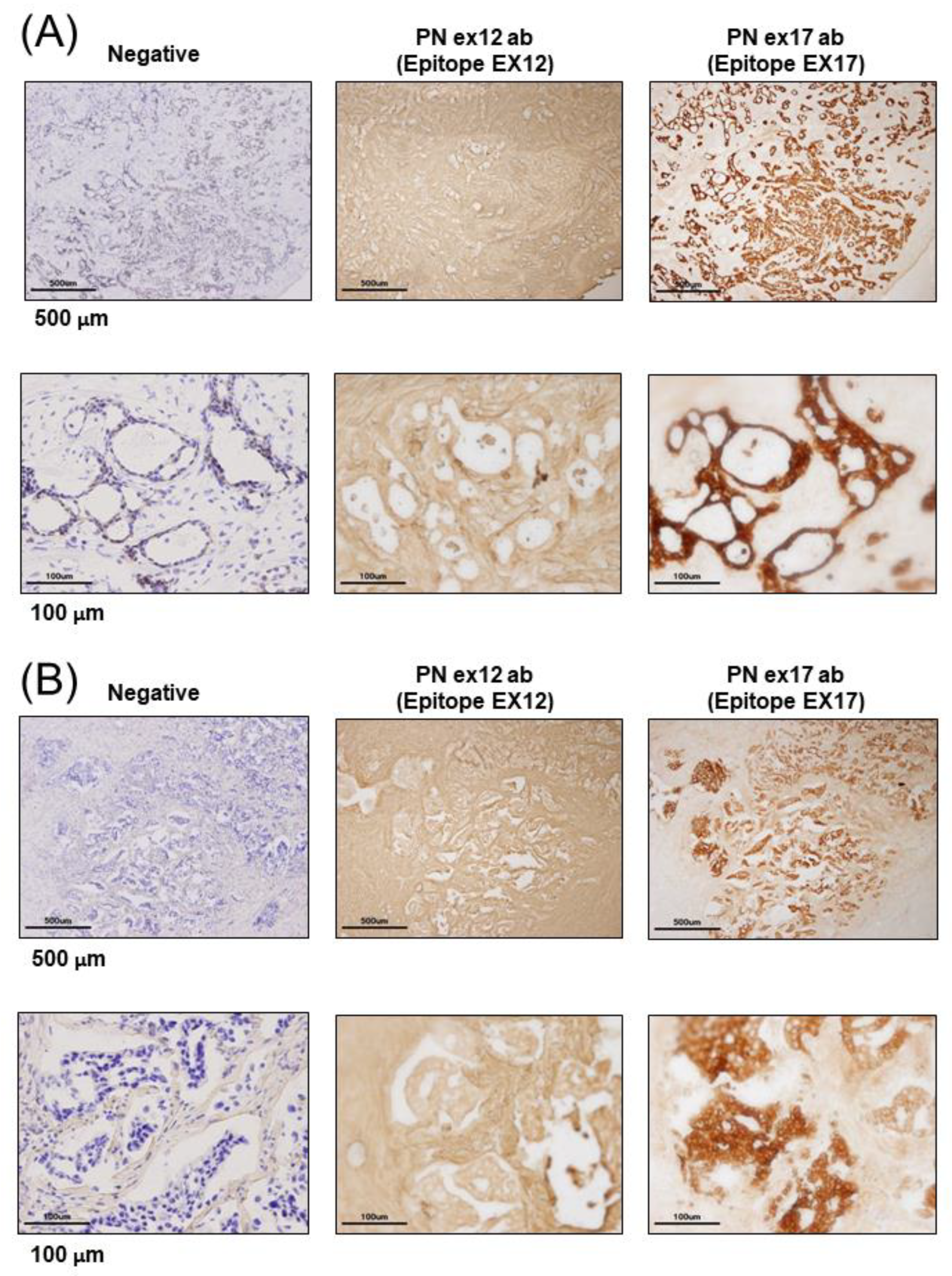
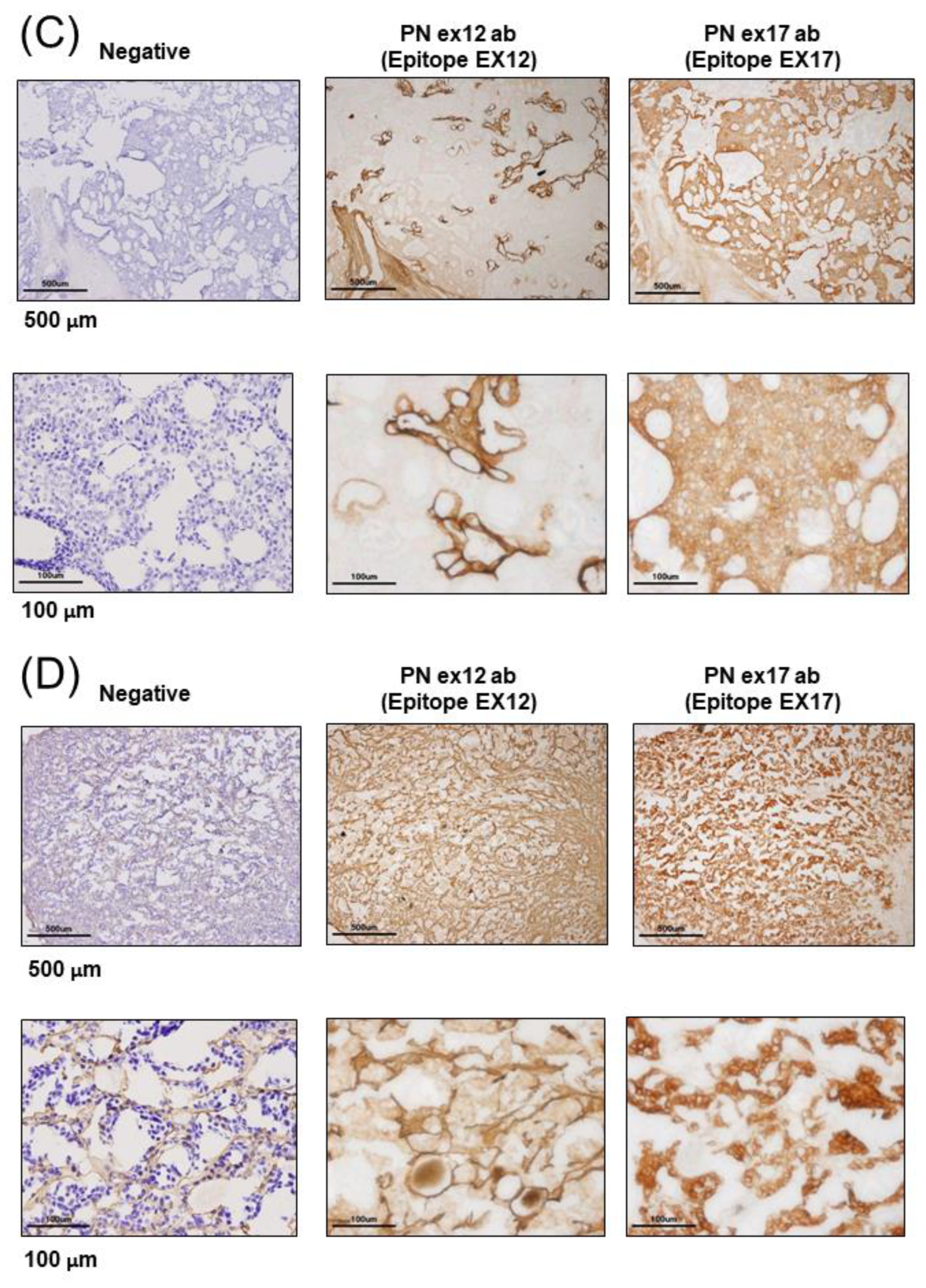
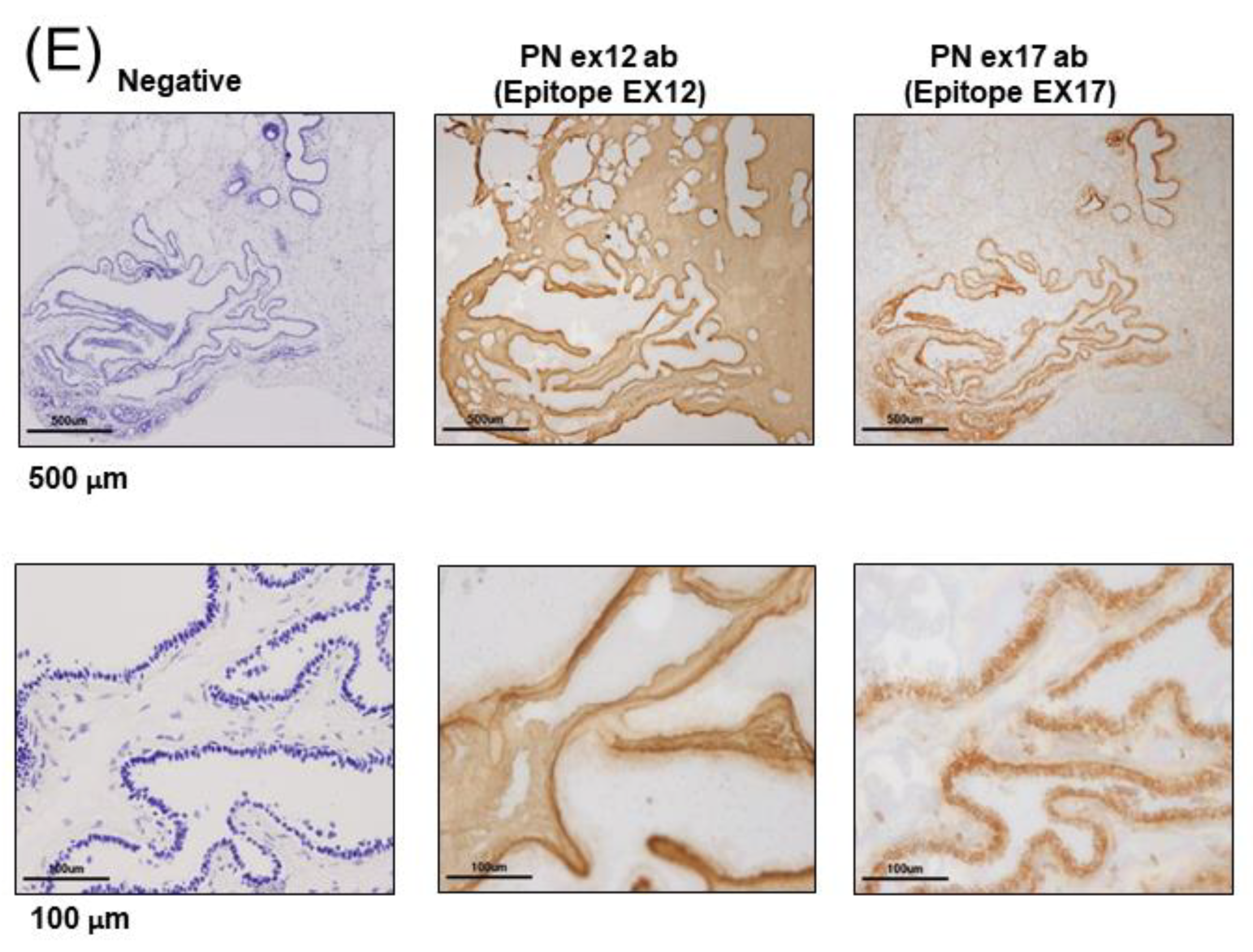
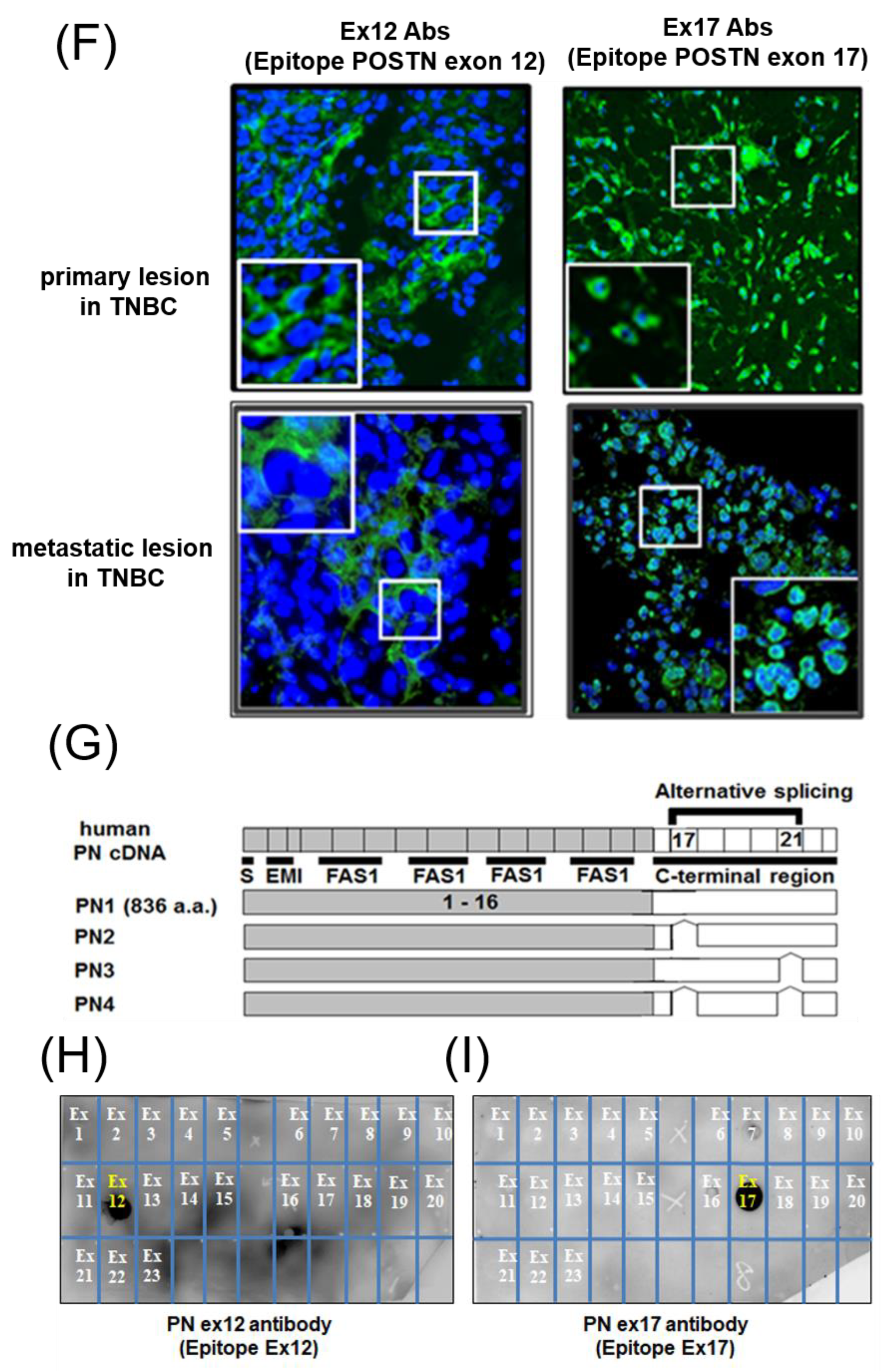
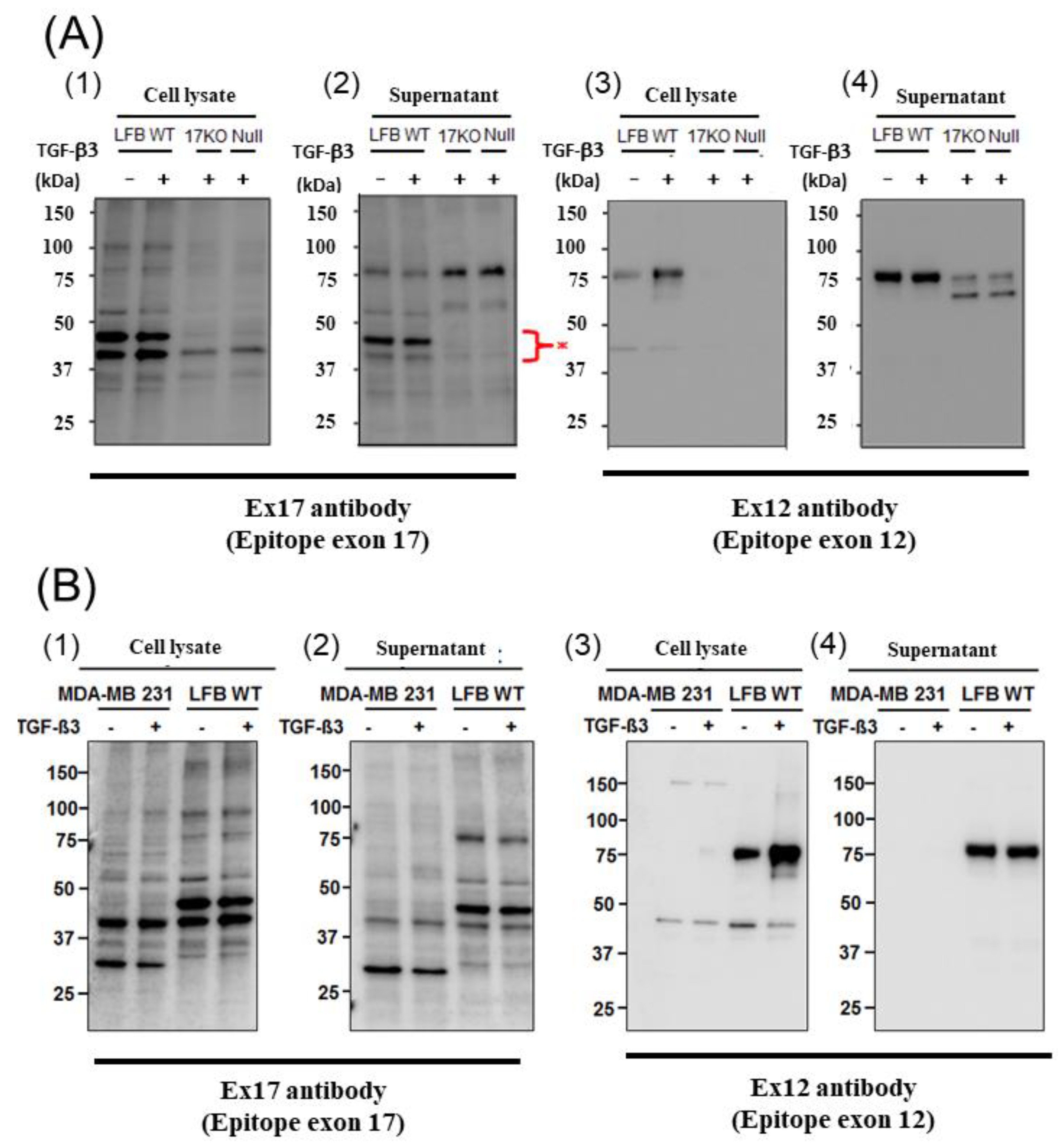
3.1. POSTN Exon 17 Is Associated with Breast Cancer Cells
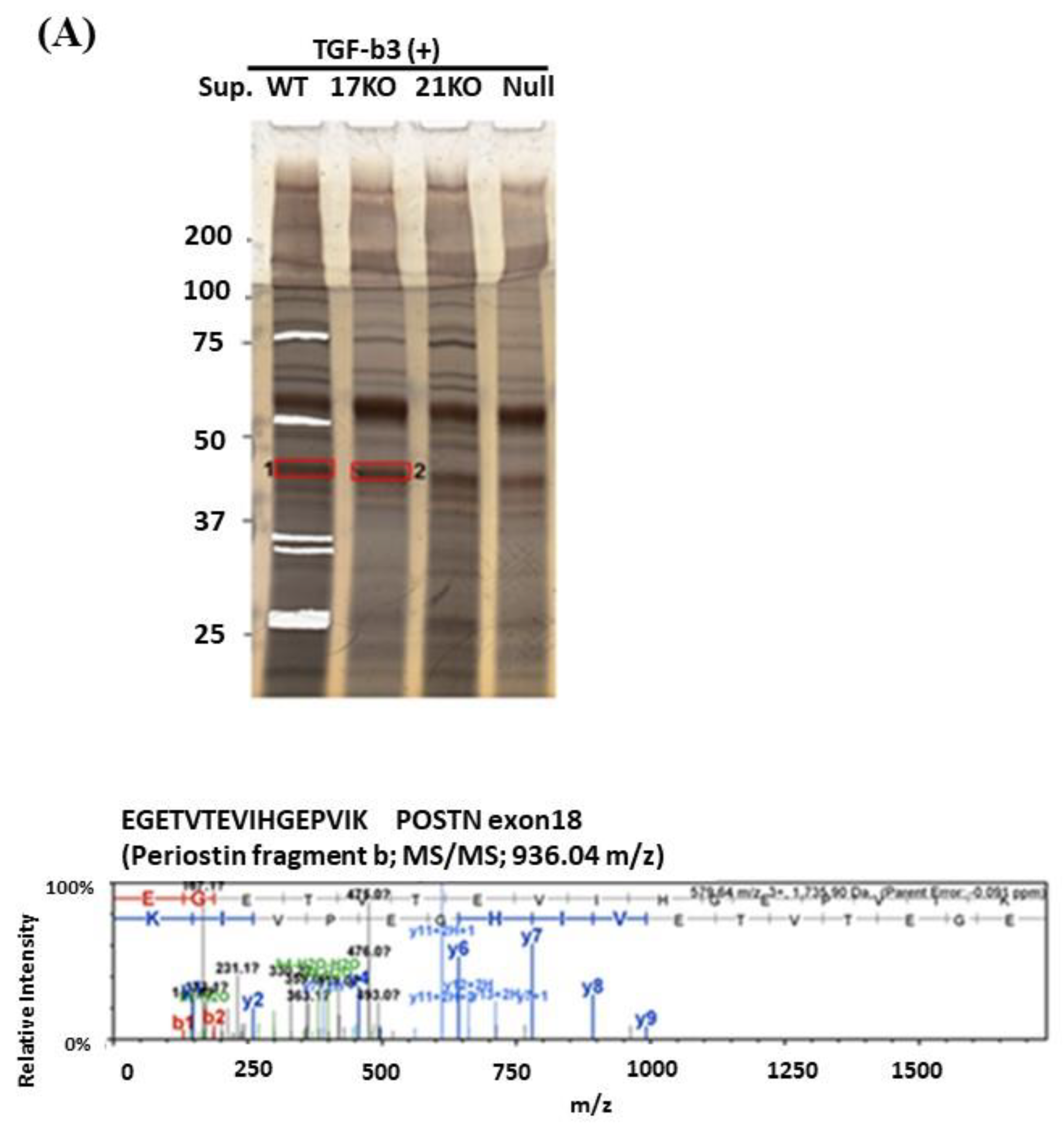
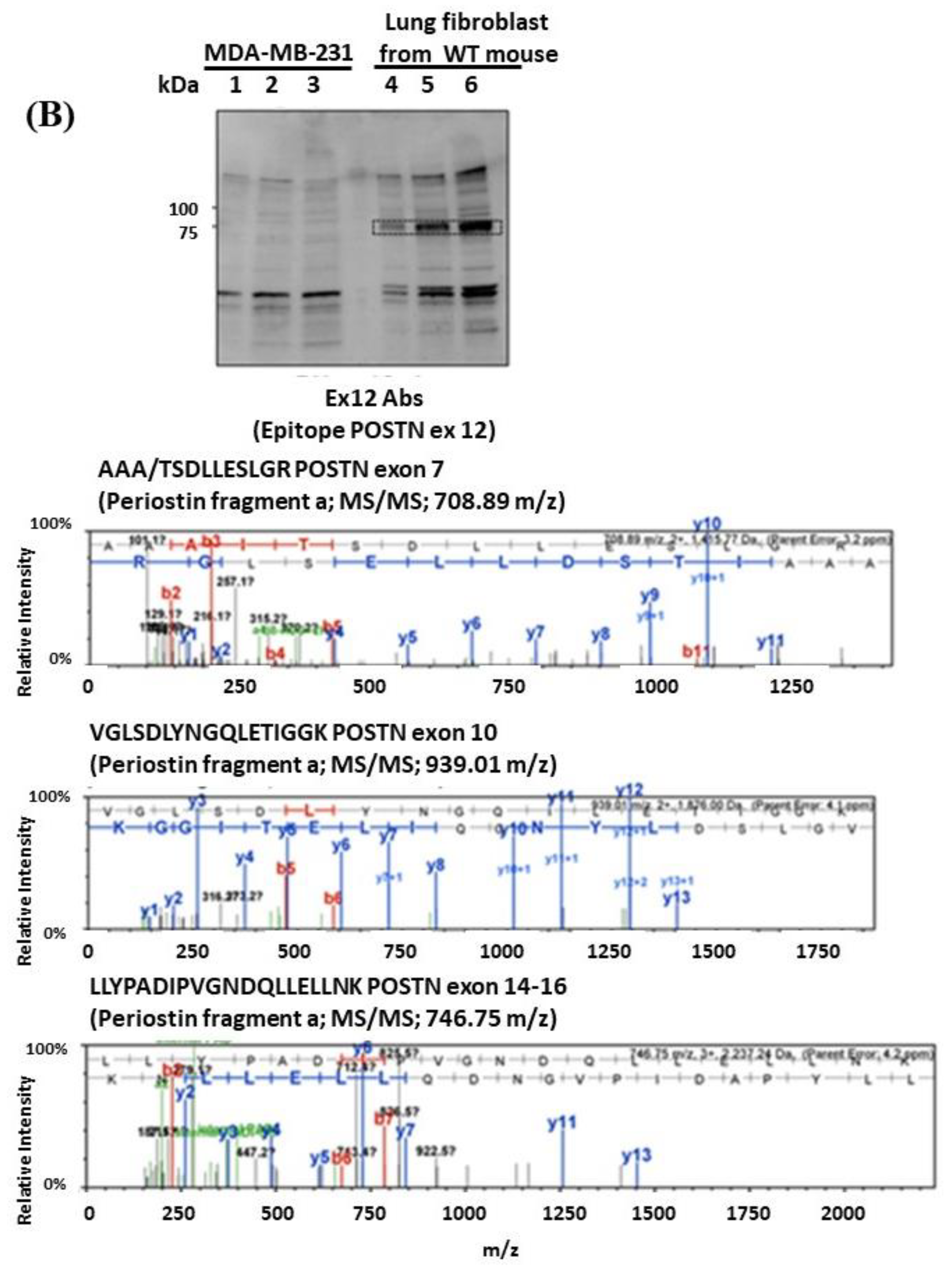
3.2. Proteomic Approach for the Identification of Short Fragments of POSTN
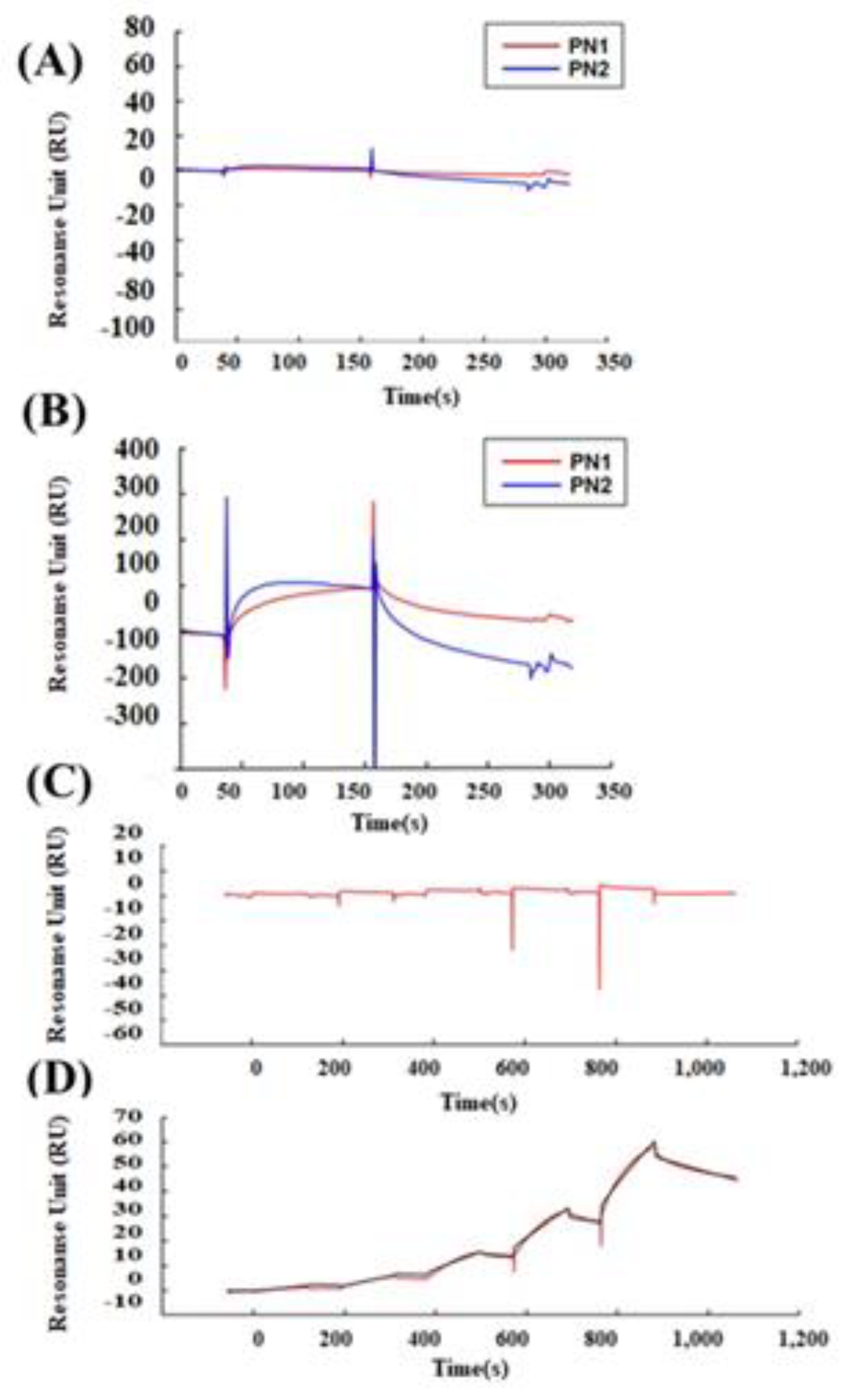
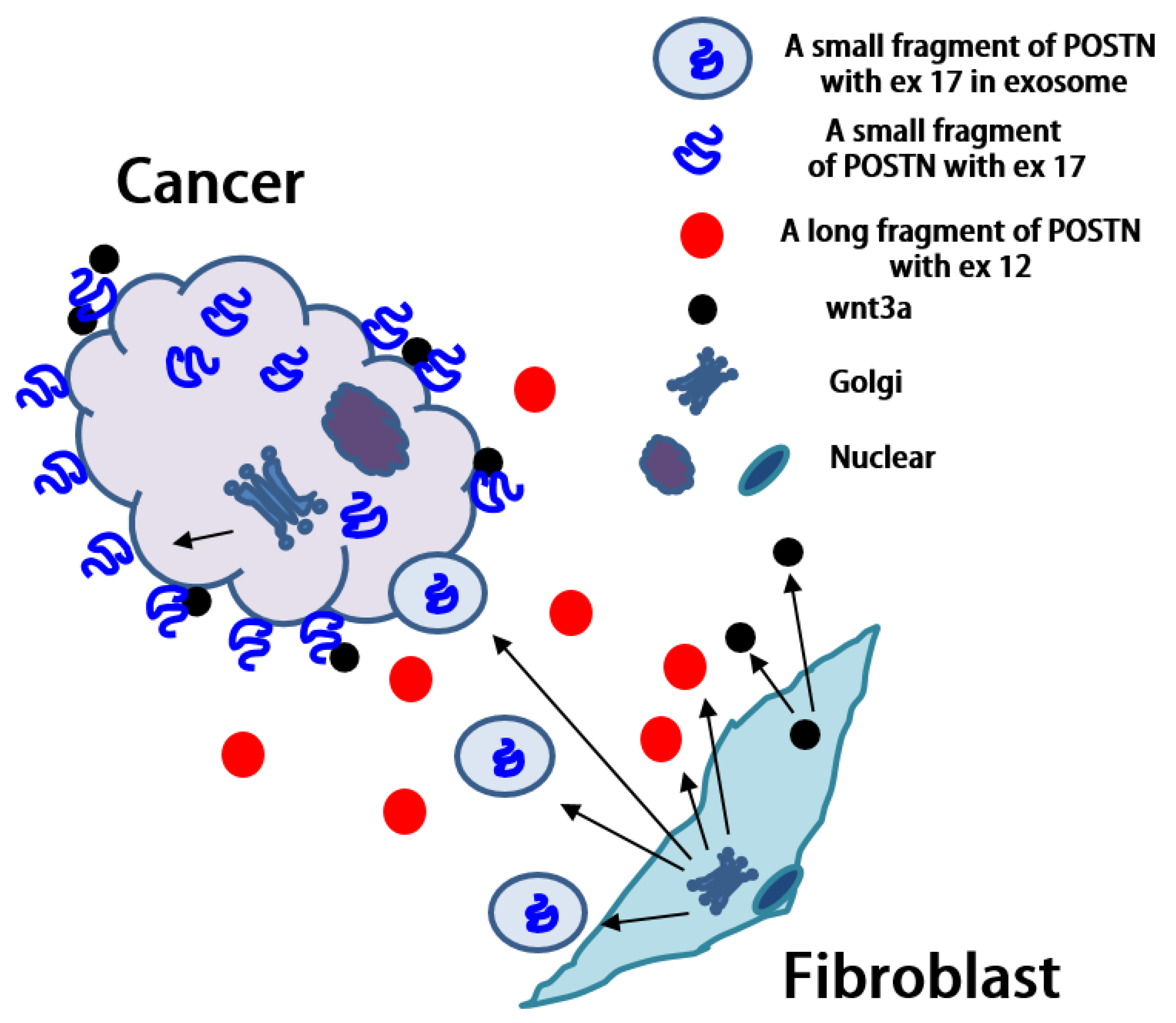
3.3. POSTN Exon 17 Region Binds to Wnt3a
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| POSTN | Periostin |
| TGF- | transforming growth factor- |
| NOG mice | NOD/Shi-scid, IL-2RγKO mice |
| KO | konck out |
| OCT | optical coherence tomography |
| Ex | exon |
| KLH | keyhole limpet hemocyanin |
| HRP | horseradish peroxidase |
| DAB | 3,3′-Diaminobenzidine |
| Ex17 antibody | antibodies against POSTN C-terminal region exon 17 |
| Ex12 antibody | antibodies against POSTN N-terminal region exon 12 |
| DMEM | Dulbecco’s modified Eagle’s minimum essential medium |
| DAPI | 4′,6-diamidino-2-phenylindole |
| CLB | cell lysis buffer |
| EDTA | ethylenediaminetetraacetic acid |
| QqTOF | quadrupole time-of flight |
| SA | streptavidin |
| BSA | bovine serum albumin |
| TNBC | triple negative breast cancer cells |
| ECM | extracellular matrix |
| TGFBI | IgH3 |
| BMP 2 | bone morphogenic protein-2 |
| FBS | fetal bovine serum |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| FAS1 | fasciclin 1 |
| PN1 | full-length POSTN |
| PN2 | POSTN lack of exon 17 |
| PN3 | POSTN lack of exon 21 |
| PN4 | POSTN lack of exons 17 and 21 |
| PCR | polymerase chain reaction |
| EMI | EMILIN-1 |
