
Exercise Training-Induced Extracellular Matrix Protein Adaptation in Locomotor Muscles: A Systematic Review

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Search Strategy
2.2. Study Selection and Data Extraction
2.3. Quality Assessment
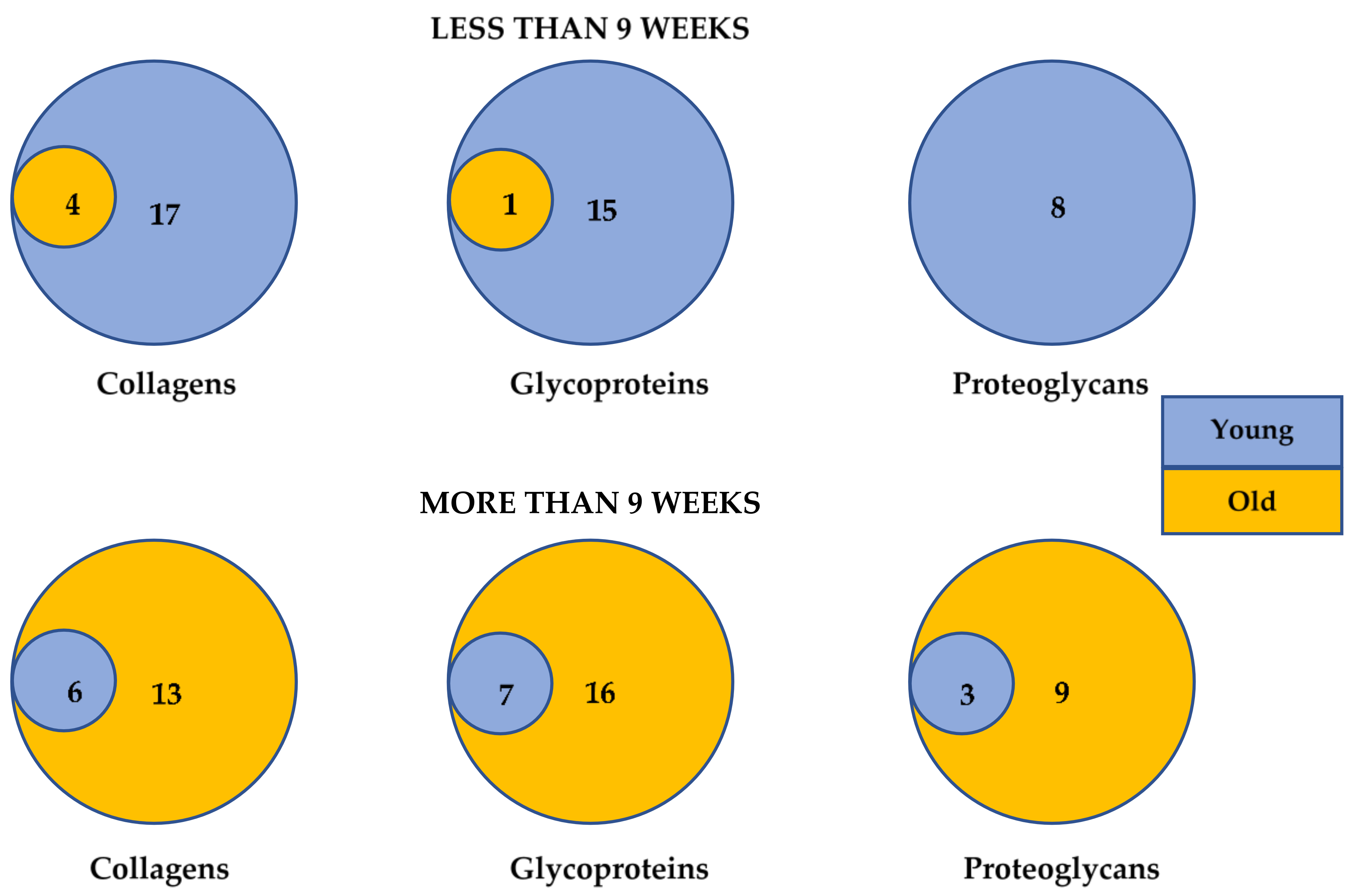
3. Results
3.1. Analysis of Risk of Bias
3.2. Study and Subject Characteristics
3.3. Exercise Training Increases the Expression of ECMs Associated with Skeletal Muscle Remodelling
3.4. ECM Adaptation Associated with Muscle Structure and Stability
3.5. ECMs Associated with Myogenic Regeneration and Repair
3.6. Angiogenesis
4. Discussion
Strength and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Neufer, P.D.; Bamman, M.M.; Muoio, D.M.; Bouchard, C.; Cooper, D.M.; Goodpaster, B.H.; Booth, F.W.; Kohrt, W.M.; Gerszten, R.E.; Mattson, M.P.; et al. Understanding the Cellular and Molecular Mechanisms of Physical Activity-Induced Health Benefits. Cell Metab. 2015, 22, 4–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz-Jentoft, A.J.; Sayer, A.A. Sarcopenia. Lancet 2019, 393, 2636–2646. [Google Scholar] [CrossRef]
- Sepulveda-Loyola, W.; Osadnik, C.; Phu, S.; Morita, A.A.; Duque, G.; Probst, V.S. Diagnosis, prevalence, and clinical impact of sarcopenia in COPD: A systematic review and meta-analysis. J. Cachexia Sarcopenia Muscle 2020, 11, 1164–1176. [Google Scholar] [CrossRef] [PubMed]
- Vogiatzis, I.; Stratakos, G.; Simoes, D.C.; Terzis, G.; Georgiadou, O.; Roussos, C.; Zakynthinos, S. Effects of rehabilitative exercise on peripheral muscle TNFalpha, IL-6, IGF-I and MyoD expression in patients with COPD. Thorax 2007, 62, 950–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogiatzis, I.; Terzis, G.; Nanas, S.; Stratakos, G.; Simoes, D.C.; Georgiadou, O.; Zakynthinos, S.; Roussos, C. Skeletal muscle adaptations to interval training in patients with advanced COPD. Chest 2005, 128, 3838–3845. [Google Scholar] [CrossRef] [PubMed]
- Simoes, D.C.M.; Vogiatzis, I. Can muscle protein metabolism be specifically targeted by exercise training in COPD? J. Thorac. Dis. 2018, 10, S1367–S1376. [Google Scholar] [CrossRef]
- Gillies, A.R.; Lieber, R.L. Structure and function of the skeletal muscle extracellular matrix. Muscle Nerve 2011, 44, 318–331. [Google Scholar] [CrossRef] [Green Version]
- Fry, C.S.; Kirby, T.J.; Kosmac, K.; McCarthy, J.J.; Peterson, C.A. Myogenic Progenitor Cells Control Extracellular Matrix Production by Fibroblasts during Skeletal Muscle Hypertrophy. Cell Stem Cell 2017, 20, 56–69. [Google Scholar] [CrossRef] [Green Version]
- Calve, S.; Odelberg, S.J.; Simon, H.G. A transitional extracellular matrix instructs cell behavior during muscle regeneration. Dev. Biol. 2010, 344, 259–271. [Google Scholar] [CrossRef] [Green Version]
- Street, S.F. Lateral transmission of tension in frog myofibers: A myofibrillar network and transverse cytoskeletal connections are possible transmitters. J. Cell. Physiol. 1983, 114, 346–364. [Google Scholar] [CrossRef]
- Csapo, R.; Gumpenberger, M.; Wessner, B. Skeletal Muscle Extracellular Matrix—What Do We Know About Its Composition, Regulation, and Physiological Roles? A Narrative Review. Front. Physiol. 2020, 11, 253. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.; Price, F.; Rudnicki, M.A. Satellite cells and the muscle stem cell niche. Physiol. Rev. 2013, 93, 23–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seynnes, O.R.; de Boer, M.; Narici, M.V. Early skeletal muscle hypertrophy and architectural changes in response to high-intensity resistance training. J. Appl. Physiol. 2007, 102, 368–373. [Google Scholar] [CrossRef] [PubMed]
- Snijders, T.; Verdijk, L.B.; van Loon, L.J. The impact of sarcopenia and exercise training on skeletal muscle satellite cells. Ageing Res. Rev. 2009, 8, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.M.; Loenneke, J.P.; Jo, E.; Wilson, G.J.; Zourdos, M.C.; Kim, J.-S. The effects of endurance, strength, and power training on muscle fiber type shifting. J. Strength Cond. Res. 2012, 26, 1724–1729. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Group, P. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. Int. J. Surg. 2010, 8, 336–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Brandt, J.; Spruit, M.A.; Derave, W.; Hansen, D.; Vanfleteren, L.E.; Burtin, C. Changes in structural and metabolic muscle characteristics following exercise-based interventions in patients with COPD: A systematic review. Expert Rev. Respir. Med. 2016, 10, 521–545. [Google Scholar] [CrossRef] [PubMed]
- Pratt, J.; Boreham, C.; Ennis, S.; Ryan, A.W.; De Vito, G. Genetic Associations with Aging Muscle: A Systematic Review. Cells 2019, 9, 12. [Google Scholar] [CrossRef] [Green Version]
- Roig, M.; O’Brien, K.; Kirk, G.; Murray, R.; McKinnon, P.; Shadgan, B.; Reid, W.D. The effects of eccentric versus concentric resistance training on muscle strength and mass in healthy adults: A systematic review with meta-analysis. Br. J. Sports Med. 2009, 43, 556–568. [Google Scholar] [CrossRef]
- Garg, K.; Boppart, M.D. Influence of exercise and aging on extracellular matrix composition in the skeletal muscle stem cell niche. J. Appl. Physiol. 2016, 121, 1053–1058. [Google Scholar] [CrossRef] [Green Version]
- Sterne, J.A.; Hernan, M.A.; Reeves, B.C.; Savovic, J.; Berkman, N.D.; Viswanathan, M.; Henry, D.; Altman, D.G.; Ansari, M.T.; Boutron, I.; et al. ROBINS-I: A tool for assessing risk of bias in non-randomised studies of interventions. BMJ 2016, 355, i4919. [Google Scholar] [CrossRef] [Green Version]
- Sterne, J.A.C.; Savovic, J.; Page, M.J.; Elbers, R.G.; Blencowe, N.S.; Boutron, I.; Cates, C.J.; Cheng, H.Y.; Corbett, M.S.; Eldridge, S.M.; et al. RoB 2: A revised tool for assessing risk of bias in randomised trials. BMJ 2019, 366, l4898. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.L.; Wang, Y.Y.; Yang, Z.H.; Huang, D.; Weng, H.; Zeng, X.T. Methodological quality (risk of bias) assessment tools for primary and secondary medical studies: What are they and which is better? Mil. Med. Res. 2020, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Damas, F.; Ugrinowitsch, C.; Libardi, C.A.; Jannig, P.R.; Hector, A.J.; McGlory, C.; Lixandrao, M.E.; Vechin, F.C.; Montenegro, H.; Tricoli, V.; et al. Resistance training in young men induces muscle transcriptome-wide changes associated with muscle structure and metabolism refining the response to exercise-induced stress. Eur. J. Appl. Physiol. 2018, 118, 2607–2616. [Google Scholar] [CrossRef]
