
Food Limitation but Not Enhanced Rates of Ejaculate Production Imposes Reproductive and Survival Costs to Male Crickets


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Rearing
2.2. Experimental Treatments
2.3. Body Mass Measures
2.4. Spermatophore Production and Sperm Viability
2.5. Longevity and Lifetime Survival
2.6. Statistical Analysis
3. Results
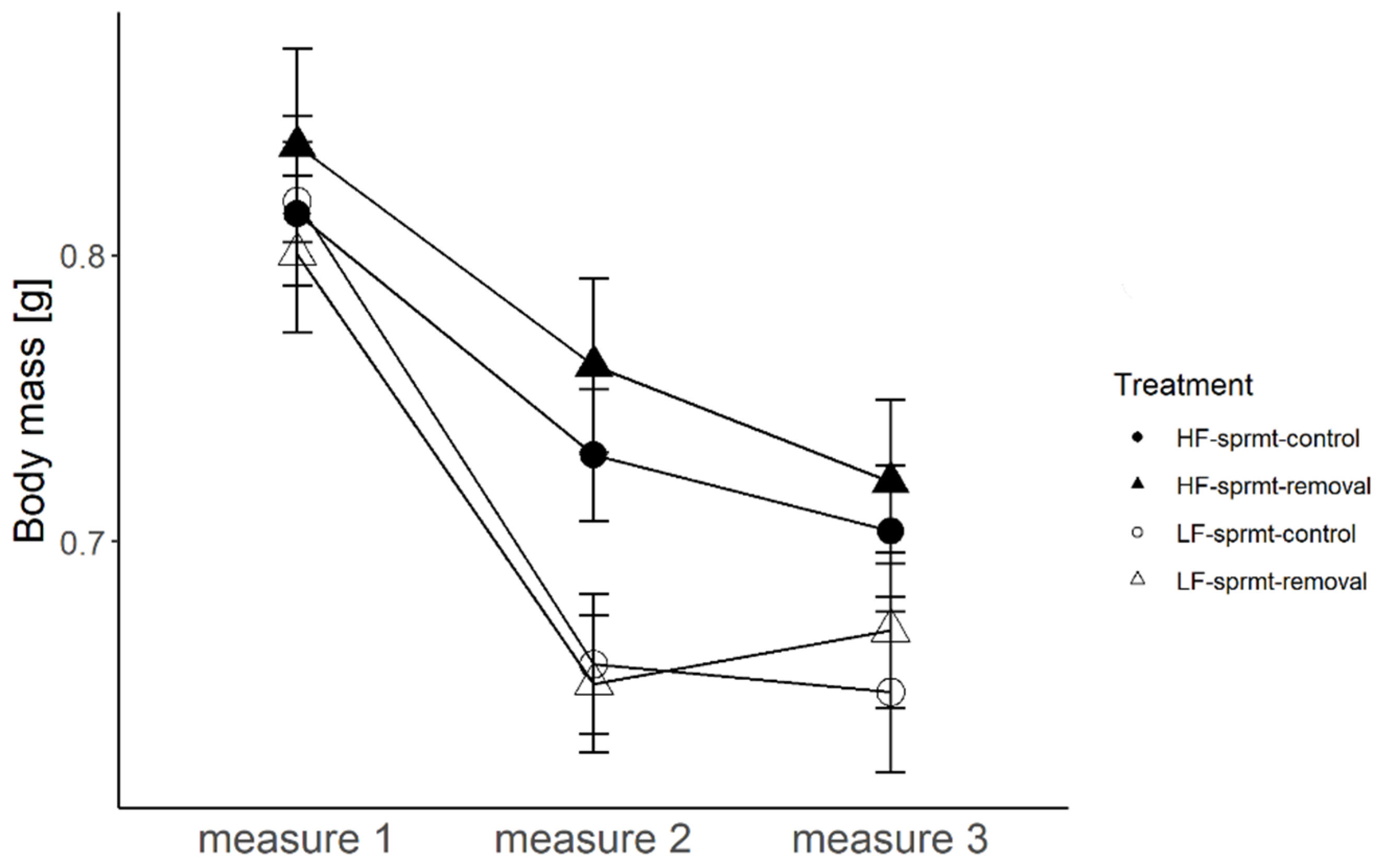
3.1. Male Body Mass
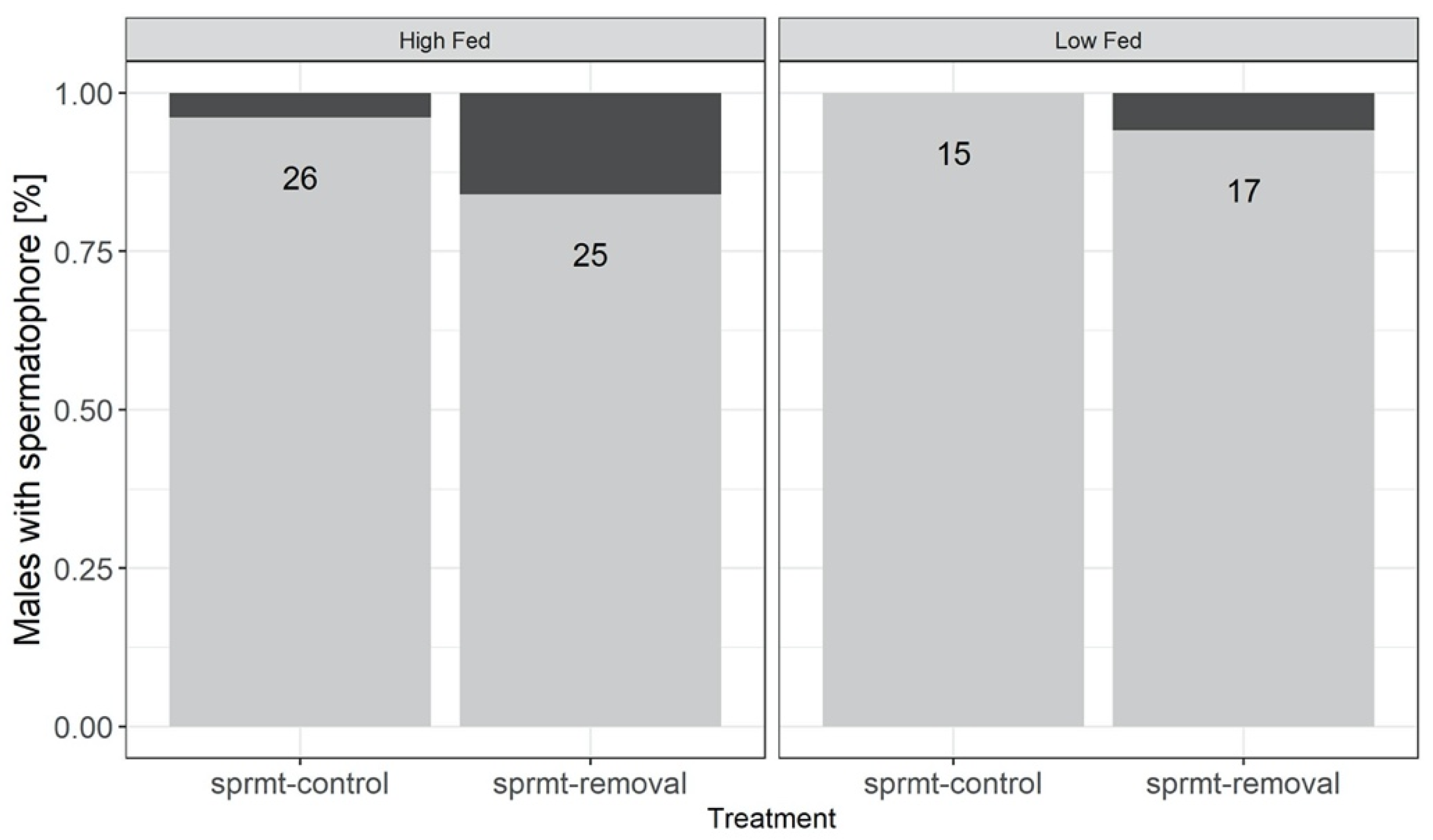
3.2. Spermatophore Production and Sperm Viability
3.3. Longevity and Lifetime Survival
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Hayward, A.; Gillooly, J.F. The Cost of Sex: Quantifying Energetic Investment in Gamete Production by Males and Females. PLoS ONE 2011, 6, e16557. [Google Scholar] [CrossRef] [PubMed]
- Dewsbury, D.A. Ejaculate Cost and Male Choice. Am. Nat. 1982, 119, 601–610. [Google Scholar] [CrossRef]
- Perry, J.C.; Tse, C.T. Extreme Costs of Mating for Male Two-Spot Ladybird Beetles. PLoS ONE 2013, 8, e81934. [Google Scholar] [CrossRef] [PubMed]
- Godwin, J.L.; Vasudeva, R.; Michalczyk, Ł.; Martin, O.Y.; Lumley, A.J.; Chapman, T.; Gage, M.J.G. Experimental Evolution Reveals That Sperm Competition Intensity Selects for Longer, More Costly Sperm. Evol. Lett. 2017, 1, 102–113. [Google Scholar] [CrossRef] [Green Version]
- Van Voorhies, W.A. Production of Sperm Reduces Nematode Lifespan. Nature 1992, 360, 456–458. [Google Scholar] [CrossRef]
- Avila, F.W.; Sirot, L.K.; LaFlamme, B.A.; Rubinstein, C.D.; Wolfner, M.F. Insect Seminal Fluid Proteins: Identification and Function. Annu. Rev. Entomol. 2011, 56, 21. [Google Scholar] [CrossRef] [Green Version]
- Poiani, A. Complexity of Seminal Fluid: A Review. Behav. Ecol. Sociobiol. 2006, 60, 289–310. [Google Scholar] [CrossRef]
- Simmons, L.W.; Fitzpatrick, J.L. Sperm Wars and the Evolution of Male Fertility. Reproduction 2012, 144, 519. [Google Scholar] [CrossRef]
- Fitzpatrick, J.L.; Lüpold, S. Sexual Selection and the Evolution of Sperm Quality. Mol. Hum. Reprod. 2014, 20, 1180–1189. [Google Scholar] [CrossRef] [Green Version]
- Parker, G.A. Sperm Competition and Its Evolutionary Consequences in the Insects. Biol. Rev. 1970, 45, 525–567. [Google Scholar] [CrossRef]
- Parker, G.A. The Evolution of Expenditure on Testes. J. Zool. 2016, 298, 3–19. [Google Scholar] [CrossRef]
- Thomsen, R.; Soltis, J.; Matsubara, M.; Matsubayashi, K.; Onuma, M.; Takenaka, O. How Costly Are Ejaculates for Japanese Macaques? Primates 2006, 47, 272–274. [Google Scholar] [CrossRef]
- Friesen, C.R.; Shine, R.; Krohmer, R.W.; Mason, R.T. Not Just a Chastity Belt: The Functional Significance of Mating Plugs in Garter Snakes, Revisited. Biol. J. Linn. Soc. 2013, 109, 893–907. [Google Scholar] [CrossRef] [Green Version]
- Colpo, K.D.; López-Greco, L.S. Dynamics of Energy Reserves and the Cost of Reproduction in Female and Male Fiddler Crabs. Zoology 2018, 126, 11–19. [Google Scholar] [CrossRef]