| MMPs | Matrix metalloproteinases |
References
- Kotobuki, Y.; Yang, L.; Serada, S.; Tanemura, A.; Yang, F.; Nomura, S.; Kudo, A.; Izuhara, K.; Murota, H.; Fujimoto, M.; et al. Periostin accelerates human malignant melanoma progression by modifying the melanoma microenvironment. Pigment. Cell Melanoma Res. 2014, 27, 630–639. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yuan, D.; Yao, Y.; Sun, W.; Shi, Y.; Su, X. Predictive and prognostic value of serum periostin in ad-vanced non-small cell lung cancer patients receiving chemotherapy. Tumour Biol. 2017, 39. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Chang, W.; Yuan, J.; Han, X.; Tan, X.; Ding, Y.; Luo, Y.; Cai, H.; Liu, Y.; Gao, X.; et al. Periostin expression in intra-tumoral stromal cells is prognostic and predictive for colorectal carcinoma via creating a cancer-supportive niche. Oncotarget 2016, 7, 798–813. [Google Scholar] [CrossRef] [Green Version]
- Kanno, A.; Satoh, K.; Masamune, A.; Hirota, M.; Kimura, K.; Umino, J.; Hamada, S.; Satoh, A.; Egawa, S.; Motoi, F.; et al. Periostin, secreted from stromal cells, has biphasic effect on cell migration and correlates with the epithelial to mesenchymal transition of human pancreatic cancer cells. Int. J. Cancer 2008, 122, 2707–2718. [Google Scholar] [CrossRef] [PubMed]
- Underwood, T.J.; Hayden, A.L.; Derouet, M.; Garcia, E.; Noble, F.; White, M.J.; Thirdborough, S.; Mead, A.; Clemons, N.; Mellone, M.; et al. Cancer-associated fibroblasts predict poor outcome and promote periostin-dependent invasion in oesophageal adenocarcinoma. J. Pathol. 2015, 235, 466–477. [Google Scholar] [CrossRef]