- Deshmukh, A.S.; Steenberg, D.E.; Hostrup, M.; Birk, J.B.; Larsen, J.K.; Santos, A.; Kjobsted, R.; Hingst, J.R.; Scheele, C.C.; Murgia, M.; et al. Deep muscle-proteomic analysis of freeze-dried human muscle biopsies reveals fiber type-specific adaptations to exercise training. Nat. Commun. 2021, 12, 304. [Google Scholar] [CrossRef]
- Hjorth, M.; Norheim, F.; Meen, A.J.; Pourteymour, S.; Lee, S.; Holen, T.; Jensen, J.; Birkeland, K.I.; Martinov, V.N.; Langleite, T.M.; et al. The effect of acute and long-term physical activity on extracellular matrix and serglycin in human skeletal muscle. Physiol. Rep. 2015, 3, e12473. [Google Scholar] [CrossRef]
- Kanzleiter, T.; Rath, M.; Gorgens, S.W.; Jensen, J.; Tangen, D.S.; Kolnes, A.J.; Kolnes, K.J.; Lee, S.; Eckel, J.; Schurmann, A.; et al. The myokine decorin is regulated by contraction and involved in muscle hypertrophy. Biochem. Biophys. Res. Commun. 2014, 450, 1089–1094. [Google Scholar] [CrossRef]
- Karlsen, A.; Soendenbroe, C.; Malmgaard-Clausen, N.M.; Wagener, F.; Moeller, C.E.; Senhaji, Z.; Damberg, K.; Andersen, J.L.; Schjerling, P.; Kjaer, M.; et al. Preserved capacity for satellite cell proliferation, regeneration, and hypertrophy in the skeletal muscle of healthy elderly men. FASEB J. 2020, 34, 6418–6436. [Google Scholar] [CrossRef] [Green Version]
- Kern, H.; Barberi, L.; Lofler, S.; Sbardella, S.; Burggraf, S.; Fruhmann, H.; Carraro, U.; Mosole, S.; Sarabon, N.; Vogelauer, M.; et al. Electrical stimulation counteracts muscle decline in seniors. Front. Aging Neurosci. 2014, 6, 189. [Google Scholar] [CrossRef] [Green Version]
- Makhnovskii, P.A.; Zgoda, V.G.; Bokov, R.O.; Shagimardanova, E.I.; Gazizova, G.R.; Gusev, O.A.; Lysenko, E.A.; Kolpakov, F.A.; Vinogradova, O.L.; Popov, D.V. Regulation of Proteins in Human Skeletal Muscle: The Role of Transcription. Sci. Rep. 2020, 10, 3514. [Google Scholar] [CrossRef] [Green Version]
- Nishida, Y.; Tanaka, H.; Tobina, T.; Murakami, K.; Shono, N.; Shindo, M.; Ogawa, W.; Yoshioka, M.; St-Amand, J. Regulation of muscle genes by moderate exercise. Int. J. Sports Med. 2010, 31, 656–670. [Google Scholar] [CrossRef] [PubMed]
- Norheim, F.; Langleite, T.M.; Hjorth, M.; Holen, T.; Kielland, A.; Stadheim, H.K.; Gulseth, H.L.; Birkeland, K.I.; Jensen, J.; Drevon, C.A. The effects of acute and chronic exercise on PGC-1alpha, irisin and browning of subcutaneous adipose tissue in humans. FEBS J. 2014, 281, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Norheim, F.; Raastad, T.; Thiede, B.; Rustan, A.C.; Drevon, C.A.; Haugen, F. Proteomic identification of secreted proteins from human skeletal muscle cells and expression in response to strength training. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E1013–E1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olstad, O.K.; Gautvik, V.T.; LeBlanc, M.; Kvernevik, K.J.; Utheim, T.P.; Runningen, A.; Wiig, H.; Kirkegaard, C.; Raastad, T.; Reppe, S.; et al. Postmenopausal osteoporosis is a musculoskeletal disease with a common genetic trait which responds to strength training: A translational intervention study. Ther. Adv. Musculoskelet. Dis. 2020, 12, 1759720X20929443. [Google Scholar] [CrossRef] [PubMed]
- Radom-Aizik, S.; Hayek, S.; Shahar, I.; Rechavi, G.; Kaminski, N.; Ben-Dov, I. Effects of aerobic training on gene expression in skeletal muscle of elderly men. Med. Sci. Sports Exerc. 2005, 37, 1680–1696. [Google Scholar] [CrossRef] [PubMed]
- Raue, U.; Trappe, T.A.; Estrem, S.T.; Qian, H.R.; Helvering, L.M.; Smith, R.C.; Trappe, S. Transcriptome signature of resistance exercise adaptations: Mixed muscle and fiber type specific profiles in young and old adults. J. Appl. Physiol. 2012, 112, 1625–1636. [Google Scholar] [CrossRef] [Green Version]
- Riedl, I.; Yoshioka, M.; Nishida, Y.; Tobina, T.; Paradis, R.; Shono, N.; Tanaka, H.; St-Amand, J. Regulation of skeletal muscle transcriptome in elderly men after 6 weeks of endurance training at lactate threshold intensity. Exp. Gerontol. 2010, 45, 896–903. [Google Scholar] [CrossRef]
- Robinson, M.M.; Dasari, S.; Konopka, A.R.; Johnson, M.L.; Manjunatha, S.; Esponda, R.R.; Carter, R.E.; Lanza, I.R.; Nair, K.S. Enhanced Protein Translation Underlies Improved Metabolic and Physical Adaptations to Different Exercise Training Modes in Young and Old Humans. Cell Metab. 2017, 25, 581–592. [Google Scholar] [CrossRef] [Green Version]
- Timmons, J.A.; Knudsen, S.; Rankinen, T.; Koch, L.G.; Sarzynski, M.; Jensen, T.; Keller, P.; Scheele, C.; Vollaard, N.B.; Nielsen, S.; et al. Using molecular classification to predict gains in maximal aerobic capacity following endurance exercise training in humans. J. Appl. Physiol. 2010, 108, 1487–1496. [Google Scholar] [CrossRef] [Green Version]
- Valdivieso, P.; Toigo, M.; Hoppeler, H.; Fluck, M. T/T homozygosity of the tenascin-C gene polymorphism rs2104772 negatively influences exercise-induced angiogenesis. PLoS ONE 2017, 12, e0174864. [Google Scholar] [CrossRef] [Green Version]
- Walton, R.G.; Kosmac, K.; Mula, J.; Fry, C.S.; Peck, B.D.; Groshong, J.S.; Finlin, B.S.; Zhu, B.; Kern, P.A.; Peterson, C.A. Human skeletal muscle macrophages increase following cycle training and are associated with adaptations that may facilitate growth. Sci. Rep. 2019, 9, 969. [Google Scholar] [CrossRef] [Green Version]
- Fragala, M.S.; Jajtner, A.R.; Beyer, K.S.; Townsend, J.R.; Emerson, N.S.; Scanlon, T.C.; Oliveira, L.P.; Hoffman, J.R.; Stout, J.R. Biomarkers of muscle quality: N-terminal propeptide of type III procollagen and C-terminal agrin fragment responses to resistance exercise training in older adults. J. Cachexia Sarcopenia Muscle 2014, 5, 139–148. [Google Scholar] [CrossRef]
- Kim, H.J.; So, B.; Choi, M.; Kang, D.; Song, W. Resistance exercise training increases the expression of irisin concomitant with improvement of muscle function in aging mice and humans. Exp. Gerontol. 2015, 70, 11–17. [Google Scholar] [CrossRef]
- Alghadir, A.H.; Gabr, S.A.; Al-Eisa, E. Cellular fibronectin response to supervised moderate aerobic training in patients with type 2 diabetes. J. Phys. Ther. Sci. 2016, 28, 1092–1099. [Google Scholar] [CrossRef] [Green Version]