- Voigt, C.C.; Michener, R.; Kunz, T.H. The Energetics of Trading Nuptial Gifts for Copulations in Katydids. Physiol. Biochem. Zool. 2005, 78, 417–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scharf, I.; Peter, F.; Martin, O.Y. Reproductive Trade-Offs and Direct Costs for Males in Arthropods. Evol. Biol. 2013, 40, 169–184. [Google Scholar] [CrossRef]
- Reznick, D. Measuring the Costs of Reproduction. Trends Ecol. Evol. 1992, 7, 42–45. [Google Scholar] [CrossRef]
- Roff, D.A. Life History Evolution. Recherche 2002, 67, 2. [Google Scholar]
- Stearns, S.C. The Evolution of Life Histories; Oxford University Press: Oxford, UK, 1992; Volume 249. [Google Scholar]
- Williams, G.C. Natural Selection, the Costs of Reproduction, and a Refinement of Lack’s Principle. Am. Nat. 1966, 100, 687–690. [Google Scholar] [CrossRef] [Green Version]
- Sella, G.; Lorenzi, M.C. Increased Sperm Allocation Delays Body Growth in a Protandrous Simultaneous Hermaphrodite. Biol. J. Linn. Soc. 2003, 78, 149–154. [Google Scholar] [CrossRef]
- Olsson, M.; Madsen, T.; Shine, R. Is Sperm Really so Cheap? Costs of Reproduction in Male Adders, Vipera Berus. Proc. R. Soc. Lond. B Biol. Sci. 1997, 264, 455–459. [Google Scholar] [CrossRef] [Green Version]
- Schwenke, R.A.; Lazzaro, B.P.; Wolfner, M.F. Reproduction–Immunity Trade-Offs in Insects. Annu. Rev. Entomol. 2016, 61, 239–256. [Google Scholar] [CrossRef] [Green Version]
- Simmons, L.W.; Lüpold, S.; Fitzpatrick, J.L. Evolutionary Trade-off between Secondary Sexual Traits and Ejaculates. Trends Ecol. Evol. 2017, 32, 964–976. [Google Scholar] [CrossRef] [PubMed]
- Rönn, J.L.; Katvala, M.; Arnqvist, G. Interspecific Variation in Ejaculate Allocation and Associated Effects on Female Fitness in Seed Beetles. J. Evol. Biol. 2008, 21, 461–470. [Google Scholar] [CrossRef] [Green Version]
- Torres-Vila, L.M.; Jennions, M.D. Male Mating History and Female Fecundity in the Lepidoptera: Do Male Virgins Make Better Partners? Behav. Ecol. Sociobiol. 2005, 57, 318–326. [Google Scholar] [CrossRef]
- Bailey, W.J.; Nuhardiyati, M. Copulation, the Dynamics of Sperm Transfer and Female Refractoriness in the Leafhopper Balclutha Incisa (Hemiptera: Cicadellidae: Deltocephalinae). Physiol. Entomol. 2005, 30, 343–352. [Google Scholar] [CrossRef]
- Reinhardt, K.; Naylor, R.; Siva-Jothy, M.T. Male Mating Rate Is Constrained by Seminal Fluid Availability in Bedbugs, Cimex Lectularius. PLoS ONE 2011, 6, e22082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Noordwijk, A.J.; de Jong, G. Acquisition and Allocation of Resources: Their Influence on Variation in Life History Tactics. Am. Nat. 1986, 128, 137–142. [Google Scholar] [CrossRef] [Green Version]
- Reznick, D.; Nunney, L.; Tessier, A. Big Houses, Big Cars, Superfleas and the Costs of Reproduction. Trends Ecol. Evol. 2000, 15, 421–425. [Google Scholar] [CrossRef]
- Boggs, C.L. Understanding Insect Life Histories and Senescence through a Resource Allocation Lens. Funct. Ecol. 2009, 23, 27–37. [Google Scholar] [CrossRef]
- Simmons, L.W. Resource Allocation Trade-off between Sperm Quality and Immunity in the Field Cricket, Teleogryllus Oceanicus. Behav. Ecol. 2012, 23, 168–173. [Google Scholar] [CrossRef] [Green Version]