- Sung, P.-L.; Jan, Y.-H.; Lin, S.-C.; Huang, C.-C.; Lin, H.; Wen, K.-C.; Chao, K.-C.; Lai, C.-R.; Wang, P.-H.; Wu, H.-H.; et al. Periostin in tumor microenvironment is associated with poor prognosis and platinum resistance in epithelial ovarian carcinoma. Oncotarget 2015, 7, 4036–4047. [Google Scholar] [CrossRef] [Green Version]
- Malanchi, I.; Santamaria-Martínez, A.; Susanto, E.; Peng, H.; Lehr, H.-A.; Delaloye, J.-F.; Huelsken, J. Interactions between cancer stem cells and their niche govern metastatic colonization. Nature 2011, 481, 85–89. [Google Scholar] [CrossRef]
- Ghajar, C.M.; Peinado, H.; Mori, H.; Matei, I.R.; Evason, K.J.; Brazier, H.; de Almeida, D.L.; Koller, A.; Hajjar, K.A.; Stainier, D.Y.R.; et al. The perivascular niche regulates breast tumour dormancy. Nat. Cell Biol. 2013, 15, 807–817. [Google Scholar] [CrossRef]
- Dass, S.; Tan, K.; Rajan, R.S.; Mokhtar, N.; Adzmi, E.M.; Rahman, W.W.A.; Din, T.T.; Balakrishnan, V. Triple Negative Breast Cancer: A Review of Present and Future Diagnostic Modalities. Medicina 2021, 57, 62. [Google Scholar] [CrossRef]
- Dawson, S.-J.; Tsui, D.W.; Murtaza, M.; Biggs, H.; Rueda, O.M.; Chin, S.-F.; Dunning, M.J.; Gale, D.; Forshew, T.; Mahler-Araujo, B.; et al. Analysis of Circulating Tumor DNA to Monitor Metastatic Breast Cancer. N. Engl. J. Med. 2013, 368, 1199–1209. [Google Scholar] [CrossRef] [Green Version]
- Horiuchi, H.; Amizuka, N.; Takeshita, S.; Takamatsu, H.; Katsuura, M.; Ozawa, H.; Toyama, Y.; Bonewald, L.F.; Kudo, A. Identification and characterization of a novel protein, periostin, with restricted expression to periosteum and periodontal ligament and increased expression by transforming growth factor beta. J. Bone Miner Res. 1999, 14, 1239–1249. [Google Scholar] [CrossRef]