- McKee, T.J.; Perlman, G.; Morris, M.; Komarova, S.V. Extracellular matrix composition of connective tissues: A systematic review and meta-analysis. Sci. Rep. 2019, 9, 10542. [Google Scholar] [CrossRef] [Green Version]
- Iozzo, R.V.; Schaefer, L. Proteoglycan form and function: A comprehensive nomenclature of proteoglycans. Matrix Biol. 2015, 42, 11–55. [Google Scholar] [CrossRef]
- Guillon, E.; Bretaud, S.; Ruggiero, F. Slow Muscle Precursors Lay Down a Collagen XV Matrix Fingerprint to Guide Motor Axon Navigation. J. Neurosci. 2016, 36, 2663–2676. [Google Scholar] [CrossRef]
- Grounds, M.D.; Sorokin, L.; White, J. Strength at the Extracellular Matrix-Muscle Interface. Scand. J. Med. Sci. Sports 2005, 15, 381–391. [Google Scholar] [CrossRef]
- Gawlik, K.I.; Durbeej, M. Skeletal muscle laminin and MDC1A: Pathogenesis and treatment strategies. Skelet. Muscle 2011, 1, 1–13. [Google Scholar] [CrossRef]
- Kielty, C.M.; Sherratt, M.J.; Shuttleworth, C.A. Elastic fibres. J. Cell Sci. 2002, 115, 2817–2828. [Google Scholar] [CrossRef]
- Ghanemi, A.; Yoshioka, M.; St-Amand, J. Secreted Protein Acidic and Rich in Cysteine as A Regeneration Factor: Beyond the Tissue Repair. Life 2021, 11, 38. [Google Scholar] [CrossRef] [PubMed]
- Brandan, E.; Gutierrez, J. Role of skeletal muscle proteoglycans during myogenesis. Matrix Biol. 2013, 32, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Goetsch, S.C.; Hawke, T.J.; Gallardo, T.D.; Richardson, J.A.; Garry, D.J. Transcriptional profiling and regulation of the extracellular matrix during muscle regeneration. Physiol. Genom. 2003, 14, 261–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogiatzis, I.; Simoes, D.C.; Stratakos, G.; Kourepini, E.; Terzis, G.; Manta, P.; Athanasopoulos, D.; Roussos, C.; Wagner, P.D.; Zakynthinos, S. Effect of pulmonary rehabilitation on muscle remodelling in cachectic patients with COPD. Eur. Respir. J. 2010, 36, 301–310. [Google Scholar] [CrossRef] [Green Version]
- Stenina-Adognravi, O.; Plow, E.F. Thrombospondin-4 in tissue remodeling. Matrix Biol. 2019, 75–76, 300–313. [Google Scholar] [CrossRef]
- Jarvinen, T.A.; Jozsa, L.; Kannus, P.; Jarvinen, T.L.; Hurme, T.; Kvist, M.; Pelto-Huikko, M.; Kalimo, H.; Jarvinen, M. Mechanical loading regulates the expression of tenascin-C in the myotendinous junction and tendon but does not induce de novo synthesis in the skeletal muscle. J. Cell Sci. 2003, 116, 857–866. [Google Scholar] [CrossRef] [Green Version]
- Crameri, R.M.; Langberg, H.; Teisner, B.; Magnusson, P.; Schroder, H.D.; Olesen, J.L.; Jensen, C.H.; Koskinen, S.; Suetta, C.; Kjaer, M. Enhanced procollagen processing in skeletal muscle after a single bout of eccentric loading in humans. Matrix Biol. 2004, 23, 259–264. [Google Scholar] [CrossRef]
- Sancho-Munoz, A.; Guitart, M.; Rodriguez, D.A.; Gea, J.; Martinez-Llorens, J.; Barreiro, E. Deficient muscle regeneration potential in sarcopenic COPD patients: Role of satellite cells. J. Cell. Physiol. 2021, 236, 3083–3098. [Google Scholar] [CrossRef]
- Thalacker-Mercer, A.E.; Dell’Italia, L.J.; Cui, X.; Cross, J.M.; Bamman, M.M. Differential genomic responses in old vs. young humans despite similar levels of modest muscle damage after resistance loading. Physiol. Genom. 2010, 40, 141–149. [Google Scholar] [CrossRef] [Green Version]
- Heinemeier, K.M.; Olesen, J.L.; Haddad, F.; Langberg, H.; Kjaer, M.; Baldwin, K.M.; Schjerling, P. Expression of collagen and related growth factors in rat tendon and skeletal muscle in response to specific contraction types. J. Physiol. 2007, 582, 1303–1316. [Google Scholar] [CrossRef]
- Wessner, B.; Liebensteiner, M.; Nachbauer, W.; Csapo, R. Age-specific response of skeletal muscle extracellular matrix to acute resistance exercise: A pilot study. Eur. J. Sport Sci. 2019, 19, 354–364. [Google Scholar] [CrossRef] [Green Version]
- Bentzinger, C.F.; Wang, Y.X.; Dumont, N.A.; Rudnicki, M.A. Cellular dynamics in the muscle satellite cell niche. EMBO Rep. 2013, 14, 1062–1072. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.K.Y.; Chan, A.S.F.; Rubab, A.; Chan, W.C.W.; Chan, D. Extracellular Matrix and Cellular Plasticity in Musculoskeletal Development. Front. Cell Dev. Biol. 2020, 8, 781. [Google Scholar] [CrossRef]
- Lukjanenko, L.; Jung, M.J.; Hegde, N.; Perruisseau-Carrier, C.; Migliavacca, E.; Rozo, M.; Karaz, S.; Jacot, G.; Schmidt, M.; Li, L.; et al. Loss of fibronectin from the aged stem cell niche affects the regenerative capacity of skeletal muscle in mice. Nat. Med. 2016, 22, 897–905. [Google Scholar] [CrossRef] [Green Version]
- Miura, T.; Kishioka, Y.; Wakamatsu, J.; Hattori, A.; Hennebry, A.; Berry, C.J.; Sharma, M.; Kambadur, R.; Nishimura, T. Decorin binds myostatin and modulates its activity to muscle cells. Biochem. Biophys. Res. Commun. 2006, 340, 675–680. [Google Scholar] [CrossRef]
- Miura, T.; Kishioka, Y.; Wakamatsu, J.; Hattori, A.; Nishimura, T. Interaction between myostatin and extracellular matrix components. Anim. Sci. J. 2010, 81, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Radwanska, A.; Grall, D.; Schaub, S.; Divonne, S.B.F.; Ciais, D.; Rekima, S.; Rupp, T.; Sudaka, A.; Orend, G.; Van Obberghen-Schilling, E. Counterbalancing anti-adhesive effects of Tenascin-C through fibronectin expression in endothelial cells. Sci. Rep. 2017, 7, 12762. [Google Scholar] [CrossRef] [Green Version]
- Pessentheiner, A.R.; Ducasa, G.M.; Gordts, P. Proteoglycans in Obesity-Associated Metabolic Dysfunction and Meta-Inflammation. Front. Immunol. 2020, 11, 769. [Google Scholar] [CrossRef]
- Yoo, H.J.; Hwang, S.Y.; Cho, G.J.; Hong, H.C.; Choi, H.Y.; Hwang, T.G.; Kim, S.M.; Bluher, M.; Youn, B.S.; Baik, S.H.; et al. Association of glypican-4 with body fat distribution, insulin resistance, and nonalcoholic fatty liver disease. J. Clin. Endocrinol. Metab. 2013, 98, 2897–2901. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Author | Bias Due to Confounding | Bias in Selection of Participants into the Study | Bias in Classification of Interventions | Bias Due to Deviations from Intended Interventions | Bias Due to Missing Data | Bias in Measurement of Outcomes | Bias in Selection of the Reported Result | Overall Bias |
|---|---|---|---|---|---|---|---|---|
| Damas et al., 2018 [24] | Low | Low | Low | Low | Low | Low | Low | Low |
| Deshmukh et al., 2021 [25] | Low | Low | Low | Low | Low | Moderate | Low | Moderate |