- Zera, A.J.; Harshman, L.G. The Physiology of Life History Trade-Offs in Animals. Annu. Rev. Ecol. Syst. 2001, 32, 95–126. [Google Scholar] [CrossRef] [Green Version]
- Duffield, K.R.; Bowers, E.K.; Sakaluk, S.K.; Sadd, B.M. A Dynamic Threshold Model for Terminal Investment. Behav. Ecol. Sociobiol. 2017, 71, 1–17. [Google Scholar] [CrossRef]
- French, S.S.; DeNardo, D.F.; Moore, M.C. Trade-Offs between the Reproductive and Immune Systems: Facultative Responses to Resources or Obligate Responses to Reproduction? Am. Nat. 2007, 170, 79–89. [Google Scholar] [CrossRef] [PubMed]
- King, E.G.; Roff, D.A.; Fairbairn, D.J. The Evolutionary Genetics of Acquisition and Allocation in the Wing Dimorphic Cricket, Gryllus Firmus. Evol. Int. J. Org. Evol. 2011, 65, 2273–2285. [Google Scholar] [CrossRef]
- Simmons, L.W.; Roberts, B. Bacterial Immunity Traded for Sperm Viability in Male Crickets. Science 2005, 309, 2031. [Google Scholar] [CrossRef] [PubMed]
- Kerr, A.M.; Gershman, S.N.; Sakaluk, S.K. Experimentally Induced Spermatophore Production and Immune Responses Reveal a Trade-off in Crickets. Behav. Ecol. 2010, 21, 647–654. [Google Scholar] [CrossRef]
- McNamara, K.B.; Van Lieshout, E.; Jones, T.M.; Simmons, L.W. Age-dependent Trade-offs between Immunity and Male, but Not Female, Reproduction. J. Anim. Ecol. 2013, 82, 235–244. [Google Scholar] [CrossRef]
- Stürup, M.; Baer, B.; Boomsma, J.J. Short Independent Lives and Selection for Maximal Sperm Survival Make Investment in Immune Defences Unprofitable for Leaf-Cutting Ant Males. Behav. Ecol. Sociobiol. 2014, 68, 947–955. [Google Scholar] [CrossRef]
- Hall, M.D.; Beck, R.; Greenwood, M. Detailed Developmental Morphology of the Spermatophore of the Mediterranean Field Cricket, Gryllus Bimaculatus (De Geer) (Orthoptera: Gryllidae). Arthropod Struct. Dev. 2000, 29, 23–32. [Google Scholar] [CrossRef]
- Sakai, M.; Taoda, Y.; Mori, K.; Fujino, M.; Ohta, C. Copulation Sequence and Mating Termination in the Male Cricket Gryllus Bimaculatus DeGeer. J. Insect Physiol. 1991, 37, 599–605, 607–615. [Google Scholar] [CrossRef]
- Tuni, C.; Mizerakis, V.; Dingemanse, N.J. Experimental Evidence That Winning or Losing a Fight Does Not Affect Sperm Quality in a Field Cricket. Ethology 2019, 125, 885–889. [Google Scholar] [CrossRef]
- Snook, R.R. Sperm in Competition: Not Playing by the Numbers. Trends Ecol. Evol. 2005, 20, 46–53. [Google Scholar] [CrossRef] [PubMed]
- García-González, F.; Simmons, L.W. Sperm Viability Matters in Insect Sperm Competition. Curr. Biol. 2005, 15, 271–275. [Google Scholar] [CrossRef] [Green Version]
- Fry, C.L.; Wilkinson, G.S. Sperm Survival in Female Stalk-eyed Flies Depends on Seminal Fluid and Meiotic Drive. Evolution 2004, 58, 1622–1626. [Google Scholar] [CrossRef]
- Hunter, F.M.; Birkhead, T.R. Sperm Viability and Sperm Competition in Insects. Curr. Biol. 2002, 12, 121–123. [Google Scholar] [CrossRef] [Green Version]