- Hoersch, S.; Andrade-Navarro, M.A. Periostin shows increased evolutionary plasticity in its alternatively spliced region. BMC Evol. Biol. 2010, 10, 30. [Google Scholar] [CrossRef] [Green Version]
- Frerker, N.; Kasprzycka, M.; Mikalsen, B.; Line, P.D.; Scott, H.; Dept, G.H. Role of Matricellular Proteins in Cardiac Allograft Fibrosis. Card. Transplant. 2012, 6, 99–122. [Google Scholar]
- Chijimatsu, R.; Kunugiza, Y.; Taniyama, Y.; Nakamura, N.; Tomita, T.; Yoshikawa, H. Expression and pathological effects of periostin in human osteoarthritis cartilage. BMC Musculoskelet. Disord. 2015, 16, 215. [Google Scholar] [CrossRef] [Green Version]
- Nakama, T.; Yoshida, S.; Ishikawa, K.; Kobayashi, Y.; Abe, T.; Kiyonari, H.; Shioi, G.; Katsuragi, N.; Ishibashi, T.; Morishita, R.; et al. Different roles played by periostin splice variants in retinal neovascularization. Exp. Eye Res. 2016, 153, 133–140. [Google Scholar] [CrossRef]
- Gao, H.; Chakraborty, G.; Lee-Lim, A.P.; Mo, Q.; Decker, M.; Vonica, A.; Shen, R.; Brogi, E.; Brivanlou, A.H.; Giancotti, F.G. The BMP Inhibitor Coco Reactivates Breast Cancer Cells at Lung Metastatic Sites. Cell 2012, 150, 764–779. [Google Scholar] [CrossRef] [Green Version]
- Masuoka, M.; Shiraishi, H.; Ohta, S.; Suzuki, S.; Arima, K.; Aoki, S.; Toda, S.; Inagaki, N.; Kurihara, Y.; Hayashida, S.; et al. Periostin promotes chronic allergic inflammation in response to Th2 cytokines. J. Clin. Investig. 2012, 122, 2590–2600. [Google Scholar] [CrossRef] [Green Version]
- Gillan, L.; Matei, D.; Fishman, D.A.; Gerbin, C.S.; Karlan, B.Y.; Chang, D.D. Periostin secreted by epithelial ovarian carcinoma is a ligand for alpha(V)beta(3) and alpha(V)beta(5) integrins and promotes cell motility. Cancer Res. 2002, 62, 5358–5364. [Google Scholar]