| Hjorth et al., 2015 [26] | Low | Moderate | Low | Low | Low | Unclear | Low | Moderate |
| Kanzleiter et al., 2014 [27] | Low | Low | Low | Low | Low | Unclear | Low | Moderate |
| Karlsen et al., 2020 [28] | Low | Low | Low | Low | Low | Moderate | Low | Moderate |
| Kern et al., 2014 [29] | Moderate | Low | Low | Low | Low | Low | Low | Moderate |
| Makhnovskii et al., 2020 [30] | Low | Moderate | Low | Unclear | Low | Low | Low | Moderate |
| Nishida et al., 2010 [31] | Low | Low | Low | Low | Low | Low | Low | Low |
| Norheim et al., 2011 [32] | Low | Unclear | Low | Low | Low | Low | Low | Moderate |
| Norheim et al., 2014 [33] | Low | Low | Low | Low | Low | Unclear | Low | Moderate |
| Olstad et al., 2020 [34] | Unclear | Low | Low | Low | Low | Low | Low | Moderate |
| Radom-Aizak et al., 2005 [35] | Low | Low | Low | Low | Low | Low | Unclear | Moderate |
| Raue et al., 2012 [36] | Low | Low | Low | Low | Low | Low | Low | Low |
| Riedl et al., 2010 [37] | Low | Low | Low | Low | Low | Low | Low | Low |
| Robinson et al., 2017 [38] | Low | Low | Low | Low | Low | Low | Low | Low |
| Timmons et al., 2010 [39] | Low | Low | Low | Low | Low | Low | Low | Low |
| Valdivierso et al., 2017 [40] | Low | Low | Low | Low | Low | Low | Low | Low |
| Walton et al., 2019 [41] | Low | Low | Low | Low | Moderate | Low | Low | Moderate |
| Author | Randomization Process Bias | Deviation from the Intended Intervention Bias | Missing Outcome Bias | Measurement of the Outcome Bias | Selection of Reported Results Bias | Overall Risk of Bias |
|---|---|---|---|---|---|---|
| Alghadir et al., 2016 [42] | High | Low | Low | High | Moderate | High |
| Fragala et al., 2014 [43] | Low | Low | Low | Unclear | Low | Moderate |
| Kim et al., 2015 [44] | Low | Low | Low | Unclear | Low | Moderate |
| First Author, Year of Publication | Country | Participant Group (n); | Age ± SD | Study Design | Experimental Group Intervention | Experimental Duration and Frequency, | Attrition | Outcome Measures | Δ Muscle Remodelling Post-Training (within Group) | Δ Muscle Remodelling Post-Training (between Groups) |
|---|---|---|---|---|---|---|---|---|---|---|
| Sex (n, %) | or ± SEM(*) | Dosage (h) | (Reasons) | |||||||
| Damas et al., 2018 [24] | Brazil | Exercise (9); Male (9, 100%) | 26 ± 2 | Pre–post study | RET: it involved two exercises for lower body. | 10 weeks (2x/week) Dosage = N/A | 1 participant (male) removed | N/A | N/A | N/A |
| Deshmukh et al., 2021 [25] | Denmark | Exercise (5); Male (5, 100%) | 24 ± 1 * | Sub-cohort of pre–post study | AET: participants performed indoor cycling exercise (intensity ranged from 75–90% of maximal heart rate): 3 out of 4 sessions performed at home, 1 out of 4 at the laboratory. | 12 weeks (4x/weeks) Dosage = 48 | None | NA | NA | NA |
| Fragala et al., 2014 [43] | USA | Exercise (12); Male (7, 58.3%), Female (5, 41.7%) | 70.5 ± 6.9 | Pilot RCT | Supervised RET: 60–90 min (≈70–85% of RM). | 6 weeks (2x/week) Dosage = 12–18 | None | Muscle strength/ capacity | ||
| Leg extension strength (kg) | Exercise: ↑ 29.0% (p < 0.001) Control: NS | NS | ||||||||
| Control (11); Male (6, 54.5%), Female (5, 45.5%) | 69.6 ± 5.5 | Maintenance of normal physical activities. | Muscle quality (relative strength) | Exercise: ↑ 28.0% (p < 0.001) Control: NS | NS | |||||
| Body composition | ||||||||||
| LBM (kg) | Exercise:↑ 0.2% (NS) | NS | ||||||||
| Control: NS | ||||||||||
| Muscle architecture | ||||||||||
| Muscle cross-sectional area (CSA) (cm2) | Exercise:↑ 8.3% (NS) Control: NS | NS | ||||||||
| Hjorth et al., 2015 [26] | Norway | Exercise (26); Male (26, 100%) | 51.2 ± 6.6 | Pre–post study | Supervised CT: 2 intervals bicycle sessions and 2 whole body strength-training sessions per week. Each session lasted 1 h. | 12 weeks (4x/week) Dosage = 48 | None | Muscle strength/ capacity | N/A | |
| Leg extension strength (kg) | ↑ 9.6% (p < 0.001) | |||||||||
| Body composition | ||||||||||
| Fat mass (L) | ↓ 8.2% (p < 0.001) | |||||||||
| Muscle architecture | ||||||||||
| Thigh muscle area (cm2) | ↑ 7.5% (p < 0.001) | |||||||||
| Ventilatory changes | ||||||||||
| VO2max (mL/kg*min) | ↑ 11.2% (p < 0.001) | |||||||||
| Kanzleiter et al., 2014 [27] | Norway/Germany | Exercise (26); Male (26, 100%); Normal glucose group (13); Pre-diabetes group (13) | 51.2 ± 6.6 | Pre–post study | Supervised CT: 2 intervals bicycle sessions and 2 whole body strength-training sessions per week. Each session lasted 1 h. | 12 weeks (4x/week) Dosage = 48 | None | N/A | N/A | N/A |
| Karlsen et al., 2020 [28] | Denmark | Pre–post study | Supervised heavy-load RET: sessions involved 3 exercises for lower body and 2 optional for upper body | 13 weeks (3x/week) Dosage = N/A | 2 Five old and two young participants did not complete the intervention. | Muscle strength/ capacity | N/A | |||
| Isometric knee extensor peak torque (Nm) | Young: ↑ 14.4 % (p < 0.01) | |||||||||
| Older: ↑ 14.3% (p < 0.001) | ||||||||||
| Isokinetic knee extensor peak torque (Nm) | Young: ↑ 11.9% (p < 0.05) | |||||||||
| Older: ↑ 9% (p < 0.05) | ||||||||||
| Body composition | ||||||||||
| Thin lean mass (kg) | Young: ↑ 6.7% (p < 0.001) | |||||||||
| Young (7); Male (7, 100%) | 25 ± 3 | Older: ↑ 6% (p < 0.001) | ||||||||
| Muscle architecture | ||||||||||
| Older (19); Male (19, 100%) | 67 ± 4 | CSA VL (μm2) | Young: ↑ 11.9% (p < 0.001) | |||||||
| Older: ↑ 14.5% (p < 0.001) | ||||||||||
| CSA QF (μm2) | Young: ↑ 8.9% (p < 0.01) | |||||||||
| Older: ↑ 10.8% (p < 0.001) | ||||||||||
| CSA type I fibres (μm2) | Young: ↓0.2% (NS) Older: ↑ 5.7% (NS) | |||||||||
| CSA type II fibres (μm2) | Young: ↑ 10.2% (NS) | |||||||||
| Older: ↑ 25.8% | ||||||||||
| (p < 0.001) | ||||||||||
| Type I fibres (%) | Young: ↓ 2.3% (NS) | |||||||||
| Older: ↑ 1.9% (NS) | ||||||||||
| Kern et al., 2014 [29] | Italy/Austria | 73.1 ± 6.9 | Pre–post study | ES training at home: performed with a two-channel custom-built battery-powered stimulator3 × 10 min each session. | 9 weeks (2x/week for the first 3 weeks and 3x/week for the following 6 weeks) Dosage = 12 | None | Muscle strength/ capacity | N/A | ||
| Torque (Nm/kg) | ↑ 6.0 ± 4.9 (p < 0.05) | |||||||||
| Muscle architecture | ||||||||||
| All fibres size (μm) | NS | |||||||||
| Exercise (16); Male (8, 50%), Female (8, 50%) | Type I fibres size (μm) | ↓ 3.6% (p < 0.0001) | ||||||||
| Type I fibres percentage (%) | ↓ 7.2% (N.S) | |||||||||