- Gomendio, M.; Martin-Coello, J.; Crespo, C.; Magaña, C.; Roldan, E.R.S. Sperm Competition Enhances Functional Capacity of Mammalian Spermatozoa. Proc. Natl. Acad. Sci. USA 2006, 103, 15113–15117. [Google Scholar] [CrossRef] [Green Version]
- den Boer, S.P.A.; Boomsma, J.J.; Baer, B. Honey Bee Males and Queens Use Glandular Secretions to Enhance Sperm Viability before and after Storage. J. Insect Physiol. 2009, 55, 538–543. [Google Scholar] [CrossRef] [Green Version]
- Bretman, A.; Tregenza, T. Measuring Polyandry in Wild Populations: A Case Study Using Promiscuous Crickets. Mol. Ecol. 2005, 14, 2169–2179. [Google Scholar] [CrossRef]
- delBarco-Trillo, J.; García-Álvarez, O.; Soler, A.J.; Tourmente, M.; Garde, J.J.; Roldan, E.R.S. A Cost for High Levels of Sperm Competition in Rodents: Increased Sperm DNA Fragmentation. Proc. R. Soc. B Biol. Sci. 2016, 283, 20152708. [Google Scholar] [CrossRef] [PubMed]
- Montrose, V.T.; Harris, W.E.; Moore, P.J. Sexual Conflict and Cooperation under Naturally Occurring Male Enforced Monogamy. J. Evol. Biol. 2004, 17, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Macartney, E.L.; Crean, A.J.; Nakagawa, S.; Bonduriansky, R. Effects of Nutrient Limitation on Sperm and Seminal Fluid: A Systematic Review and Meta-Analysis. Biol. Rev. 2019, 94, 1722–1739. [Google Scholar] [CrossRef]
- Scheuber, H.; Jacot, A.; Brinkhof, M.W.G. The Effect of Past Condition on a Multicomponent Sexual Signal. Proc. R. Soc. Lond. B Biol. Sci. 2003, 270, 1779–1784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagao, T.; Shimozawa, T. A Fixed Time-Interval between Two Behavioural Elements in the Mating Behaviour of Male Crickets, Gryllus Bimaculatus. Anim. Behav. 1987, 35, 122–130. [Google Scholar] [CrossRef]
- Ootsubo, T.; Sakai, M. Initiation of Spermatophore Protrusion Behavior in the Male Cricket Gryllus Bimaculatus DeGeer (Physology). Zool. Sci. 1992, 9, 955–969. [Google Scholar]
- Kumashiro, M.; Tsuji, Y.; Sakai, M. Auto-Spermatophore Extrusion in Male Crickets. J. Exp. Biol. 2003, 206, 4507–4519. [Google Scholar] [CrossRef] [Green Version]
- Loher, W. Circadian Control of Spermatophore Formation in the Cricket Teleogryllus Commodus Walker. J. Insect Physiol. 1974, 20, 1155–1172. [Google Scholar] [CrossRef]
- Reinhardt, K.; Siva-Jothy, M.T. An Advantage for Young Sperm in the House Cricket Acheta Domesticus. Am. Nat. 2005, 165, 718–723. [Google Scholar] [CrossRef] [PubMed]
- Tuni, C.; Perdigón Ferreira, J.; Fritz, Y.; Munoz Meneses, A.; Gasparini, C. Impaired Sperm Quality, Delayed Mating but No Costs for Offspring Fitness in Crickets Winning a Fight. J. Evol. Biol. 2016, 29, 1643–1647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasparini, C.; Lu, C.; Dingemanse, N.J.; Tuni, C. Paternal-effects in a Terrestrial Ectotherm Are Temperature Dependent but No Evidence for Adaptive Effects. Funct. Ecol. 2018, 32, 1011–1021. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Multimodel Inference: Understanding AIC and BIC in Model Selection. Sociol. Methods Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- Harrison, X.A. A Comparison of Observation-Level Random Effect and Beta-Binomial Models for Modelling Overdispersion in Binomial Data in Ecology & Evolution. PeerJ 2015, 3, e1114. [Google Scholar] [PubMed] [Green Version]