- Planche, A.; Bacac, M.; Provero, P.; Fusco, C.; Delorenzi, M.; Stehle, J.C.; Stamenkovic, I. Identification of prog-nostic molecular features in the reactive stroma of human breast and prostate cancer. PLoS ONE 2011, 6, e18640. [Google Scholar] [CrossRef]
- Xu, D.; Xu, H.; Ren, Y.; Liu, C.; Wang, X.; Zhang, H.; Lu, P. Cancer Stem Cell-Related Gene Periostin: A Novel Prognostic Marker for Breast Cancer. PLoS ONE 2012, 7, e46670. [Google Scholar] [CrossRef]
- Ratajczak-Wielgomas, K.; Grzegrzolka, J.; Piotrowska, A.; Matkowski, R.; Wojnar, A.; Rys, J.; Ugorski, M.; Dziegiel, P. Expression of periostin in breast cancer cells. Int. J. Oncol. 2017, 51, 1300–1310. [Google Scholar] [CrossRef]
- Wang, X.; Liu, J.; Wang, Z.; Huang, Y.; Liu, W.; Zhu, X.; Cai, Y.; Fang, X.; Lin, S.; Yuan, L.; et al. Periostin Contributes to the Acquisition of Multipotent Stem Cell-Like Properties in Human Mammary Epithelial Cells and Breast Cancer Cells. PLoS ONE 2013, 8, e72962. [Google Scholar] [CrossRef] [Green Version]
- Kyutoku, M.; Taniyama, Y.; Katsuragi, N.; Shimizu, H.; Kunugiza, Y.; Iekushi, K.; Koibuchi, N.; Sanada, F.; Oshita, Y.; Morishita, R. Role of periostin in cancer progression and metastasis: Inhibition of breast cancer progression and metastasis by antiperiostin antibody in a murine model. Int. J. Mol. Med. 2011, 28, 181–186. [Google Scholar]
- Katsuragi, N.; Morishita, R.; Nakamura, N.; Ochiai, T.; Taniyama, Y.; Hasegawa, Y.; Kawashima, K.; Kaneda, Y.; Ogihara, T.; Sugimura, K. Periostin as a Novel Factor Responsible for Ventricular Dilation. Circulation 2004, 110, 1806–1813. [Google Scholar] [CrossRef] [Green Version]