| Type IIa fibres size (μm) | ↑ 2.2% (p < 0.0001) | |||||||||
| Type IIa fibres percentage (%) | ↑ 8.9% (N.S) | |||||||||
| Kim et al., 2015 [44] | Korea | Exercise (22); Female (22, 100%) | 74.5 ± 0.6 * | RCT | RET: it involved 2 supervised and 3 home-based sessions. Progressive intensity of the intervention. | 12 weeks (5x/week) Dosage = 60 | 3 Ten participants did not complete the intervention. | Muscle strength/ capacity | ||
| Grip strength (kg) | Exercise:↑ 27.0% (p < 0.001) | p < 0.001 | ||||||||
| Knee extensor strength 60°/s (N) | Exercise:↑ 42.1% (p < 0.001) | p = 0.019 | ||||||||
| Knee flexor strength (N) | Exercise:↓ 1.5% (NS) | p = 0.002 | ||||||||
| Control (8); Female (8, 100%) | 76.05 ± 2.0 * | Maintenance of normal physical activities and performance of one hour stretching once a week | Knee extensor strength 180°/s (N) | Exercise:↑ 33.7% (NS) | NS | |||||
| Knee flexor strength (N) | Exercise:↓ 19.4% (p < 0.001) | p = 0.028 | ||||||||
| Body composition | ||||||||||
| Waist–hip ratio (WHR) | Exercise:↓ 1.2% (NS) | N/A | ||||||||
| Arm circumference (cm) | Exercise:↓ 5.1% (NS) | N/A | ||||||||
| Thigh circumference (cm) | Exercise:↓ 1.3% (NS) | N/A | ||||||||
| Makhnovskii et al., 2020 [30] | Russia | Exercise (7); Male (7, 100%) | 22.5 ± 1.5 * | Pre–post study | AET: participants alternated continuous (intensity at 70% LT4) and intermittent exercise ((3 min, 50% LT4 + 2 min, 85% LT4) x12)) on different days. | 5 weeks (7x/week) Dosage = 35 | None | N/A | N/A | N/A |
| Nishida et al., 2010 [31] | Japan | Exercise (6); Male (6, 100%) | 19–32 | Pre–post study | Supervised AET: participants performed the session for 60 min using an upright cycle ergometer. Training intensity at the LT level. | 12 weeks (5x/week) Dosage = 60 | None | Body composition | N/A | |
| Fat percentage (%) | ↓ 2.2% (NS) | |||||||||
| Ventilatory changes | ||||||||||
| VO2max (mL/kg * min) | ↑ 8.7% (NS) | |||||||||
| VO2 at LT (mL/kg * min) | ↑ 62.5% (p < 0.05) | |||||||||
| VO2max at LT (%) | ↑ 48.9% (p < 0.05) | |||||||||
| Norheim et al., 2011 [32] | Norway | Exercise (13); Male (13, 100%) | 26.8 (19–35) | Sub-cohort of pre–post study | RET: it involved 1–3 sets of leg press, leg extension, leg curl, seated chest press, seated rowing, latissimus dorsi pull-down, biceps curl, and shoulder press. | 11 weeks (3x/week) Dosage = N/A | None | N/A | N/A | N/A |
| Norheim et al., 2014 [33] | Norway | Exercise (26); Male (26, 100%), Normal glucose group (13), Pre-diabetes group (13) | 51.2 ± 6.6 | Pre–post study | Supervised CT: it involved 2 interval bicycle sessions and 2 whole body strength-training sessions per week. Each session lasted 1 h. | 12 weeks (4x/week) Dosage = 48 | None | N/A | N/A | N/A |
| Radom-Aizak et al., 2005 [35] | Israel | Exercise (6); Male (6, 100%) | 68.0 ± 2.7 * | Pre–post study | AET: participants performed 45 min sessions (from the 3rd–12th week) on a cycle ergometer at 80% of the predetermined HRmax. | 12 weeks (3x/week) Dosage = 27 | None | Ventilatory changes | N/A | |
| VO2max (L/min) | ↑ 17.8% (p = 0.009) | |||||||||
| Anaerobic threshold (%) | ↑ 21% (p = 0.008) | |||||||||
| Raue et al., 2012 [36] | USA | Young (16); Male (8, 50%), Female (8, 50%) | 24 ± 4 | Pre–post study | RET: it involved 3 sets of 10 bilateral knee extensions (70–75% of 1 RM). | 12 weeks (3x/week) Dosage = N/A | None | Muscle strength/ capacity | N/A | |
| Leg extension strength (kg) | ↑ 5.7–↑ 41.3 kg | |||||||||
| Older (12); Male (6, 50%), Female (6, 50%) | 84 ± 3 | |||||||||
| Muscle architecture | ||||||||||
| Thigh muscle CSA (cm2) | ↓ 1.2–↑ 10.4 cm2 | |||||||||
| Riedl et al., 2010 [37] | Japan | Exercise (7); Male (7, 100%) | 64 ± 2.6 | Pre–post study | Supervised AET: participants performed sessions of 60 min on a cycle ergometer. Training intensity at the LT level | 6 weeks (5x/week) Dosage = 30 | None | Body composition | N/A | |
| Fat percentage (%) | ↓ 9.9% (p < 0.05) | |||||||||
| Ventilatory changes | ||||||||||
| VO2 at LT (%) | ↑ 8.3% (p < 0.05) | |||||||||
| VO2 max (mL/FFM kg/min) | ↑ 7.3% (p < 0.05) | |||||||||
| Robinson et al., 2017 [38] | USA | Young HIIT exercise (10); Male, Female | Pre–post study | HIIT: participants performed 3 sessions per week of cycling (4 × 4 min at >90% of VO2max separated by 3 min of pedalling at no load) and 2 sessions per week of treadmill walking (45 min at 70% of VO2max). | HIIT: 12 weeks (5x/week) Dosage = 34.8 | 4 Five young and three older participants did not complete the intervention. There was no information on number of males and females completing the study | Young HIIT | Absolute VO2max (mL/min) in young: ↑ following HIIT (p < 0.0001) > ↑ following RET (p < 0.048) and CT (p = 0.0001) | ||
| 25.4 ± 4.3 | ||||||||||
| Ventilatory changes | ||||||||||
| VO2max (mL/kg BW/min) | ↑ (p < 0.001) | |||||||||
| Older HIIT exercise (8); Male, Female | Body composition | |||||||||
| 70.7 ± 4.6 | FFM (kg) | ↑ (p < 0.05) | ||||||||
| Muscle strength/ capacity | Absolute VO2max (mL/min) in older: ↑ following HIIT (p < 0.0091) and CT (p = 0.0096) >↑ following RET (ns) | |||||||||
| Young RET (10); Male, Female | 23.7 ± 3.5 | Maximal leg strength (1 RM) leg press (AU/kg Leg FFM) | ↑ (NS) | |||||||
| RET: participants performed 2 sessions of lower and upper body exercises (4 sets of 8–12 repetitions), 2 days each per week. | RET: 12 weeks (5x/week) Dosage = N/A | |||||||||
| Older RET (8); Female | 70.3 ± 3.9 | |||||||||
| Older HIIT | ||||||||||
| Ventilatory changes | Relative VO2max (mL/min) in young: ↑ ~28% following HIIT (p < 0.0001) > ↑ ~17% following CT (p < 0.0001) > RET (ns) | |||||||||
| Young Combined exercise (8); Male, Female | 26.3 ± 2.7 | VO2max (mL/kg BW/min) | ↑ (p < 0.01) | |||||||
| CT after a 3 months SED: Following SED, participants underwent metabolic studies and performed CT of 5 days per week cycling (30 min at 70% VO2max) and 4 days per week weightlifting with fewer repetitions than RET. | Combined:12 weeks (9x/week) Dosage > 30 | Body composition | ||||||||
| FFM (kg) | ↑ (p < 0.05) | |||||||||
| Muscle strength/ capacity | ||||||||||
| Older Combined exercise (7); Male, Female | 68.6 ± 3.4 | |||||||||
| Maximal leg strength (1 RM) leg press) (AU/kg Leg FFM | ↑ (NS) | Relative VO2max (mL/min) in older: ↑ ~21% following CT (p < 0.0001) > ↑ ~17% following HIIT (p < 0.0001) > RET (ns) | ||||||||