- McNamara, K.B.; Elgar, M.A.; Jones, T.M. A Longevity Cost of Re-Mating but No Benefits of Polyandry in the Almond Moth, Cadra Cautella. Behav. Ecol. Sociobiol. 2008, 62, 1433–1440. [Google Scholar] [CrossRef]
- Martin, O.Y.; Hosken, D.J. Copulation Reduces Male but Not Female Longevity in Saltella Sphondylli (Diptera: Sepsidae). J. Evol. Biol. 2004, 17, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Wagner, W.E., Jr.; Reiser, M.G. The Importance of Calling Song and Courtship Song in Female Mate Choice in the Variable Field Cricket. Anim. Behav. 2000, 59, 1219–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holzer, B.; Jacot, A.; Brinkhof, M.W. Condition-Dependent Signaling Affects Male Sexual Attractiveness in Field Crickets, Gryllus Campestris. Behav. Ecol. 2003, 14, 353–359. [Google Scholar] [CrossRef] [Green Version]
- Scheuber, H.; Jacot, A.; Brinkhof, M.W. Condition Dependence of a Multicomponent Sexual Signal in the Field Cricket Gryllus Campestris. Anim. Behav. 2003, 65, 721–727. [Google Scholar] [CrossRef]
- Simmons, L.W. Female Choice in the Field Cricket Gryllus Bimaculatus (De Geer). Anim. Behav. 1986, 34, 1463–1470. [Google Scholar] [CrossRef]
- Adamo, S.A.; Hoy, R.R. Agonistic Behaviour in Male and Female Field Crickets, Gryllus Bimaculatus, and How Behavioural Context Influences Its Expression. Anim. Behav. 1995, 49, 1491–1501. [Google Scholar] [CrossRef]
- Rowe, L.; Houle, D. The Lek Paradox and the Capture of Genetic Variance by Condition Dependent Traits. Proc. R. Soc. Lond. B Biol. Sci. 1996, 263, 1415–1421. [Google Scholar]
- Cotton, S.; Fowler, K.; Pomiankowski, A. Do Sexual Ornaments Demonstrate Heightened Condition-Dependent Expression as Predicted by the Handicap Hypothesis? Proc. R. Soc. Lond. B Biol. Sci. 2004, 271, 771–783. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.M.; Kelley, J.L.; Evans, J.P. Condition-Dependent Expression of Pre- and Postcopulatory Sexual Traits in Guppies. Ecol. Evol. 2013, 3, 2197–2213. [Google Scholar] [CrossRef]
- Perry, J.C.; Rowe, L. Condition-Dependent Ejaculate Size and Composition in a Ladybird Beetle. Proc. R. Soc. Lond. B Biol. Sci. 2010, 277, 3639–3647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehlis, M.; Rick, I.P.; Bakker, T.C.M. Dynamic Resource Allocation between Pre-and Postcopulatory Episodes of Sexual Selection Determines Competitive Fertilization Success. Proc. R. Soc. B Biol. Sci. 2015, 282, 20151279. [Google Scholar] [CrossRef]
- Stürup, M.; Baer-Imhoof, B.; Nash, D.R.; Boomsma, J.J.; Baer, B. When Every Sperm Counts: Factors Affecting Male Fertility in the Honeybee Apis Mellifera. Behav. Ecol. 2013, 24, 1192–1198. [Google Scholar] [CrossRef] [Green Version]
- Dávila, F.; Aron, S. Protein Restriction Affects Sperm Number but Not Sperm Viability in Male Ants. J. Insect Physiol. 2017, 100, 71–76. [Google Scholar] [CrossRef]