- Taniyama, Y.; Katsuragi, N.; Sanada, F.; Azuma, J.; Iekushi, K.; Koibuchi, N.; Okayama, K.; Ikeda-Iwabu, Y.; Muratsu, J.; Otsu, R.; et al. Selective Blockade of Periostin Exon 17 Preserves Cardiac Performance in Acute Myocardial Infarction. Hypertension 2016, 67, 356–361. [Google Scholar] [CrossRef]
- Iekushi, K.; Taniyama, Y.; Azuma, J.; Katsuragi, N.; Dosaka, N.; Sanada, F.; Koibuchi, N.; Nagao, K.; Ogihara, T.; Morishita, R. Novel Mechanisms of Valsartan on the Treatment of Acute Myocardial Infarction Through Inhibition of the Antiadhesion Molecule Periostin. Hypertension 2007, 49, 1409–1414. [Google Scholar] [CrossRef] [Green Version]
- Shimamura, M.; Taniyama, Y.; Katsuragi, N.; Koibuchi, N.; Kyutoku, M.; Sato, N.; Allahtavakoli, M.; Wakayama, K.; Nakagami, H.; Morishita, R. Role of Central Nervous System Periostin in Cerebral Ischemia. Stroke 2012, 43, 1108–1114. [Google Scholar] [CrossRef] [Green Version]
- Shimamura, M.; Taniyama, Y.; Nakagami, H.; Katsuragi, N.; Wakayama, K.; Koriyama, H.; Kurinami, H.; Tenma, A.; Tomioka, H.; Morishita, R. Long-term expression of periostin during the chronic stage of ischemic stroke in mice. Hypertens. Res. 2014, 37, 494–499. [Google Scholar] [CrossRef] [PubMed]
- Okada, T.; Ohama, T.; Takafuji, K.; Kanno, K.; Matsuda, H.; Sairyo, M.; Zhu, Y.; Saga, A.; Kobayashi, T.; Masuda, D.; et al. Shotgun proteomic analysis reveals proteome al-terations in HDL of patients with cholesteryl ester transfer protein deficiency. J. Clin. Lipidol. 2019, 13, 317–325. [Google Scholar] [CrossRef]
- Shao, R.; Bao, S.; Bai, X.; Blanchette, C.; Anderson, R.M.; Dang, T.; Gishizky, M.L.; Marks, J.R.; Wang, X.-F. Acquired Expression of Periostin by Human Breast Cancers Promotes Tumor Angiogenesis through Up-Regulation of Vascular Endothelial Growth Factor Receptor 2 Expression. Mol. Cell. Biol. 2004, 24, 3992–4003. [Google Scholar] [CrossRef] [Green Version]