| Young RET | ||||||||||
| Ventilatory changes | ||||||||||
| VO2max (mL/kg | ↑ (NS) | |||||||||
| BW/min) | ||||||||||
| Body composition | Leg strength: ↑ Following RET and CT > HIIT (NS) | |||||||||
| FFM (kg) | ↑ 4% (p < 0.0001) | |||||||||
| Muscle strength/ | ||||||||||
| Maximal leg strength | ↑ (p < 0.05)↑ | |||||||||
| (1 RM) leg press | ||||||||||
| (AU/kg Leg FFM) | ||||||||||
| Older RET | ||||||||||
| Ventilatory changes | ||||||||||
| VO2max (mL/kg BW/min) | ↑ (NS) | |||||||||
| Body composition | ||||||||||
| FFM (kg) | ↑ (p < 0.01) | |||||||||
| Muscle strength/ capacity | ||||||||||
| Maximal leg strength (1 RM) leg press (AU/kg Leg FFM) | ↑ (p < 0.05) | |||||||||
| Young CT | ||||||||||
| Ventilatory changes | ||||||||||
| VO2max (mL/kg BW/min) | ↑ (p < 0.001) | |||||||||
| Body composition | ||||||||||
| FFM (kg) | ↑ (p < 0.05) | |||||||||
| Muscle strength/ capacity | ||||||||||
| Maximal leg strength (1 RM) leg press (AU/kg Leg FFM) | ↑ (p < 0.05) | |||||||||
| Older CT | ||||||||||
| Ventilatory changes | ||||||||||
| VO2max (mL/kg BW/min) | ↑ (p < 0.01) | |||||||||
| Body composition | ||||||||||
| FFM (kg) | ↑ (p < 0.05) | |||||||||
| Muscle strength/ capacity | ||||||||||
| Maximal leg strength (1 RM) leg press (AU/kg Leg FFM) | ↑ (p < 0.05) | |||||||||
| Timmons et al., 2010 [39] | Sweden, Denmark, UK, USA | Exercise (24); Male (24, 100%) | 23 | Pre–post study | Supervised AET: participants performed 45 min cycling sessions. Training intensity customized to 70% of the pretraining VO2max. | 6 weeks (4x/week) Dosage = 18 | None | Ventilatory changes | N/A | |
| VO2 max (L/min) | ↑ 14% (N/A) | |||||||||
| Submax RER (ratio) | ↓ 10% (N/A) | |||||||||
| Valdivierso et al., 2017 [40] | Switzerland | Exercise (61); Male (61, 100%), A/A alleles (12), A/T alleles (38), T/T alleles (11) | 29.5 ± 9.3 | Pre–post study | AET: participants performed 30 min sessions on a cycle ergometer at a heart rate corresponding to 65% of Pmax. Training intensity maintained at ≈90% of maximal heart rate | 6 weeks (5x/week) Dosage = 15 | None | Muscle architecture | ||
| Muscle fibre area (μm2) | ↑ 8.3% (NS) | |||||||||
| Biopsy myofibrils (%) | ↓ 4.0% (p < 0.05) | |||||||||
| Capillary-to-fibre ratio (ALL) | ↑ 12.1% (p < 0.05) | |||||||||
| Capillary-to-fibre ratio: | ||||||||||
| A/A genotype | ↑ 25.0% (NS) | A/A vs. T/T (p < 0.05) | ||||||||
| A/T genotype | ↑ 12.6% (p < 0.05) | A allele carriers vs. T/T (p < 0.05) | ||||||||
| T/T genotype | ↓ 12.5% (NS) | |||||||||
| Capillary density (mm−2) | ↓ 5.5% (p < 0.05) | |||||||||
| Ventilatory changes | ||||||||||
| VO2max (mL/kg * min) | ↑ 8.5% (p < 0.05) | |||||||||
| Pmax (ergospirometry) (W) | ↑ 12.7% (p < 0.05) | |||||||||
| Walton et al., 2019 [41] | USA | Exercise (20); Male (4, 25%), Female (16, 75%) | 49.8 ± 2.3 * | Pre–post study | AET: participants performed 45 min sessions using a stationary cycle ergometer (at a target intensity corresponding to 65% of VO2max and ≈75–80% of maximum heart rate) | 12 weeks (3x/week) Dosage = 27 | None | N/A | N/A | N/A |
| Alghadir et al., 2016 [42] | Saudi Arabia | Exercise (25); Male with T2D (25, 100%) | 48.8 ± 14.6 | RCT | Supervised AET: 50 min in intensity defined by heart rate (THR max; 60–70%) | 12 weeks (3x/week) Dosage = 30 | None | N/A | N/A | N/A |
| Control (25); Male (25, 100%) | 48.7 ± 3.4 | Sedentary lifestyle | ||||||||
| Olstad et al., 2020 [34] | Norway | Pre–post study | Supervised heavy-load RET: it involved all major muscle groups. Gradual progression on the training loads was applied. Each session lasted ≈60 min. | 13 weeks (3x/week) Dosage = 39 | 5 One participant did not complete the study | Muscle strength/ capacity | N/A | |||
| Exercise healthy (18); Female (18, 100%) | 73.9 ± 5.7 | |||||||||
| Relative strength | Healthy: ↑ 32 ± 16% | |||||||||
| Osteoporotic: ↑ 31 ± 19% | ||||||||||
| Exercise osteoporotic (17); Female (17, 100%) | 78.0 ± 6.2 | |||||||||
| First Author, Year of Publication | Outcome Measure | Δ ECM Outcome Post Training within Group | Association between ECM and Muscle Remodelling Outcome within the Study |
|---|---|---|---|
| Damas et al., 2018 [24] | mRNA expression | N/A | |
| Collagens | |||
| COL3A1 | ↑ 146% (p < 0.05) | ||
| COL4A1 | ↑ 112% (p < 0.05) | ||
| COL5A2 | ↑ 95% (p < 0.05) | ||
| Glycoproteins | |||
| CTHRC1 | ↑ 105% (p < 0.05) | ||
| LAMB1 | ↑ 79% (p < 0.05) | ||
| THBS4 | ↑ 144% (p < 0.05) | ||
| PXDN | ↑ 81% (p < 0.05) | ||
| Deshmukh et al., 2021 [25] | Protein expression | N/A | |
| Glycoproteins | |||
| Agrin | Slow fibres: ↑ 147% (NS) | ||
| Fast fibres: ↑ 460% (p < 0.001) | |||
| Whole muscle: ↑ 130% (NS) | |||
| Thrombospondin 4 | Slow fibres: ↑ 240% (NS) | ||
| Fast fibres: ↑ 360% (p = 0.015) | |||
| Whole muscle: N/A | |||
| Peroxidasin | Slow fibres: ↑ 320% (p = 0.0017) | ||
| Fast fibres: ↑ 300% (NS) | |||
| Whole muscle: ↑ 360% (p = 0.0018) | |||
| Dermatopontin | Slow fibres: ↓ 70% (NS) | ||
| Fast fibres: ↓ 85% (NS) | |||
| Whole muscle: ↓ 16% (p = 0.0046) | |||
| Fibrillin-1 | Slow fibres: ↓ 100% (NS) | ||
| Fast fibres: ↓ 82% (NS) | |||
| Whole muscle: ↑ 280% (p = 0.0063) | |||
| Irisin precursor, fibronectin type III | Slow fibres: ↑ 108% (NS) | ||
| Fast fibres: ↓ 90% (NS) | |||
| Whole muscle: ↓ 70% (p = 0.025) | |||
| IGFN1 (Immunoglobulin-like and fibronectin type III domain containing | Slow fibres: ↑ 790% (p = 0.0028) | ||
| Fast fibres: ↑ 560% (p = 0.011) | |||
| Whole muscle: ↑ 225% (NS) | |||
| Laminin subunit alpha-1 | Slow fibres: N/A | ||
| Fast fibres: N/A | |||
| Whole muscle: ↑ 370% (p = 0.0415) | |||
| Laminin subunit alpha 4 | Slow fibres: ↑ 160% (p = 0.0119) | ||
| Fast fibres: ↑ 143% (NS) | |||
| Whole muscle: ↓ 93% (NS) | |||
| Laminin subunit alpha 5 | Slow fibres: ↑ 119% (NS) | ||
| Fast fibres: ↑ 99% (NS) | |||
| Whole muscle: ↑ 60% (p = 0.0431) | |||
| Microfibrillar-associated protein 4 | Slow fibres: ↓ 90% (NS) | ||
| Fast fibres: ↓ 39% (p = 0.043) | |||
| Whole muscle: ↑ 137% (NS) | |||
| Microfibrillar-associated protein 5 | Slow fibres: ↑ 440% (p = 0.0197) | ||
| Fast fibres: ↑ 62% (NS) | |||
| Whole muscle: ↑ 162% (NS) | |||
| Picachurin (EGFLAM) | Slow fibres: ↑ 300% (p = 0.0197) | ||
| Fast fibres: ↑ 510% (p < 0.001) | |||
| Whole muscle: ↑ 330% (p = 0.0008) | |||
| Fragala et al., 2014 [43] | Circulating markers (serum) | P3NP vs. LBM (r = 0.422, p = 0.045) | |