- Bunning, H.; Rapkin, J.; Belcher, L.; Archer, C.R.; Jensen, K.; Hunt, J. Protein and Carbohydrate Intake Influence Sperm Number and Fertility in Male Cockroaches, but Not Sperm Viability. Proc. R. Soc. B Biol. Sci. 2015, 282, 20142144. [Google Scholar] [CrossRef]
- Joseph, P.N.; Sasson, D.A.; Allen, P.E.; Somjee, U.; Miller, C.W. Adult Nutrition, but Not Inbreeding, Affects Male Primary Sexual Traits in the Leaf-footed Cactus Bug Narnia Femorata (Hemiptera: Coreidae). Ecol. Evol. 2016, 6, 4792–4799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duxbury, A.E.; Weathersby, B.; Sanchez, Z.; Moore, P.J. A Study of the Transit Amplification Divisions during Spermatogenesis in Oncopetus Fasciatus to Assess Plasticity in Sperm Numbers or Sperm Viability under Different Diets. Ecol. Evol. 2018, 8, 10460–10469. [Google Scholar] [CrossRef] [Green Version]
- Ng, S.H.; Simpson, S.J.; Simmons, L.W. Macronutrients and Micronutrients Drive Trade-offs between Male Pre-and Postmating Sexual Traits. Funct. Ecol. 2018, 32, 2380–2394. [Google Scholar] [CrossRef]
- Svärd, L.; Wiklund, C. Mass and Production Rate of Ejaculates in Relation to Monandry/Polyandry in Butterflies. Behav. Ecol. Sociobiol. 1989, 24, 395–402. [Google Scholar] [CrossRef]
- Simmons, L.W. Male Size, Mating Potential and Lifetime Reproductive Success in the Field Cricket, Gryllus Bimaculatus (De Geer). Anim. Behav. 1988, 36, 372–379. [Google Scholar] [CrossRef]
- Zuk, M. The Effects of Gregarine Parasites, Body Size, and Time of Day on Spermatophore Production and Sexual Selection in Field Crickets. Behav. Ecol. Sociobiol. 1987, 21, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Sturm, R. Comparison of Sperm Number, Spermatophore Size, and Body Size in Four Cricket Species Comparison of Sperm Number, Spermatophore Size, and Body Size in Four Cricket Species. J. Orthoptera Res. 2014, 23, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Vahed, K. The Function of Nuptial Feeding in Insects: A Review of Empirical Studies. Biol. Rev. 1998, 73, 43–78. [Google Scholar] [CrossRef]
- Lehmann, G.U.C.; Lehmann, K.; Neumann, B.; Lehmann, A.W.; Scheler, C.; Jungblut, P.R. Protein Analysis of the Spermatophore Reveals Diverse Compositions in Both the Ampulla and the Spermatophylax in a Bushcricket. Physiol. Entomol. 2018, 43, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Mccartney, J.; Potter, M.A.; Robertson, A.W.; Telscher, K.; Lehmann, G.; Lehmann, A.; Von-Helversen, D.; Reinhold, K.; Achmann, R.; Heller, K.-G. Understanding Nuptial Gift Size in Bush-Crickets: An Analysis of the Genus Poecilimon (Tettigoniidae: Orthoptera). J. Orthoptera Res. 2008, 17, 231–242. [Google Scholar] [CrossRef]
- Sevgili, H. Spermatophore Weight and Sperm Number in the Southernmost Species of the Genus Poecilimon Fischer, 1853 (Orthoptera: Phaneropterinae). Acta Biologica Turcica 2016, 29, 83–89. [Google Scholar]
- Sakaluk, S.K. Spermatophore Size and Its Role in the Reproductive Behaviour of the Cricket, Gryllodes Supplicans (Orthoptera: Gryllidae). Can. J. Zool. 1985, 63, 1652–1656. [Google Scholar] [CrossRef]