- Vardaki, I.; Ceder, S.; Rutishauser, D.; Baltatzis, G.; Foukakis, T.; Panaretakis, T. Periostin is identified as a putative metastatic marker in breast cancer-derived exosomes. Oncotarget 2016, 7, 74966–74978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luga, V.; Zhang, L.; Viloria-Petit, A.M.; Ogunjimi, A.A.; Inanlou, M.R.; Chiu, E.; Buchanan, M.; Hosein, A.N.; Basik, M.; Wrana, J.L. Exosomes Mediate Stromal Mobilization of Autocrine Wnt-PCP Signaling in Breast Cancer Cell Migration. Cell 2012, 151, 1542–1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Underhaug, J.; Koldsø, H.; Runager, K.; Nielsen, J.T.; Sørensen, C.S.; Kristensen, T.; Otzen, D.E.; Karring, H.; Malmendal, A.; Schiøtt, B.; et al. Mutation in Transforming Growth Factor Beta Induced protein associated with Granular Corneal Dystrophy Type 1 Reduces the Proteolytic Susceptibility through Local Structural Stabilization#. Biochim. Biophys. Acta Proteins Proteom. 2013, 1834, 2812–2822. [Google Scholar] [CrossRef] [Green Version]
- Kudo, A. Periostin in fibrillogenesis for tissue regeneration: Periostin actions inside and outside the cell. Cell. Mol. Life Sci. 2011, 68, 3201–3207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Y.; Chen, Y.; Wang, Q.; Jayasinghe, U.; Luo, X.; Wei, Q.; Wang, J.; Xiong, H.; Chen, C.; Xu, B.; et al. Exosome: Emerging biomarker in breast cancer. Oncotarget 2017, 8, 41717–41733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakane, H.; Yamamoto, H.; Matsumoto, S.; Sato, A.; Kikuchi, A. Localization of glypican-4 in different membrane microdomains is involved in the regulation of Wnt signaling. J. Cell Sci. 2012, 125, 449–460. [Google Scholar] [CrossRef] [Green Version]