| C-terminal agrin fragment (CAF) | Exercise: ↑ 10.4% (NS) | ||
| Control: ↑ 0.3% (NS) | CAF vs. VL CSA (r = 0.542, p = 0.008) | ||
| N-terminal peptide of procollagen type III (P3NP) | Exercise: ↑ 7.9% (NS) | P3NP vs. muscle strength/quality (NS) | |
| Control: ↑ 1.9% (NS) | |||
| CAF vs. muscle strength/quality (NS) | |||
| Hjorth et al., 2015 [26] | mRNA expression | N/A | |
| Collagens | |||
| COL1A1 | ↑ 140% (p < 0.001) | ||
| COL1A2 | ↑ 80% (p < 0.001) | ||
| COL3A1 | ↑ 140% (p < 0.001) | ||
| COL4A1 | ↑ 140% (p < 0.001) | ||
| COL4A2 | ↑ 120% (p < 0.001) | ||
| COL5A1 | ↑ 50% (p < 0.001) | ||
| COL5A2 | ↑ 60% (p < 0.001) | ||
| COL6A6 | ↑ 100% (p < 0.001) | ||
| COL14A1 | ↑ 80% (p < 0.001) | ||
| COL15A1 | ↑ 50% (p < 0.001) | ||
| COL18A1 | ↑ 50% (p < 0.001) | ||
| Proteoglycans | |||
| ASPN | ↑ 80% (p < 0.001) | ||
| BGN | ↑ 110% (p < 0.001) | ||
| HSPG2 | ↑ 50% (p < 0.001) | ||
| OGN | ↑ 110% (p < 0.001) | ||
| OMD | ↑ 80% (p < 0.001) | ||
| ECM2 | ↑ 60% (p < 0.001) | ||
| LUM | ↑ 50% (p < 0.001) | ||
| GPC4 | ↓ N/A (p < 0.001) | ||
| CHAD | ↓ 52% (p < 0.001) | ||
| CSPG4 | ↑ 50% (p < 0.001) | ||
| Glycoproteins | |||
| AGRN | ↑ 60% (p < 0.001) | ||
| LAMA4 | ↑ 70% (p < 0.001) | ||
| LAMB1 | ↑ 70 % (p < 0.001 | ||
| LAMB3 | ↓ 56% (p < 0.001) | ||
| LAMC3 | ↑ 70% (p < 0.001) | ||
| THBS1 | ↑ 60% (p < 0.001) | ||
| THBS4 | ↑ 220% (p < 0.001) | ||
| NID1 | ↑ 60% (p < 0.001) | ||
| NID2 | ↑ 70% (p < 0.001) | ||
| PXDN | ↑ 200% (p < 0.001) | ||
| ELN | ↑ 50% (p < 0.001) | ||
| EMILIN3 | ↑ 60% (p < 0.001) | ||
| SPARC | ↑ 80% (p < 0.001) | ||
| CTHRC1 | ↑ 70% (p < 0.001) | ||
| Kanzleiter et al., 2014 [27] | mRNA expression | ||
| Proteoglycans | Δ Decorin expression vs. ΔLeg | ||
| DCN | Healthy: ↑ (p < 0.05) | press strength (kg) (r = 0.56, p = 0.047) | |
| Pre-diabetes: ↓ (NS) | |||
| Karlsen et al., 2020 [28] | mRNA expression | ||
| Collagens | |||
| COL1A1 | Young: (NS) | ||
| Older: ↑ (values not reported) (p < 0.05) | |||
| Kern et al., 2014 [29] | mRNA expression | ||
| Collagens | |||
| COL1 | ↑ (p < 0.005) | ||
| COL3 | ↑ (p < 0.005) | ||
| COL6 | ↑ (p < 0.005) | ||
| Kim et al., 2015 [44] | Circulating markers (serum) | ||
| Irisin | Exercise: ↑ 22.5% (p < 0.05) | Irisin vs. grip strength | |
| Control: NS | (r = 0.526, p = 0.002) | ||
| Irisin vs. leg strength | |||
| (r = 0.414, p = 0.003) | |||
| Makhnovskii et al., 2020 [30] | mRNA expression | N/A | |
| Collagens | |||
| COL1A1 | ↑ 1060% (p < 0.05) | ||
| COL1A2 | ↑ 440% (p < 0.05) | ||
| COL3A1 | ↑ 656% (p < 0.05) | ||
| COL4A2 | ↑ 439% (p < 0.05) | ||
| COL6A1 | ↑ 196% (p < 0.05) | ||
| COL6A2 | ↑ 232% (p < 0.05) | ||
| COL6A3 | ↑ 273% (p < 0.05) | ||
| COL14A1 | ↑ 608% (p < 0.05) | ||
| COL15A1 | ↑ 271% (p < 0.05) | ||
| Proteoglycans | |||
| ASPN | ↑ 359% (p < 0.05) | ||
| BGN | ↑ 435% (p < 0.05) | ||
| HSPG2 | ↑ 204% (p < 0.05) | ||
| OGN | ↑ 421% (p < 0.05) | ||
| LUM | ↑ 447% (p < 0.05) | ||
| DCN | ↑ (NS) | ||
| PRELP | ↑ (NS) | ||
| Glycoproteins | |||
| LAMB1 | ↑ 326% (p < 0.05) | ||
| LAMC1 | ↑ 175% (p < 0.05) | ||
| Protein expression | |||
| Collagens | |||
| Collagen Type I Alpha 1 Chain | ↑ 171% (p < 0.05) | ||
| Collagen Type I Alpha 2 Chain | ↑ 173% (p < 0.05) | ||
| Collagen Type III Alpha 1 Chain | ↑ 221% (p < 0.05) | ||
| Collagen Type XIV Alpha 1 Chain | ↑ 164% (p < 0.05) | ||
| Collagen Type VI Alpha 1 Chain | ↑124% (p < 0.05) | ||
| Collagen Type VI Alpha 2 Chain | ↑121% (p < 0.05) | ||
| Collagen Type VI Alpha 3 Chain | ↑124% (p < 0.05) | ||
| Proteoglycans | |||
| Asporin | ↑ 152% (p < 0.05) | ||
| Lumican | ↑ 123% (p < 0.05) | ||
| Prolargin (or Proline and arginine rich end leucine rich repeat protein) | ↑ 118% (p < 0.05) | ||
| Nishida et al., 2010 [31] | mRNA expression (using SAGE) | N/A | |
| Collagens | |||
| COL1A2 | ↑ 1200% (p < 0.05) | ||
| Proteoglycan | |||
| DCN | ↓ 2100% (p < 0.05) | ||
| Norheim et al., 2011 [32] | mRNA expression | N/A | |
| Collagens | |||
| COL1A1 | M. VL: ↑ 520 (p < 0.05) | ||
| M. TRAP: ↑ 4340% (p < 0.05) | |||
| Proteoglycans | |||
| LUM | M. VL: ↑ 250 (p < 0.05) | ||
| M. TRAP: ↑ 430 (p < 0.05) | |||
| ECM1 | M. VL: ↑ 180 (p < 0.05) | ||
| M. TRAP: ↑ 190 (p < 0.05) | |||
| Glycoproteins | |||
| SPARC | M. VL: ↑ 290 (p < 0.05) | ||
| M. TRAP: ↑ 960 (p < 0.05) | |||
| FN1 | M. VL: ↑ 180 (p < 0.05) | ||
| M. TRAP: ↑ 250 (p < 0.05) | |||
| Norheim et al., 2014 [33] | mRNA expression | N/A | |
| FNDC5 (Irisin) | Healthy: ↑ 40% (p < 0.05) | ||
| Pre-diabetes: ↑ 100% (p < 0.01) | |||
| Circulating markers (serum) | Healthy: ↓ (NS) | ||
| Irisin | Pre-diabetes: ↓ (NS) | ||
| Radom-Aizak et al., 2005 [35] | mRNA expression | N/A | |
| Collagens | |||
| COL3A1 | ↑ 111% (p = 0.0178) | ||
| Raue et al., 2012 [36] | mRNA expression | Pooled mRNA expression of COL4α2 vs. 1-RM (r = −0.418) | |
| Collagens | |||
| COL1A1 | Young: N/A | Pooled mRNA expression of COL4α3 vs. 1-RM (r = −0.344-(−0.486)) | |
| Older: ↑ 220–300% | |||
| COL1A2 | Young: N/A | Pooled mRNA expression of COL4α4 vs. 1-RM (r = -0.540-(0.623)) | |
| Older: ↑ 150–200% | Pooled mRNA expression of COL4α5 vs. 1-RM (r = −0.547) | ||
| COL3A1 | Young: N/A | Pooled mRNA expression of COLQ vs. 1-RM (r = 0.652) | |
| Older: ↑ 260–210% | Pooled mRNA expression of COL27α1 vs. 1-RM (r = −0.355-(−0.457)) | ||
| COL4A1 | Young: ↑ 190–200% | Pooled mRNA expression of COL28α1 vs. 1-RM (r = −0.180) | |
| Older: N/A | Pooled mRNA expression of CSPG4 vs. 1-RM (r = 0.421) | ||
| COL4A2 | Young: ↑ 170% | Pooled mRNA expression of COL4α2 vs. CSA (r = −0.405) | |
| Older: ↑ 220% | Pooled mRNA expression of COL4α3 vs. CSA (r = −0.345-(−0.462)) | ||
| COL5A1 | Young: N/A | Pooled mRNA expression of COL4α4 vs. CSA (r = −0.461-(0.486)) | |
| Older: ↑ 290% | Pooled mRNA expression of COL4α5 vs. CSA (r = −0.406) | ||
| COL5A2 | Young: N/A | Pooled mRNA expression of COLQ vs. CSA (r= 0.540) | |
| Older: ↑ 180% | Pooled mRNA expression of COL27α1 vs. CSA (r = −0.348) | ||
| COL5A3 | Young: N/A | ||
| Older: ↑ 170–180% | |||
| COL15A1 | Young: N/A | ||
| Older: ↑ 150% | |||
| Proteoglycans | |||
| ASPN | Young: N/A | ||
| Older: ↑ 200% | |||
| Glycoproteins | |||
| LAMA4 | Young: N/A | ||
| Older: ↑ 174% | |||
| LAMB1 | Young: N/A | ||
| Older: ↑ 165% | |||
| NID1 | Young: N/A | ||
| Older: ↑ 160–200% | |||
| NID2 | Young: N/A | ||
| Older: ↑ 194% | |||