- Simmons, L.W.; Beveridge, M. Seminal Fluid Affects Sperm Viability in a Cricket. PLoS ONE 2011, 6, e17975. [Google Scholar] [CrossRef] [Green Version]
- Kaldun, B.; Otti, O. Condition-dependent Ejaculate Production Affects Male Mating Behavior in the Common Bedbug Cimex Lectularius. Ecol. Evol. 2016, 6, 2548–2558. [Google Scholar] [CrossRef] [Green Version]
- Clutton-Brock, T.H.; Parker, G.A. Potential Reproductive Rates and the Operation of Sexual Selection. Q. Rev. Biol. 1992, 67, 437–456. [Google Scholar] [CrossRef]
- Nakatsuru, K.E.N.; Kramer, D.L. Is Sperm Cheap? Limited Male Fertility and Female Choice in the Lemon Tetra (Pisces, Characidae). Science 1982, 216, 753–755. [Google Scholar] [CrossRef] [PubMed]
- Preston, B.T.; Stevenson, I.R.; Pemberton, J.M.; Wilson, K. Dominant Rams Lose out by Sperm Depletion. Nature 2001, 409, 681–682. [Google Scholar] [CrossRef]
- Hettyey, A.; Vagi, B.; Hevizi, G.; Toeroek, J. Changes in Sperm Stores, Ejaculate Size, Fertilization Success, and Sexual Motivation over Repeated Matings in the Common Toad, Bufo Bufo (Anura: Bufonidae). Biol. J. Linn. Soc. 2009, 96, 361–371. [Google Scholar] [CrossRef] [Green Version]
- Sturm, R. The Effect of Remating on Sperm Number in the Spermatophores of Teleogryllus Commodus (Gryllidae). Invertebr. Biol. 2011, 130, 362–367. [Google Scholar] [CrossRef]
- Sirot, L.K.; Buehner, N.A.; Fiumera, A.C.; Wolfner, M.F. Seminal Fluid Protein Depletion and Replenishment in the Fruit Fly, Drosophila Melanogaster: An ELISA-Based Method for Tracking Individual Ejaculates. Behav. Ecol. Sociobiol. 2009, 63, 1505–1513. [Google Scholar] [CrossRef] [Green Version]
- Radhakrishnan, P.; Taylor, P.W. Ability of Male Queensland Fruit Flies to Inhibit Receptivity in Multiple Mates, and the Associated Recovery of Accessory Glands. J. Insect Physiol. 2008, 54, 421–428. [Google Scholar] [CrossRef]
- Abraham, S.; Moyano, A.; Murillo Dasso, S.; Van Nieuwenhove, G.; Ovruski, S.; Pérez-Staples, D. Male Accessory Gland Depletion in a Tephritid Fly Affects Female Fecundity Independently of Sperm Depletion. Behav. Ecol. Sociobiol. 2020, 74, 1–9. [Google Scholar] [CrossRef]
- Simmons, L.W.; Moore, A.J. Evolutionary quantitative genetics of sperm. In Sperm Biology: An Evolutionary Perspective; Birkhead, T.R., Hosken, D.J., Pitnick, S., Eds.; Academic Press: Burlington, NJ, USA, 2008; pp. 405–434. [Google Scholar]
- Miller, L.K.; Brooks, R. The Effects of Genotype, Age, and Social Environment on Male Ornamentation, Mating Behavior, and Attractiveness. Evolution 2005, 59, 2414–2425. [Google Scholar] [CrossRef]
- Gems, D.; Riddle, D.L. Longevity in Caenorhabditis Elegans Reduced by Mating but Not Gamete Production. Nature 1996, 379, 723–725. [Google Scholar] [CrossRef]
- Wedell, N.; Gage, M.J.G.; Parker, G.A. Sperm Competition, Male Prudence and Sperm-Limited Females. Trends Ecol. Evol. 2002, 17, 313–320. [Google Scholar] [CrossRef]
- Cornwallis, C.K.; Birkhead, T.R. Social Status and Availability of Females Determine Patterns of Sperm Allocation in the Fowl. Evolution 2006, 60, 1486–1493. [Google Scholar] [CrossRef]