- Nadanaka, S.; Ishida, M.; Ikegami, M.; Kitagawa, H. Chondroitin 4-O-Sulfotransferase-1 Modulates Wnt-3a Signaling through Control of E Disaccharide Expression of Chondroitin Sulfate. J. Biol. Chem. 2008, 283, 27333–27343. [Google Scholar] [CrossRef] [Green Version]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ikeda-Iwabu, Y.; Taniyama, Y.; Katsuragi, N.; Sanada, F.; Koibuchi, N.; Shibata, K.; Shimazu, K.; Rakugi, H.; Morishita, R. Periostin Short Fragment with Exon 17 via Aberrant Alternative Splicing Is Required for Breast Cancer Growth and Metastasis. Cells 2021, 10, 892. https://doi.org/10.3390/cells10040892
Ikeda-Iwabu Y, Taniyama Y, Katsuragi N, Sanada F, Koibuchi N, Shibata K, Shimazu K, Rakugi H, Morishita R. Periostin Short Fragment with Exon 17 via Aberrant Alternative Splicing Is Required for Breast Cancer Growth and Metastasis. Cells. 2021; 10(4):892. https://doi.org/10.3390/cells10040892
Chicago/Turabian StyleIkeda-Iwabu, Yuka, Yoshiaki Taniyama, Naruto Katsuragi, Fumihiro Sanada, Nobutaka Koibuchi, Kana Shibata, Kenzo Shimazu, Hiromi Rakugi, and Ryuichi Morishita. 2021. "Periostin Short Fragment with Exon 17 via Aberrant Alternative Splicing Is Required for Breast Cancer Growth and Metastasis" Cells 10, no. 4: 892. https://doi.org/10.3390/cells10040892
APA StyleIkeda-Iwabu, Y., Taniyama, Y., Katsuragi, N., Sanada, F., Koibuchi, N., Shibata, K., Shimazu, K., Rakugi, H., & Morishita, R. (2021). Periostin Short Fragment with Exon 17 via Aberrant Alternative Splicing Is Required for Breast Cancer Growth and Metastasis. Cells, 10(4), 892. https://doi.org/10.3390/cells10040892