| SPARC | Young: N/A | ||
| Older: ↑ 150–160% | |||
| THBS4 | Young: N/A | ||
| Older: ↑ 168% | |||
| CTHRC1 | Young: N/A | ||
| Older: ↑ 200% | |||
| Riedl et al., 2010 [37] | Number of tags per 100,000 SAGE tags | ||
| Collagens | |||
| COL3A1 | ↑ 14 | ||
| COL4A1 | ↑ 15 | ||
| Glycoproteins | |||
| SPARC | ↑ 20 | ||
| Robinson et al., 2017 [38] | mRNA expression | N/A | |
| Collagens | |||
| COL4A1 | HIIT young: ↑ 217% (p ≤ 0.05) | ||
| HIIT older: ↑ 361% (p ≤ 0.05) | |||
| RET young: ↑ 267% (p ≤ 0.05) | |||
| RET older: ↑ 236% (p ≤ 0.05) | |||
| CT young: ↑ 185% (p ≤ 0.05) | |||
| CT older: ↑ 197% (p ≤ 0.05) | |||
| COL4A2 | HIIT young: ↑ 188% (p ≤ 0.05) | ||
| HIIT older: ↑ 303% (p ≤ 0.05) | |||
| RET young: ↑ 219% (p ≤ 0.05) | |||
| RET older: ↑ 202% (p ≤ 0.05) | |||
| CT young: ↑ 172% (p ≤ 0.05) | |||
| CT older: ↑ 182% (p ≤ 0.05) | |||
| COL14A1 | HIIT young: (NS) | ||
| HIIT older: ↑ 165% (p ≤ 0.05) | |||
| RET young: (NS) | |||
| RET older: (NS) | |||
| CT young: (NS) | |||
| CT older: (NS) | |||
| Proteoglycans | HIIT young: ↑ 174% (p ≤ 0.05) | ||
| ASPN | HIIT older: ↑ 232% (p ≤ 0.05) | ||
| RET young: ↑ 187% (p ≤ 0.05) | |||
| RET older: ↑ 243% (p ≤ 0.05) | |||
| CT young: ↑ 158% (p ≤ 0.05) | |||
| CT older: ↑ 177% (p ≤ 0.05) | |||
| LUM | HIIT young: (NS) | ||
| HIIT older: ↑ 156% (p ≤ 0.05) | |||
| RET young: (NS) | |||
| RET older: (NS) | |||
| CT young: (NS) | |||
| CT older: (NS) | |||
| ECM2 | HIIT young: ↑ 175% (p ≤ 0.05) | ||
| HIIT older: ↑ 180% (p ≤ 0.05) | |||
| RET young: ↑161% (p ≤ 0.05) | |||
| RET older: ↑173% (p ≤ 0.05) | |||
| CT young: (NS) | |||
| CT older: (NS) | |||
| Glycoproteins | |||
| LAMB1 | HIIT young: ↑ 171% (p ≤ 0.05) | ||
| HIIT older: ↑ 170% (p ≤ 0.05) | |||
| RET young: ↑ 154% (p ≤ 0.05) | |||
| RET older: (NS) | |||
| CT young: (NS) | |||
| CT older: ↑ 160% (p ≤ 0.05) | |||
| NID1 | HIIT young: ↑ 160% (p ≤ 0.05) | ||
| HIIT older: ↑ 205% (p ≤ 0.05) | |||
| RET young: ↑152% (p ≤ 0.05) | |||
| RET older: ↑157% (p ≤ 0.05) | |||
| CT young: ↑155% (p ≤ 0.05) | |||
| CT older: ↑156% (p ≤ 0.05) | |||
| PXDN | HIIT young: ↑ 196% (p ≤ 0.05) | ||
| HIIT older: ↑266% (p ≤ 0.05) | |||
| RET young: ↑234% (p ≤ 0.05) | |||
| RET older: ↑209% (p ≤ 0.05) | |||
| CT young: ↑162% (p ≤ 0.05) | |||
| CT older: ↑166% (p ≤ 0.05) | |||
| SPARC | HIIT young: ↑ 188% (p ≤ 0.05) | ||
| HIIT older: ↑224% (p ≤ 0.05) | |||
| RET young: ↑170% (p ≤ 0.05) | |||
| RET older: ↑179% (p ≤ 0.05) | |||
| CT young: (NS) | |||
| CT older: ↑165% (p ≤ 0.05) | |||
| ELN | HIIT young: (NS) | ||
| HIIT older: ↑181% (p ≤ 0.05) | |||
| RET young: (NS) | |||
| RET older: (NS) | |||
| CT young: (NS) | |||
| CT older: (NS) | |||
| POSTN | HIIT young: (NS) | ||
| HIIT older: ↑ 167% (p ≤ 0.05) | |||
| RET young: (NS) | |||
| RET older: (NS) | |||
| CT young: (NS) | |||
| CT older: (NS) | |||
| Timmons et al., 2010 [39] | mRNA expression | N/A | |
| Collagens | |||
| COL1A1 | ↑ 370–510% | ||
| COL1A2 | ↑ 90–550% | ||
| COL3A1 | ↑ 80–540% | ||
| COL4A1 | ↑ 350–430% | ||
| COL4A2 | ↑ 280–350% | ||
| COL5A1 | ↑ 240–270% | ||
| COL5A2 | ↑ 250–290% | ||
| COL5A3 | ↑ 50% | ||
| COL6A1 | ↑ 70% | ||
| COL6A2 | ↑ 210% | ||
| COL6A3 | ↑ 230% | ||
| COL8A1 | ↑ 260% | ||
| COL12A1 | ↑ 50% | ||
| COL14A1 | ↑ 250% | ||
| COL15A1 | ↑ 80% | ||
| COL18A1 | ↑ 60% | ||
| PLOD2 | ↑ 60% | ||
| Proteoglycans | |||
| ASPN | ↑ 220–290% | ||
| BGN | ↑ 250–560% | ||
| CSPG2 | ↑ 170–590% | ||
| HSPG2 | ↑ 90% | ||
| LUM | ↑ 270% | ||
| OGN | ↑ 420–500% | ||
| Glycoproteins | |||
| AGRN | ↑ 60–80% | ||
| LAMA4 | ↑ 240% | ||
| LAMB1 | ↑ 250–270% | ||
| LAMC1 | ↑ 50% | ||
| SPARC | ↑ 50–250% | ||
| NID1 | ↑ 80% | ||
| NID2 | ↑ 290% | ||
| FBN1 | ↑ 50–730% | ||
| FN1 | ↑ 60–90% | ||
| TNC | ↑ 430–530% | ||
| THBS4 | ↑ 280% | ||
| POSTN | ↑ 390% | ||
| PXDN | ↑ 220–250% | ||
| FNDC1 | ↑ 300% | ||
| CTHRC1 | ↑ 340% | ||
| Valdivierso et al., 2017 [40] | Protein expression | ||
| Glycoproteins | |||
| Tenascin C | A/A alleles: ↑ 138% (p < 0.05) | Capillary/fibre ↑ | |
| A/T alleles: ↑ 77% (p < 0.05) | Capillary/fibre ↑ | ||
| T/T alleles: (NS) | Capillary/fibre ↓ 15% | ||
| Walton et al., 2019 [41] | mRNA expression | M2 macrophages/fibre vs. COL5A1 expression (r = 0.56, p = 0.021) | |
| Collagens | M2 macrophages/fibre vs. COL6A1 expression (r = 0.54, p = 0.026) | ||
| COL5A1 | ↑ 53.6% (p = 0.013) | M2 macrophages/fibre vs. SPARC expression (r = 0.63, p = 0.007) | |
| COL6A1 | ↑ 29.5% (p = 0.009) | M2 macrophages/fibre vs. MMP14 expression (r = 0.69, p = 0.002) | |
| Glycoproteins | M2 macrophages/fibre vs. TGFβ1 expression (r = 0.50, p = 0.04) | ||
| SPARC | ↑ 56.6% (p < 0.001) | ||
| Alghadir et al., 2016 [42] | Circulating markers (serum) | Level of cFN vs. Physical activity (PA): | |
| Cellular Fibronectin (or cFN) | Exercise: ↓ 53% (p < 0.001) | ↑ in cFN after in low PA (r = 0.18, p < 0.001) | |
| Control: ↓ 2.1% (NS) | ↑ in cFN after moderate PA (r = 0.12, p < 0.001) | ||
| ↑ in cFN after ↑ in high PA vs. (r = 0.14, p < 0.001) | |||
| Olstad et al., 2020 [34] | mRNA expression | N/A | |
| Proteoglycans | |||
| DCN | Healthy: N/A | ||
| Osteoporotic: ↑ 129.4% | |||
| Glycoproteins | |||
| SPARC | Healthy: N/A | ||
| Osteoporotic: ↑ 141.6% | |||
| MGP | Healthy: N/A | ||
| Osteoporotic: ↑ 128.2% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kritikaki, E.; Asterling, R.; Ward, L.; Padget, K.; Barreiro, E.; C. M. Simoes, D. Exercise Training-Induced Extracellular Matrix Protein Adaptation in Locomotor Muscles: A Systematic Review. Cells 2021, 10, 1022. https://doi.org/10.3390/cells10051022
Kritikaki E, Asterling R, Ward L, Padget K, Barreiro E, C. M. Simoes D. Exercise Training-Induced Extracellular Matrix Protein Adaptation in Locomotor Muscles: A Systematic Review. Cells. 2021; 10(5):1022. https://doi.org/10.3390/cells10051022
Chicago/Turabian StyleKritikaki, Efpraxia, Rhiannon Asterling, Lesley Ward, Kay Padget, Esther Barreiro, and Davina C. M. Simoes. 2021. "Exercise Training-Induced Extracellular Matrix Protein Adaptation in Locomotor Muscles: A Systematic Review" Cells 10, no. 5: 1022. https://doi.org/10.3390/cells10051022
APA StyleKritikaki, E., Asterling, R., Ward, L., Padget, K., Barreiro, E., & C. M. Simoes, D. (2021). Exercise Training-Induced Extracellular Matrix Protein Adaptation in Locomotor Muscles: A Systematic Review. Cells, 10(5), 1022. https://doi.org/10.3390/cells10051022