- Warner, R.R.; Shapiro, D.Y.; Marcanato, A.; Petersen, C.W. Sexual Conflict: Males with Highest Mating Success Convey the Lowest Fertilization Benefits to Females. Proc. R. Soc. Lond. B Biol. Sci. 1995, 262, 135–139. [Google Scholar]
- Clutton-Brock, T.H. Reproductive Effort and Terminal Investment in Iteroparous Animals. Am. Nat. 1984, 123, 212–229. [Google Scholar] [CrossRef]
- Candolin, U. The Relationship between Signal Quality and Physical Condition: Is Sexual Signalling Honest in the Three-Spined Stickleback? Anim. Behav. 1999, 58, 1261–1267. [Google Scholar] [CrossRef] [Green Version]
- O’Dea, R.E.; Jennions, M.D.; Head, M.L. Male Body Size and Condition Affects Sperm Number and Production Rates in Mosquitofish, Gambusia Holbrooki. J. Evol. Biol. 2014, 27, 2739–2744. [Google Scholar] [CrossRef] [PubMed]
- Ollero, M.; Muiño-Blanco, T.; López-Pérez, M.J.; Cebrián-Pérez, J.A. Viability of Ram Spermatozoa in Relation to the Abstinence Period and Successive Ejaculations. Int. J. Androl. 1996, 19, 287–292. [Google Scholar] [CrossRef]
- Reinhardt, K. Evolutionary Consequences of Sperm Cell Aging. Q. Rev. Biol. 2007, 82, 375–393. [Google Scholar] [CrossRef]
- Gasparini, C.; Dosselli, R.; Evans, J.P. Sperm Storage by Males Causes Changes in Sperm Phenotype and Influences the Reproductive Fitness of Males and Their Sons. Evol. Lett. 2017, 1, 16–25. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Response Variable | Effect (Wald X2 or F; df; P) | ||||
|---|---|---|---|---|---|
| N | Food Treatment | Spermatophore Removal Treatment | Food x Spermatophore Treatment | Timepoint of Measure | |
| Body mass 1 | 188 | 18.78; 1; <0.0001 | 0.0006; 1; 0.98 | 0.0001; 1; 0.99 | 41.53; 1; <0.0001 |
| Spermatophore production (% males) 2 | 84 | 20.6; 1; <0.0001 | 0.72; 1; 0.4 | 1.12; 1; 0.29 | - |
| Sperm viability (% live sperm) 1,2 | 73 | 3.97; 1; 0.046 | 0.08; 1; 0.77 | 3.53; 1; 0.06 | - |
| Survival post-spermatophore removal (% males) 2 | 109 | 3.99; 1; 0.046 | 3.87; 1; 0.049 | 0.0; 1; 1 | - |
| Longevity (N days alive) | 104 | 104.2; 1; <0.0001 | 0.0001; 1; 0.99 | 3.68; 1; 0.06 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McMahon, S.; Matzke, M.; Tuni, C. Food Limitation but Not Enhanced Rates of Ejaculate Production Imposes Reproductive and Survival Costs to Male Crickets. Cells 2021, 10, 1498. https://doi.org/10.3390/cells10061498
McMahon S, Matzke M, Tuni C. Food Limitation but Not Enhanced Rates of Ejaculate Production Imposes Reproductive and Survival Costs to Male Crickets. Cells. 2021; 10(6):1498. https://doi.org/10.3390/cells10061498
Chicago/Turabian StyleMcMahon, Saoirse, Magdalena Matzke, and Cristina Tuni. 2021. "Food Limitation but Not Enhanced Rates of Ejaculate Production Imposes Reproductive and Survival Costs to Male Crickets" Cells 10, no. 6: 1498. https://doi.org/10.3390/cells10061498
APA StyleMcMahon, S., Matzke, M., & Tuni, C. (2021). Food Limitation but Not Enhanced Rates of Ejaculate Production Imposes Reproductive and Survival Costs to Male Crickets. Cells, 10(6), 1498. https://doi.org/10.3390/cells10061498